
Nader A. Ashmawy
Department of Plant Pathology, Faculty of Agriculture, Alexandria University, El-Shatby, 21545, Alexandria, Egypt
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 4 | Page No.: 168-174







ABSTRACT
Brown rot disease of potato caused by the bacterium Ralstonia solanacearum (smith) is a destructive disease limiting potato production in several areas of the world. Nineteen isolates of the bacterium were recovered from infected potato tubers, showing typical symptoms of the disease that had been collected from Alexandria, Behera, Ismailia and Menofia Governorates. Isolates were identified as R. solanacearum biovar 2 (race 3), based on nutritional biochemical and enzymatic features as well as, standard biovar determination tests. All isolates were identified at the molecular level through Polymerase Chain Reaction (PCR) using a pair of specific primers (OLI-1 and Y2) that produced a 288 bp specific PCR product. Nested-PCR with the primer pair OLI-1/OLI-2 followed by JE2/Y2 proved to be highly specific and sensitive, producing the expected 220 bp band with all isolates tested beside a number of naturally infected potato tuber samples. The PCR- restriction fragment length polymorphism (PCR-RFLP) technique, based on the hypersensitive response and pathogenicity (hrp) gene region was used to determine race and biovar of nine R. solanacearum isolates. The RFLP banding patterns produced specific restriction profiles corresponded to biovar 2. The PCR-RFLP can, thus be used as a reliable sensitive technique to rapidly identify potential R. solanacearum isolates.
PDF Abstract XML References Citation
Received: July 03, 2015;
Accepted: August 11, 2015;
Published: September 29, 2015
How to cite this article
Nader A. Ashmawy, 2015. Detection and Molecular Characterization of some Egyptian Isolates of Ralstonia solanacearum by Nested-PCR and PCR-RFLP Analyses. Plant Pathology Journal, 14: 168-174.
DOI: 10.3923/ppj.2015.168.174
URL: https://scialert.net/abstract/?doi=ppj.2015.168.174
DOI: 10.3923/ppj.2015.168.174
URL: https://scialert.net/abstract/?doi=ppj.2015.168.174
INTRODUCTION
Bacterial wilt caused by Ralstonia solanacearum (Smith) (Yabuuchi et al., 1995) is one of the most serious diseases of crops in tropics, subtropics and warm temperate regions of the world. Ralstonia solanacearum causes brown rot and bacterial wilt on potato and consider one of the most important bacterial diseases in Egypt (Kabeil et al., 2008).
There has been significant progress in using PCR to detect and to identify of R. solanacearum from plants and the most useful primers (OLI1, OLI2, Y1 and Y2). These primers have been reported that permit amplification of a fragment specifically from R. solanacearum (Seal et al., 1992, 1993; Seal and Elphinstone, 1994; Opina et al., 1997; Hartung et al., 1998; Ito et al., 1998). The rapidity and sensitivity of detection of R. solanacearum are desirable characteristics that have been met by the use of a nested-PCR procedure (McManus and Jones, 1995; Niepold and Schober-Butin, 1997; Poussier and Luisetti, 2000 ; Khakvar et al., 2008).
Isolates of R. solanacearum are generally grouped, based on utilization of disaccharides and hexose alcohols into biovars and based on host range into races (Hayward, 1964; He et al., 1983; Kumar et al., 1993). A PCR-RFLP method has previously been used to assess the genetic diversity within a worldwide collection of R. solanacearum (Poussier et al., 1999). The whole hrp gene region, which is involved in the hypersensitive reaction and pathogenicity (Arlat et al., 1992; Boucher et al., 1992), was explored and this led to an accurate distinction of the major biovars of R. solanacearum.
With due attention to importance of R. solanacearum in Egypt and several irreparable losses that every year caused by this bacterium, this is prompted current study to use a PCR-based method using the previously reported primers (OLI-1, OLI-2, Y2 and JE2) for their sensitivity and specificity detection of the bacterium in potato tubers. Moreover, use a PCR-RFLP analysis to characterize and determine the biovar of R. solanacearum isolates.
