
N. Taleb-Hossenkhan
Department of Biosciences, Faculty of Science, University of Mauritius, Reduit, 80837, Mauritius
A. Ibrahim
Department of Biosciences, Faculty of Science, University of Mauritius, Reduit, 80837, Mauritius
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 3 | Page No.: 113-122







ABSTRACT
Phytophthora infestans, the causal agent of potato and tomato late blight disease continues to be a serious threat in almost all potato and tomato growing areas around the world, with dramatic and disastrous economic consequences. In Mauritius, the disease is reported every year and many of the commercial potato varieties (Spunta, Delaware and Mondial) previously found to be moderately resistant or susceptible to the disease, have now become highly susceptible. This study reports the first phenotypic and genotypic characterization of local strains of Phytophthora infestans from the island of Mauritius. Seven isolates of Phytophthora infestans were collected from distinct geographical areas during the blight seasons of 2012 and 2013 and tested for aggressiveness on four potato and three tomato cultivars, metalaxyl sensitivity and subjected to isozyme analysis at the Gpi and Pep loci. Results show that all the isolates exhibited a clear host adaptation to potato, being only very weakly pathogenic on tomato. Most of the isolates (five out of seven) had high or intermediate metalaxyl resistance. Isozyme analysis revealed that at the Gpi locus, two isolates were 86/86, one isolate was 86/100, one isolate was 100/100 and one isolate was 122/122. At the Pep locus, two isolates were 92/92, two isolates were 92/100 and three isolates were 100/100. The combined results of the tests indicate that the seven isolates collected over the island are genetically distinct from each other and do not represent the same strain.
PDF Abstract XML References Citation
Received: April 03, 2015;
Accepted: June 02, 2015;
Published: June 27, 2015
How to cite this article
N. Taleb-Hossenkhan and A. Ibrahim, 2015. Pathogenic Fitness, Metalaxyl Sensitivity and Isozyme Analysis of Phytophthora infestans Isolates from Mauritius. Plant Pathology Journal, 14: 113-122.
DOI: 10.3923/ppj.2015.113.122
URL: https://scialert.net/abstract/?doi=ppj.2015.113.122
DOI: 10.3923/ppj.2015.113.122
URL: https://scialert.net/abstract/?doi=ppj.2015.113.122
INTRODUCTION
Among all the major food crops grown worldwide, potato (Solanum tuberosum L.), is known to suffer the greatest losses from disease attack (Olanya et al., 2001) and one of the major diseases affecting the crop is late blight disease, caused by the oomycete Phytophthora infestans (Mont.) de Bary. Historically, the oomycete is best known for having caused the Great Irish famine of 1845 and up till today, it still continues to be responsible for significant losses and increases production costs through the need for recurrent fungicide usage, annual reductions in marketable yields and postharvest losses. It is now generally accepted that the center of origin of P. infestans is in the central highlands of Mexico's Toluca Valley because of the high nuclear genetic diversity and the presence of sexual reproduction of the pathogen there (Gomez-Alpizar et al., 2007) and first migrated from there to the United States in infected wild potato tubers in the 19th century to cause famine-era epidemics. In the U.S., the pathogen infected potatoes and then spread to Europe and the rest of the world. The pathogen was thought to be asexual until the 1950s, when oospores were detected in blight lesions of potato in the central valley of Mexico (Niederhauser, 1991). Soon after, the A1 and A2 mating types of Phytophthora infestans were identified there but all the populations which were sampled outside Mexico were found to be of A1 mating type and thus asexual and belonged to a single clonal lineage, designated US-1 (Ib mtDNA haplotype). However, in the late 1970’s, different genotypes were identified in Europe and these were thought to result from a second migration event from Central Mexico (Goodwin et al., 1994). These migrant populations were characterized by the A2 mating type, which was shown to be widespread but present at highly variable frequencies and often quite rare. The working hypothesis was that the asexual global population had become established from a single A1 strain migrating from a center of diversity in Mexico in the mid 1800’s and that more recent migrations from Mexico of both A1 and A2 strains and subsequent mating among these had resulted in greater variation. Wherever, new genotypes have been detected, the old clonal lineage had rapidly died out and several studies have indicated that some new genotypes had traits (e.g., resistance to metalaxyl, rapid asexual reproduction) which gave them a selective advantage over the old clone (Kato et al., 1997).