MATERIALS AND METHODS
Bacterial isolation and culture conditions: Isolation trials were carried out from infected potato tubers with well developed symptoms of brown rot. Isolation source was from four locations in Egypt (Alexandria, Behera, Menofia and Ismailia governorates) during 2012-2013. Diseased potato tubers were surface sterilized using 1% sodium hypochlorite for 3 min, rinsed three times in sterile distilled water then, macerated in 3 mL sterile distilled water. The resulting suspension was streaked on 2,3,5-Triphenyl Tetrazolium Chloride (TTC) plates (Hugh and Leifson, 1953). Plates were incubated at 28°C and examined after 48 h. Presumptive colonies were purified by sub-culturing on Sucrose Peptone Agar (SPA) (Lelliott and Stead, 1987). Single cell colonies were transferred on to King’s B Medium (KBM) slants and stored at 4°C for further studies.
Nutritional and enzymatic tests: The following set of nutritional and enzymatic tests was conducted: Fluorescent pigment production, Gram reaction, metabolism of glucose, catalase activity, growth at 40°C, growth in 1 and 7% NaCl, arginine dihydrolase activity, levan formation, aesculin hydrolysis, starch hydrolysis and gelatin hydrolysis (Lelliott and Stead, 1987; Schaad et al., 2001).
Biovar determination: In order to determine the biovar of the isolated and confirmed R. solanacearum isolates, we tested their ability to utilize the following disaccharides and hexose alcohols: Maltose, lactose, D (+) cellobiose, mannitol, sorbitol and dulcitol (Hayward, 1964; Hayward et al., 1990).
DNA extraction protocol: Bacterial isolates were grown overnight in LB medium at 28°C with constant shaking at 200 rpm. Cells from 3 mL culture were pelleted by centrifugation at 6000 g for 5 min using a microcentrifuge. Cells of each culture were washed in TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0), then resuspended in a mixture of 567 μL Tris EDTA, 30 μL of 10% Sodium Dodecyl Sulphate (SDS) and 3 μL Proteinase K (20 mg mL–1). After incubation at 37°C for 1 h, 100 μL 5 M NaCl and 80 μL of CTAB/NaCl solution were added and the tubes were inverted well before incubation for 10 min in a water bath at 65°C. Phenol/chloroform/isoamyl alcohol (0.8 mL) was then added, mixed thoroughly and the tubes were centrifuged at 11000 g for 5 min. The aqueous supernatant was then taken and the phenol/chloroform step was repeated one more time. The DNA was precipitated by adding equal volume of isopropanol and washed with 70% ethanol. The DNA pellets were suspended in 100 μL sterilized distilled water (Ausubel et al., 1995).
Conventional PCR: A specific PCR was done using the primer pairs OLI-1 and Y2 (Table 1) as described by Seal et al. (1993). Total volume of 25 μL PCR mixture (3 μL genomic DNA, 12.5 μL Green Master Mix (Thermo Scientific™), 0.5 μL of each primer (10 μM) and 8.5 μL molecular grade water) was used. After initial heating to 94°C for 2 min, 35 amplification cycles were performed in a DNA thermal cycler (Techne, UK) through phases of denaturation (94°C for 20 sec), annealing (68°C for 20 sec) and extension (72°C for 20 sec), with a final period of 10 min at 72°C. The PCR product was run on 1.5% agarose gel containing ethidium bromide and visualized with a UV transilluminator.
Detection through nested-PCR: The reaction was preformed in final volume of 25 μL, reaction mixture contained 12.5 μL of 1ΧPCR Green Master Mix (Thermo Scientific™), 3 μL of genomic DNA or boiled extract of naturally infected potato tuber samples, 0.5 μL of each primer (10 μM) OLI-1 (Seal et al., 1993) and primer OLI-2 (Elphinstone et al., 1996) (Table 1) and 8.5 μL molecular grade water. After initial heating to 94°C for 2 min, 35 amplification cycles were performed in a DNA thermal cycler (Techne, UK) through phases of denaturation (94°C for 20 sec), annealing (68°C for 20 sec) and extension (72°C for 20 sec), with a final period of 10 min at 72°C. For the second PCR stage the same 25 μL reaction mixture was used, but with nested primers JE2 (Elphinstone et al., 1996) and Y2 (Seal et al., 1993) (Table 1), added 1 μL of the product from the first reaction. In the second stage, 30 cycles of denaturation (94°C for 20 sec), annealing (55°C for 20 sec) and extension (72°C for 20 sec), with a final period of 10 min at 72°C. PCR product was run on 1.5% agarose gel containing ethidium bromide and visualized.
PCR-restriction fragment length polymorphism (PCR-RFLP) analysis: The hrp genes of nine isolates of R. solanacearum were amplified by PCR using two different primer pair, RS600/RS61 and RS20/RS201 presented in Table 1 (Fouche-Weich et al., 2006).
| Table 1: | Nucleotide sequence of primers used in this study |
 | |
Amplified PCR products were analyzed by RFLP and digested with 3 units of Fast Digest restriction enzymes (Thermo Scientific™), HaeII for digestion of PCR product of primers RS600/RS61 and AvaI, PvuII for digestion of PCR product of primers Rs20/Ra201, in a total volume 30 μL containing 10 μL of PCR product and 2.5 μL enzyme buffer. Reaction mixtures were incubated at 37°C for 30 min (Poussier et al., 1999; Milijasevic-Marcic et al., 2013). The restriction fragments were separated by electrophoresis in 2.5% agarose gel containing ethidium bromide and visualized under UV-transilluminator.
RESULTS
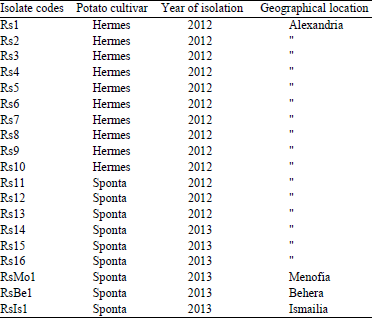
Isolation of the brown rot pathogen: Pure cultures of Ralstonia solanacearum were isolated from diseased potato tubers on TTC medium. A total of nineteen isolates were obtained from four governorates in Egypt and described in Table 2.
Nutritional and enzymatic identification tests: All bacterial isolates were arginine dihydrolase negative and oxidase, catalase positive. On the other hand, none of the isolates either hydrolyzed starch, aesculin or liquefied gelatin. All the isolates were highly sensitive to NaCl at 7% but not at 1%. Moreover, levan not formed from sucrose in all isolates. The results of these tests are shown in Table 3. According to these results, all bacterial isolates conformed closely to the set of nutritional and enzymatic features that are characteristic of R. solanacearum.
Biovar determination: All bacterial isolates utilized maltose, lactose and D (+) cellobiose, but not mannitol, sorbitol and dulcitol. According to these results, the investigated isolates were classified as biovar 2 of R. solanacearum (Table 4).
Conventional PCR: All of the nineteen R. solanacearum isolates showed one specific band with a molecular length of 288 bp when their DNA was used for amplification with specific primers OLI-1/Y2 (Fig. 1).
Detection of R. solanacearum though nested-PCR: Analysis of nested PCR products obtained from the nineteen R. solanacearum isolates with the primers (Y2 and JE2) was highly specific and sensitive; producing the expected band 220 bp with all tested isolates (Fig. 2), as well as from naturally infected potato tuber samples (Fig. 3).
PCR-RFLP analysis: PCR-RFLP used to more accurately identify the isolates as R. solanacearum. The hrp genes of R. solanacearum were amplified by PCR using two different primer pair, RS600/RS61 and RS20/RS201.
| Table 2: | Origin of the nineteen Ralstonia solanacearum isolates used in the present study |
 | |
| Table 3: | Nutritional and enzymatic tests |
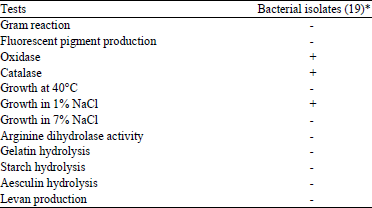 | |
| *Figures are No. of isolates, +: Positive result, -: Negative result | |
| Table 4: | Biovar determination tests |
 | |
| *Figures are No. of isolates, +: Positive reaction, -: Negative reaction | |
These primers enabled amplification of products of 905 bp (Fragment A) and 1452 bp (Fragment B) from nine isolates of R. solanacearum, respectively (Fig. 4). The restriction patterns of these PCR products were as follows, digestion of Fragment A with HaeII produced two main fragments, 410 and 540 bp (Fig. 5). Whereas digestion of Fragment B with AvaI produced three fragments, 610, 450 and 387 bp. However, when PvuII was used to digest the same material it produced three fragments, 595, 430 and 370 bp (Fig. 5). Results of RFLP banding patterns produced specific restriction profiles corresponded to biovar 2.