Although, late blight has been widely reported in the potato growing countries of Sub-Saharan Africa (SSA) ranging from East and Central African countries, to Southern Africa and the West African country of Cameroon, the epidemiology and genetic identity of the prevailing P. infestans populations in these parts of the world and in Tropical Africa, is less known. The widespread transportation routes now characterizing global commerce means that the new diverse populations of the pathogen that have evolved in Europe, America and Asia, can more easily migrate to other places around the world. The introduction of the oomycete and its subsequent spread in Tropical Africa has most logically been accomplished primarily through the movement of potato seed, mainly from Europe but the primary sources of inoculum for P. infestans in most African countries remain unknown. In order to understand epidemiological patterns and be able to retrace primary sources of inoculum, it is essential to phenotypically and genotypically characterize local P. infestans isolates each year. Population studies of P. infestans in SSA have been conducted primarily on isolates from Uganda, Kenya and S. Africa and all isolates tested were shown to be of A1 mating type (US-1 clonal lineage) and exhibited variable response to metalaxyl (Vega Sanchez et al., 2000; Olanya et al., 2001). The isolates collected in South Africa had the characteristics typical of the pre-1980 population (A1 mating type, 86/100 Gpi genotype, US-1 fingerprint pattern and mtDNA haplotype Ib) previously found worldwide and 50% of the isolates were reported highly resistant to metalaxyl (McLeod et al., 1998). In Cameroon, 233 isolates of P. infestans collected from several hosts exhibited host preferences, variable aggressivity levels and variable levels of resistance to metalaxyl but no information is available on the genetic identity of the isolates (Fontem et al., 2005).
Mauritius is an island nation covering an area of 2,040 km2 in the Indian Ocean about 2,000 km (1,200 miles) off the Southeast coast of the African continent. Just like most SSA countries, the island is also not spared from late blight epidemics. Outbreaks date back to the 1970’s but the first reported devastating outbreak was recorded in 2004 that destroyed many potato fields, especially in the Centre of the island, where favorable conditions prevail i.e., low temperature and high humidity (MSIRI., 2005). Millions of rupees are spent on fungicides annually, although it is difficult to assess the economic impact of late blight uniquely, as potatoes are often infected with several pathogens at the same time, making it only possible to assess the combined effect of loss due to disease. An informal survey carried out in 2007, showed that the high cost of pest and disease control were among the major difficulties planters faced with their production, along with high cost of seed and shortage of labor, placing late blight as one of the most important limiting factors affecting potato production in the country, where the annual potato consumption is estimated to be around 18 kg/person. However, what also became apparent in the last ten years is that potato varieties previously known to be moderately resistant or susceptible to late blight, such as Spunta, Delaware and Mondial were now highly susceptible. The Mauritius Sugar Industry Research Institute therefore, tested several other varieties that were resistant to late blight and that would suit the local market. Variety Belle Isle was found to be very resistant to the disease, coupled with a higher rate of seed multiplication, was released in 2005 as a result of these efforts and strongly recommended to planters. However, it failed to be adopted by local planters as the market value of this variety was not as interesting as the traditionally cultivated ones. In 2011, the stakeholders targeted 24,000 tonnes of potato for that year in order to achieve self-sufficiency and the potato market was opened for importation of seed tubers by individuals, a prerogative that had been previously held by the Agricultural Marketing Board only. It is estimated that such a liberalization could lead to the introduction and spread of new seed-borne diseases such as late blight, especially in the absence of any controlled screening of seed tubers for potential pathogens as they are imported into the country.
The major factors favoring late blight epidemics such as use of susceptible varieties, diversity of pathogen virulence and races, lack of adequate disease management tactics and favorable environmental conditions have incidentally and consequently led to perpetuation and increase in late blight disease in the country. The paucity of information regarding the epidemiology and genetic characteristics of local late blight fungal isolates makes it very difficult to design and develop the most appropriate, site-specific, late blight management tactics on the field. The aim of this study was therefore, to start characterizing local P. infestans isolates, collected over a period of two years (2012 and 2013), in terms of aggressivity, metalaxyl sensitivity and isozyme analysis at the Gpi and Pep loci, in an attempt to build a larger database to report and monitor the exact identity of the various isolates that are present on the island.