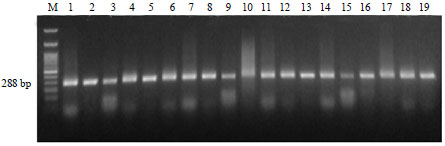 | |
| Fig. 1: | PCR amplification products resulting from the use of two specific primers OLY1/Y-2 in the bacterium Ralstonia solanacearum. Analysis included 19 isolates representing 4 different locations in Egypt. Products were observed upon separation by agarose gel electrophoresis followed by staining with ethidium bromide and visualization under UV-light lanes 1-16 are those of Alexandria isolates, Rs1, Rs2, Rs3, Rs4, Rs5 and Rs6, Rs7, Rs8, Rs9, Rs10, Rs11, Rs12, Rs13, Rs14, Rs15 and Rs16, respectively. Lanes 7, 8 and 9 correspond to the Menofia (RsMo1), the Behera (RsBe1) and the Ismailia (RsIs1) isolates, in the order given. M, a 100 bp DNA marker ladder. The molecular length of the DNA fragments constituting the amplification product band is indicated in base pairs (bp) |
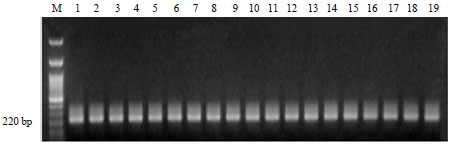 | |
| Fig. 2: | Agarose gel electrophoresis analysis of nested PCR products obtained from bacterial DNA of 19 isolates of Ralstonia solanacearum from 4 different locations in Egypt. The 220 bp PCR product amplified by second stage primer pairs JE2/Y2 is visible on the ethidium bromide stained gel. Lanes 1-16 are those of Alexandria isolates, Rs1, Rs2, Rs3, Rs4, Rs5 and Rs6, Rs7, Rs8, Rs9, Rs10, Rs11, Rs12, Rs13, Rs14, Rs15 and Rs16, respectively. Lanes 7, 8 and 9 correspond to the Menofia (RsMo1), the Behera (RsBe1) and the Ismailia (RsIs1) isolates, in the order given. M, a 100 bp DNA marker ladder. The molecular length of the DNA fragments constituting the amplification product band is indicated in base pairs (bp) |
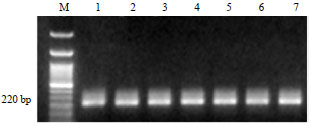 | |
| Fig. 3: | Detection of Ralstonia solanacearum by nested-PCR from potato tuber extracts. The 220 bp PCR product amplified by second stage primer pairs JE2/Y2 is visible on the ethidium bromide stained gel. Lanes 1-6 are naturally infected potato tuber samples. Lane 7 is a positive control prepared from R. solanacearum isolate RsMo1. M, a 100 bp DNA |
DISCUSSION
Potato brown rot, caused by Ralstonia solanacearum (Yabuuchi et al., 1995), has been reported in Egypt many years ago (Briton-Jones, 1925). The disease has created a lot of quarantine problems during the course of exportation of potatoes to Europe (Farag, 2000).
This study involved nineteen isolates of R. solanacearum isolated from four governorates in Egypt (Alexandria, Behera, Menofia and Ismailia). As expected, all isolates subjected to this study had the same cultural, morphological and physiological features as they all were highly fluidal white colored colonies with light pink center on TTC medium, short rods, Gram negative, motile and non-spore forming and gave the same biochemical reactions, when exposed to standard physiological testing, as those reported in the literature (Hayward, 1964; Lelliott and Stead, 1987; Hildebrand et al., 1988; Schaad et al., 2001).
Ralstonia solanacearum is a species complex that has been classified into five races based on host range (Buddenhagen et al., 1962; Pegg and Moffet, 1971; He et al., 1983) and into six biovars based on biochemical properties (Hayward, 1964, 1991). The biovar determination was made according to their ability to produce acids from hexose and alcohol sugars.
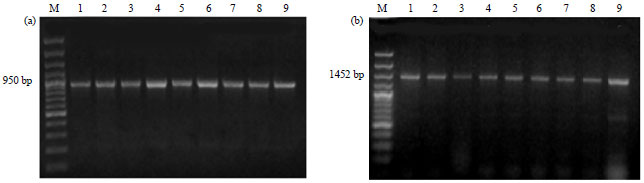 | |
| Fig. 4(a-b): | PCR amplification products resulting from the use of two primer pairs, (a) RS600-61 (Fragment A) and (b) RS20-201 (Fragment B) derived from (hrp) genes region in the bacterium Ralstonia solanacearum. Analysis included 9 isolates obtained from four different locations in Egypt. Lanes 1-6 are those of Alexandria isolates, Rs1, Rs4, Rs7, Rs10, Rs13 and Rs15, respectively. Lanes 7, 8 and 9 correspond to the Menofia (RsMo1), the Behera (RsBe1) and the Ismailia (RsIs1) isolates, respectively. M, a 100 bp DNA marker ladder. The molecular length of the DNA fragments constituting the amplification product band is indicated in base pairs (bp) |
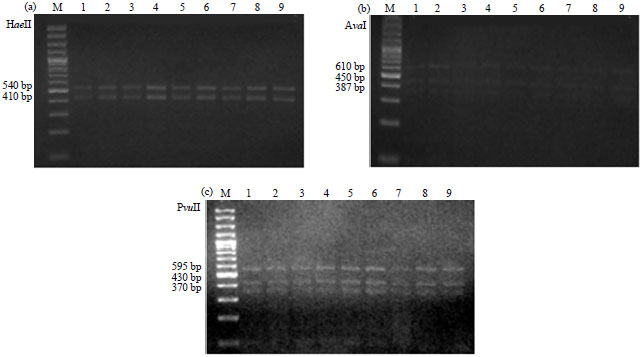 | |
| Fig. 5(a-c): | Restriction fragment patterns of hrp genes region for nine Ralstonia solanacearum isolates following digestion with the restriction enzymes, (a) HaeII for Fragment A, (b) AvaI and (c) PvuII for Fragment B. Lanes 1-6 are those of Alexandria isolates, Rs1, Rs4, Rs7, Rs10, Rs13 and Rs15, respectively. Lanes 7, 8 and 9 correspond to the Menofia (RsMo1), the Behera (RsBe1) and the Ismailia (RsIs1) isolates, respectively. M, a 100 bp DNA marker ladder. The molecular length of the DNA restriction fragments constituting each of the resulting bands is indicated in base pairs (bp) |
The nineteen isolates subject to identification were able to produce acids from maltose, lactose and cellobiose. All isolates, were unable to produce acids from mannitol, sorbitol and dulcitol denoting that the dominant race in Egypt is race 3, biovar II (Hayward, 1964; Hayward et al., 1990).
In recent years, studies on improved identification and detection of plant pathogens have mostly concentrated on molecular approaches because of their potential advantages of increased specificity and sensitivity. With the development of R. solanacearum specific PCR methods (Seal et al., 1993, 1999) were able to detect 16S rDNA sequences from a single cell grown in culture. In the present study, the 16S rRNA gene was amplified by PCR with primers OLI1 and Y2 (Seal et al., 1993, 1999). The PCR products with the expected band size were obtained from all the nineteen tested isolates of R. solanacearum confirming the isolated pathogen as R. solanacearum. Several variants have been developed to improve sensitivity of conventional PCR. Among the first described, nested PCR, with both internal and external primers to the target sequence, was reported to increase sensitivity and reduce the effect of inhibitors (Elphinstone et al., 1996, 1998; Poussier et al., 2000). In our study, Nested-PCR with the primer pair OLI-1/OLI-2 followed by Y2/JE2 proved to be highly specific and sensitive, producing the expected band with the naturally infected potato tuber samples.
Previous reports stated that DNA from closely related species, such as; Ralstonia eutropha, Ralstonia pickettii and Burkhoderia cepacia may be amplified with OLI1-Y2 primers (Seal et al., 1999; Poussier et al., 2000), resulting in false positive identifications of R. solanacearum. We, therefore, used PCR-RFLP based on (hrp) gene region to more accurately identify the isolates as R. solanacearum (race 3, biovar II) as described by Poussier et al. (1999, 2000), which described that the biovars could also be separated based on PCR-RFLP analysis.