MATERIALS AND METHODS
Sample collection, isolation and maintenance: Isolates were collected from infected potato fields over seven geographical locations during the blight seasons of 2012 and 2013. It is to be noted that these were the only blight outbreaks that growers officially reported to the authorities. These locations included La Marie, Bambous, Glen Park, St Pierre, Nouvelle France, Mare Longue and Plaine Sophie, situated in the Central and Eastern parts of the island mainly (Fig. 1). The isolates were henceforth, named LM, BB, GP, SP, NF, ML and PS. For each isolate, P. infestans-infected leaves from potato fields were collected and placed in a closed plastic bag with a moist piece of cotton wool which was then placed at 18°C overnight to favor sporulation.
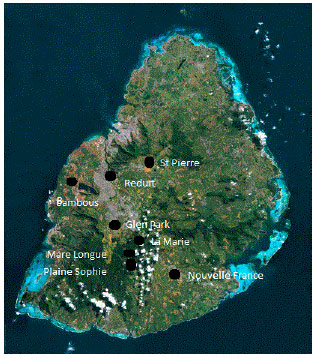 | |
| Fig. 1: | Map of Mauritius showing the locations where Phytophthora infestans isolates were collected during the blight seasons of 2012 and 2013 |
The next day, small pieces of infected leaf tissue from the sporulating border of lesions were cut and placed under potato slices of known susceptible potato cultivars in a petri dish. Dishes were incubated at 18°C for 1 week, until abundant sporulation could be observed on the surface of the potato slices. The mycelium was then transferred to pea agar (125 g of peas were heated in 50 mL of sterile distilled water for 20 min, allowed to cool then blended in a kitchen blender, sieved through sterile double layered muslin and the filtrate made up to 1000 mL with sterile distilled water), amended with antibiotics (50 mg L-1 Vancomycin, 50 mg L-1 Polymixin B, 200 mg mL-1, Ampicillin, 20 mg mL-1 Rifampicin, 50 mg L-1 PCNB and 50 mg L-1 benlate). Isolates were subsequently maintained by sub-culturing on pea agar with the same composition.
Aggressiveness testing: Aggressiveness tests were carried out using a detached-leaf assay as previously described (Spielman et al., 1992; Miller et al., 1998; Lebreton et al., 1999; Carlisle et al., 2002; Flier et al., 2003; Revelo et al., 2011; Fontem et al., 2005). Tests were carried out on four potato varieties, namely Belle Isle (highly resistant), Delaware (moderately resistant), Spunta (susceptible) and Pontiac (susceptible) and three tomato varieties, namely Tachian, Super Rose and NS501, whose resistance to late blight had not been tested. Three independent sets of experiments were carried out to ensure reproducibility of results. Measurements taken were: Incubation period, calculated as the number of days between inoculation and lesion development; Percentage infection frequency, calculated as the proportion of infected leaves (of the 10 replicates) for each isolate; Area Under the Lesion Expansion Curve (AULEC), the cumulative totals of the diseased lesion areas estimated daily (days 1-7) and sporulation intensity, estimated as the number of sporangia produced per centimeter square of lesion area seven days after inoculation.
Metalaxyl tests: Metalaxyl resistance was determined according to the method used by Shattock et al. (1990), Deahl et al. (1993), Hermansen et al. (2000) and Elansky et al. (2007) where, the mean percentage growth of each isolate on metalaxyl amended agar is compared with growth on non-amended agar and isolates are classified as being resistant, intermediate or sensitive to metalaxyl.
Isozyme analysis: Isozyme electrophoresis was conducted for the enzyme glucose-6-phosphate isomerase (Gpi) and peptidase (Pep) on cellulose acetate using a Helena Electrophoresis Unit (Helena Laboratories, Beaumont, TX), as previously described with the agar overlay method (Goodwin et al., 1995), with some slight modifications to the sample processing procedure. Samples were concentrated by collecting a small amount of mycelia from agar plates, grinding with 1 mL sterile distilled water, centrifuging at 13,000× g, removing excess supernatant, so that the final remaining volume was about 50 μL, grinding again with a micropipette tip, followed by centrifugation at 13,000× g for 1 min. Thirty microliter of the supernatant was then removed and mixed with bromophenol blue to be loaded onto the cellulose acetate sheets. Electrophoresis was carried out on cellulose acetate sheets at 200V for 15 min. Allozyme genotypes (inferred from banding pattern phenotypes on cellulose acetate) were scored as described previously and represent the mobilities of the enzyme alleles.