Results of PCR-RFLP analysis were in agreement with those described for R. solanacearum confirming identification of the bacterium and allowing biovar determination (EU., 1998; Poussier et al., 1999, 2000).
The molecular methods presented here, Nested-PCR and PCR-RFLP are much less time consuming than conventional identification and detection methods. As a matter of fact, the early detection and identification of R. solanacearum is an integral part of successful disease management and this is specially important in relation to the importation of foreign plant material. The rapid detection of R. solanacearum like any other pathogen, allows for the appropriate control measures to be applied prior to the further spread of the disease or its introduction.
REFERENCES
- Arlat, M., C.L. Gough, C. Zischek, P.A. Barberis, A. Trigalet and C.A. Boucher, 1992. Transcriptional organization and expression of the large hrp gene cluster of Pseudomonas solanacearum. Mol. Plant-Microbe Interact., 5: 187-193.
PubMedDirect Link - Boucher, C.A., C.L. Gough and M. Arlat, 1992. Molecular genetics of pathogenicity determinants of Pseudomonas solanacearum with special emphasis on Hrp genes. Annu. Rev. Phytopathol., 30: 443-461.
CrossRefDirect Link - Elphinstone, J.G., J. Hennessy, J.K. Wilson and D.E. Stead, 1996. Sensitivity of different methods for the detection of Ralstonia solanacearum in potato tuber extracts. EPPO Bull., 26: 663-678.
CrossRefDirect Link - Elphinstone, J.G., H. Stanford and D.E. Stead, 1998. Survival and transmission of Ralstonia solanacearum in aquatic plants of Solatium dulcamara and associated surface water in England. EPPO Bull., 28: 93-94.
CrossRefDirect Link - Fouche-Weich, J., S. Poussier, D. Trigalet-Demery, D. Berger and T. Coutinho, 2006. Molecular identification of some African strains of Ralstonia solanacearum from eucalypt and potato. J. Gen. Plant Pathol., 72: 369-373.
CrossRefDirect Link - Hartung, F., R. Werner, H.P. Muhlbach and C. Buttner, 1998. Highly specific PCR-diagnosis to determine Pseudomonas solanacearum strains of different geographical origins. Theoret. Applied Genet., 96: 797-802.
CrossRefDirect Link - Hayward, A.C., 1964. Characteristics of Pseudomonas solanacearum. J. Applied Bacteriol., 27: 265-277.
CrossRefDirect Link - Hayward, A.C., H.M. El-Nashaar, U. Nvdegger and L. de Lindo, 1990. Variation in nitrate metabolism in biovars of Pseudomonas solanacearum. J. Applied Bacteriol., 69: 269-280.
CrossRefDirect Link - Hayward, A.C., 1991. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol., 29: 65-87.
CrossRefDirect Link - He, L.Y., L. Sequeira and A. Kelman, 1983. Characteristics of strains of Pseudomonas solanacearum from China. Plant Dis., 67: 265-277.
CrossRefDirect Link - Hugh, R. and E. Leifson, 1953. The taxonomic significance of fermentative versus oxidative metabolism of carbohydrates by various gram negative bacteria. J. Bacteriol., 66: 24-26.
CrossRefDirect Link - Ito, S., Y. Ushuima, T. Fujii, S. Tanaka, M. Kameya‐Iwaki, S. Yoshiwara and F. Kishi, 1998. Detection of viable cells of Ralstonia solanacearum in soil using a semiselective medium and a PCR technique. J. Phytopathol., 146: 379-384.
CrossRefDirect Link - Kabeil, S.S., S.M. Lashin, M.H. El-Masry, M.A. El-Saadani, M.M. Abd-Elgawad and A.M. Aboul-Einean, 2008. Potato brown rot disease in Egypt: Current status and prospects. Am. Eurasian J. Agric. Environ. Sci., 4: 44-54.
Direct Link - Khakvar, R., K. Sijam, W.M. Yun, S. Radu and T.K. Lin, 2008. Improving a PCR-based method for identification of Ralstonia solanacearum in natural sources of West Malaysia. Am. J. Agric. Biol. Sci., 3: 490-493.