RESULTS
Aggressiveness testing: Figure 2 shows the aggressiveness (pathogenic fitness) of the seven collected P. infestans isolates on the four potato varieties (Belle Isle, Delaware, Pontiac and Spunta) and three tomato varieties (Super Rose, Tachian and NS501) tested and Fig. 3 shows the AULEC values for the seven isolates on the same potato and tomato varieties.
There are several important trends that can be inferred from Fig. 2. Firstly, all seven isolates exhibit host preference, with higher aggressiveness on potato varieties compared to the tomato varieties. Secondly, potato varieties Spunta, Pontiac and Delaware consistently prove to be more susceptible that potato variety Belle Isle. Thirdly, the aggressiveness characteristics of the seven isolates, although very similar, are not identical. For example, the aggressiveness order of isolate LM is Spunta>Pontiac>Delaware>Belle Isle, that of isolate BB is Delaware>Spunta>Pontiac>Belle Isle and that of isolate GP is Spunta>Delaware>Pontiac>Belle Isle. This indicates that the isolates have different pathogenic fitness properties and it is therefore, highly unlikely that these seven isolates represent the same strain of P. infestans. The AULEC values shown in Fig. 3 also indicate host specificity; values for the three tomato varieties Super Rose, Tachian and NS501 are hardly visible because they are comparatively so low.
 | |
| Fig. 2(a-g): | Aggressiveness of 7 Phytophthora infestans isolates tested on four potato and three tomato varieties, given as infection efficiency across 7 days post-inoculation, (a) LM, (b) BB, (c), GP, (d) SP, (e) NF, (f) ML and (g) PS |
Once again, a differential response is observed from the seven isolates on the four tested potato varieties, substantiating the claim that these seven isolates are most likely genetically different. Figure 3 also clearly indicates that the order of resistance to late blight in the four different potato varieties is Belle Isle>Pontiac> Delaware>Spunta.
Metalaxyl tests: For metalaxyl sensitivity, isolates were classified as resistant (relative growth of isolates on medium with fungicide concentration of 5 and 100 μg mL-1 is equal or greater than 40% compared to control (0 μg mL-1), Intermediate (relative growth of isolates on medium with fungicide concentration of 5 μg mL-1 is equal or greater than 40% compared to control, or sensitive (relative growth of isolates on medium with fungicide concentration of 5 and 100 μg mL-1 less than 40% compared to control).
 | |
| Fig. 3: | AULEC values for 7 P. infestans isolates tested on four potato and three tomato varieties, measured in mm2, 7 days post-inoculation. Each bar represents response from potato varieties Belle Isle, Spunta, Delaware and Pontiac and tomato varieties SuperRose, Tachian and NS501, respectively. (a) AULEC values for isolate LM, (b) AULEC values for isolate BB, (c) AULEC values for isolate GP, (d) AULEC values for isolate SP, (e) AULEC values for isolate NF, (f) AULEC values for isolate ML and (g) AULEC values for isolate PS |
 | |
| Fig. 4(a-b): | Graphical representations of the zymograms obtained for, (a) Gpi and (b) Pep analysis for all seven P. infestans isolates tested |
| Table 1: | Comparative growth of Phytophthora infestans isolates in metalaxyl-amended and non-amended agar |
 | |
Results show that isolates LM, GP, SP and ML were resistant; isolate BB was intermediate and isolates NF and PS were sensitive (Table 1).