CrossRefDirect Link - Milijasevic-Marcic, S., B. Todorovic, I. Potocnik, E. Rekanovic, M. Stepanovic, J. Mitrovic and B. Duduk, 2013. Ralstonia solanacearum-a new threat to potato production in Serbia. Pestic. Phytomed., 28: 229-237.
CrossRefDirect Link - McManus, P.S. and A.L. Jones, 1995. Detection of Erwinia amylovora by nested PCR and PCR-dot-blot and reverse-blot hybridizations. Phytopathology, 85: 618-623.
CrossRefDirect Link - Niepold, F. and B. Schober-Butin, 1997. Application of the one-tube PCR technique in combination with a fast DNA extraction procedure for detecting Phytophthora infestans in infected potato tubers. Microbiol. Res., 152: 345-351.
CrossRefDirect Link - Opina, N., F. Tavner, G. Hollway, J.F. Wang and T.H. Li et al., 1997. A novel method for development of species and strain-specific DNA probes and PCR primers for identifying Burkholderia solancearum (formerly Pseudomonas solancearum). Asia Pac. J. Mol. Biol. Biotechnol., 5: 19-30.
Direct Link - Pegg, K.G. and M.L. Moffett, 1971. Host range of the ginger strain of Pseudomonas solanacearum in Queensland. Aust. J. Exp. Agric. Anim. Husbandry, 11: 696-698.
CrossRefDirect Link - Poussier, S., P. Vandewalle and J. Luisetti, 1999. Genetic diversity of African and worldwide strains of Ralstonia solanacearum as determined by PCR-restriction fragment length polymorphism analysis of the hrp gene region. Applied Environ. Microbiol., 65: 2184-2194.
PubMedDirect Link - Poussier, S. and J. Luisetti, 2000. Specific detection of biovars of Ralstonia solanacearum in plant tissues by nested-PCR-RFLP. Eur. J. Plant Pathol., 106: 255-265.
CrossRefDirect Link - Poussier, S., D. Trigalet-Demery, P. Vandewalle, B. Goffinet, J. Luisetti and A. Trigalet, 2000. Genetic diversity of Ralstonia solanacearum as assessed by PCR-RFLP of the hrp gene region, AFLP and 16S rRNA sequence analysis and identification of an African subdivision. Microbiology, 146: 1679-1692.
Direct Link - Seal, S.E., L.A. Jackson and M.J. Daniels, 1992. Isolation of a Pseudomonas solanacearum-specific DNA probe by subtraction hybridization and construction of species-specific oligonucleotide primers for sensitive detection by the polymerase chain reaction. Applied Environ. Microbiol., 58: 3751-3758.
Direct Link - Seal, S.E., L.A. Jackson, J.P.W. Young and M.J. Daniels, 1993. Differentiation of Pseudomonas solanacearum, Pseudomonas syzygii, Pseudomonas pickettii and the blood disease bacterium by partial 16S rRNA sequencing: Construction of oligonucleotide primers for sensitive detection by polymerase chain reaction. J. Gen. Microbiol., 139: 1587-1594.
CrossRefPubMedDirect Link - Seal, S.E., M. Taghavi, N. Fegan, A.C. Hayward and M. Fegan, 1999. Determination of Ralstonia (Pseudomonas) solanacearum rDNA subgroups by PCR tests. Plant Pathol., 48: 115-120.
CrossRefDirect Link - Yabuuchi, E., Y. Kosako, I. Yano, H. Hotta and Y. Nishiuchi, 1995. Transfer of two Burkholderia and an Alcaligenes species to Ralstonia Gen. Nov.: Proposal of Ralstonia pickettii (Ralston, Palleroni and Doudoroff 1973) Comb. Nov., Ralstonia solanacearum (Smith 1896) Comb. Nov. and Ralstonia eutropha (Davis 1969) Comb. Nov. Microbiol. Immunol., 39: 897-904.
CrossRefDirect Link - EU., 1998. Council directive 98/57/EC of 20 July 1998 on the control of Ralstonia solanacearum (Smith) Yabuuchi et al. Official J. Eur. Communities, L235: 1-74.
Direct Link