Isozyme analysis: Isozyme analysis was conducted for the enzyme glucose-6-phosphate isomerase (Gpi) and peptidase (Pep) on cellulose acetate. At the Gpi locus, three different alleles are known to commonly occur: 86, 100 and 122 and at the Pep locus, two different alleles are known to commonly occur: 92 and 100. Figure 4 shows a graphical representation of the zymograms obtained after Gpi and Pep staining on the seven P. infestans isolates, carried out in three replicates each.
| Table 2: | Allozyme genotypes at the Gpi and Pep loci, metalaxyl resistance and pathogenic fitness of seven Phytophthora infestans isolates collected from the island of Mauritius from 2012-2013 |
 | |
Gpi staining was inconclusive for isolates NF and PS. Results show a wide diversity of allozyme genotypes among the seven P. infestans isolates at both the Gpi and Pep loci. Four out of five isolates tested proved to be homozygous at the Gpi locus, with either genotype 86/86, 100/100 or 122/122 and one showed to be heterozygous with genotype 86/100. At the Pep locus, five out of the seven tested isolates showed homozygosity, with genotypes either 92/92 or 100/100, while two showed to be heterozygous, with genotype 92/100.
Table 2 summarizes the main results of this study in terms of pathogenic fitness, metalaxyl sensitivity and allozyme genotypes of the seven P. infestans isolates collected in this study.
DISCUSSION
All of the isolates tested in this study were derived from infected potato fields and exhibited host specificity, being highly pathogenic on the four potato varieties tested and only very weakly pathogenic on the three tomato varieties tested. Host adaptation of Phytophthora infestans has been widely reported (Andrivon et al., 2004; Vega Sanchez et al., 2000), with isolates collected from potato plants being only weakly aggressive on tomato plants which is observed in this study and vice-versa. Several studies utilizing genetic markers have shown that distinct genotypes are generally associated with each host; in North-Western Mexico (Goodwin et al., 1992), the Philippines (Koh et al., 1994), the Netherlands (Fry et al., 1991), France (Lebreton and Andrivon, 1998), Ecuador (Oyarzun et al., 1998) and the USA (Legard et al., 1995). Host adaptation of Phytophthora infestans has important epidemiological consequences in areas where two or more potential hosts grow in close proximity which is often the case in Mauritius, where potato and tomato are often grown in the same field. If multiple hosts are cultivated and inoculum can pass readily among them, then disease management activities must take all hosts into consideration. All the isolates collected in this study, seem to be potato-adapted, thereby limiting the extent of cross-contamination between nearby potato and tomato infected fields. However, since P. infestans is known to evolve extremely rapidly, it is not unlikely that isolates that are specifically pathogenic on one host quickly adapt to other hosts, especially in areas where blight management strategies are inadequate. Although some exceptions to this phenomenon of host preference have been reported (Fraser et al., 1999; Goodwin et al., 1998), many authors have observed that isolates tend to be specifically adapted to either potato or tomato, both in "old" and in "new" populations (Legard et al., 1995; Lebreton et al., 1998; Oyarzun et al., 1998; Reis et al., 2003), so the cases where a single strain is equally pathogenic on multiple hosts is the exception rather than the rule. Host specificity is also of evolutionary significance, because the possibility for infecting more than one host determines to a large extent the availability of "green bridges" during the pathogen’s life cycle. These are critical in maximizing survival opportunities in species with very low saprophytic abilities, such as P. infestans and probably condition the extent of gene flow between isolates (Andrivon, 1995). The ability to survive and multiply on several hosts can thus be regarded as an evolutionary advantage and is an ability that will be evolutionary selected for to ensure the maximal survival chances of the pathogen in the long run.
Metalaxyl is a phenylamide systemic protectant fungicide which has been widely marketed for use in the control of late blight in several countries around the world. However, in almost all countries that have reported the occurrence of the "new" populations of P. infestans, these populations have been found to be mostly metalaxyl resistant. Metalaxyl resistance is now widespread in Europe (Carter et al., 1982; Davidse et al., 1981; Holmes and Channon, 1984; Dowley and O'Sullivan, 1985; Cooke, 1991; Daggett et al., 1993), the Middle East (Cohen and Reuveni, 1983), Asia (Kim et al., 1993; Therrien et al., 1993; Koh et al., 1994), Mexico (Matuszak et al., 1994), Canada and the United States (Deahl et al., 1993). In Europe, resistance to metalaxyl has been found to develop within 2-3 years of its use (Singh and Singh, 2006). It is not surprising therefore that, in this study, five out of the seven tested isolates were found to be metalaxyl resistant (one showing intermediate resistance), with only two being sensitive. Resistance arises when the frequency of resistant phenotypes of the pathogen increases as the result of a selection pressure imposed on the population by the use of the fungicide. Variation of phenotype in a population as well as existence of resistant phenotypes at low frequency, are necessary conditions for selection to occur. This selection pressure acts on phenotypes and hence, on the inheritable genes or quantitative traits which control the fungicide resistance phenotype. This means that the more widespread the use of the fungicide is, the greater the selection pressure imposed on the pathogen population which favors the occurrence of fungicide-resistant strains. This would be the case in areas where there is over-use of fungicides which has happened in many potato-growing countries before the phenomenon of fungicide resistance was properly understood and passed down to the grower community. Interestingly, metalaxyl is no longer an active ingredient commonly used in Mauritius. Fungicides applied include Infinito (fluopicolide+ propamocarb hydrochloride), Acrobat (dimethomorph), Electis (zoxamide+mancozeb) and Pennfluid (mancozeb). The metalaxyl-resistant P. infestans isolates found in this study could have developed their metalaxyl resistance as a result of the continuous use of this fungicide in the past as it has been widely used since the 1970’s until the early 2000’s. The rapidity with which P. infestans develop resistance to metalaxyl and the preponderance of metalaxyl-resistant P. infestans isolates in every part of the world today makes it clear that this phenylamide fungicide can no longer be considered as efficient to prevent blight outbreaks.
Interestingly, two of the metalaxyl-resistant isolates, GP and SP, also showed a phenomenon of growth stimulation by metalaxyl, i.e., the isolates grew at a quicker rate on metalaxyl-amended agar than on non-amended agar (data not shown). This has been previously reported (Zhang et al., 1997; Cohen and Reuveni, 1983), where metalaxyl was shown to promote sporulation in some isolates of P. infestans. While studying antibiotic resistance in P. infestans, Shattock and Shaw (1975) observed a similar stimulation of growth by an ordinarily inhibitory compound, where a streptomycin-resistant mutant of P. infestans only grew on solid medium containing streptomycin. Metalaxyl is known to inhibit RNA synthesis, specifically targeting RNA Polymerase I and/or III and resistance is due to alteration of the target site. Zhang et al. (1997) suggest that a possible explanation for the stimulation of growth by metalaxyl in some metalaxyl-resistant isolates is due to an altered RNA Polymerase I or RNA Polymerase III which, when bound to metalaxyl, exhibits enhanced enzyme activity. If this phenomenon occurs in vitro, it is also possible that metalaxyl might stimulate the same and closely related isolates in the field. Field studies analogous to such in vitro conducted tests are needed to assess the possible stimulatory role of metalaxyl in the field, as such data will have serious implications regarding the continued use of metalaxyl as a protectant fungicide in many countries.
Cellulose acetate electrophoresis provided a fairly good resolution of Gpi and Pep allozymes of P. infestans (Fig. 4), with the exception of Gpi assays for isolates NF and PS and scoring of genotypes was generally straightforward and easy. P. infestans is a diploid organism and Gpi and Pep are dimeric enzymes. Each individual will have two copies of each chromosome. Since, the enzymes being tested are dimeric, two subunits are required to form the active site. Homozygous individuals will produce single bands on gel and heterozygous will produce three bands, with the middle intermediate band being thicker. Homodimer bands are produced when subunits coded by same alleles come together to form the active enzyme and heterodimer bands arise when subunits coded by different alleles associate to form the active enzyme (Goodwin et al., 1995), with the heterodimer band usually migrating halfway between the two homodimer bands and being of twice the intensity of the two homodimer bands. For Gpi allozymes, the most commonly-occurring alleles are 86, 100 and 122 and for Pep, the most commonly-occurring alleles are 92 and 100. For Gpi allozymes, the band with the lowest electrophoretic mobility migrated a distance of 40 mm from the line of origin and this was assigned allelic identity 122. The band with the highest electrophoretic mobility migrated a distance of 65 mm from the line of origin and this was assigned allelic identity of 86. The band which migrated 45 mm from the line of origin was therefore, assigned allelic identity of 100. Therefore, for Gpi allozyme analysis, four different allozyme genotypes were found: 122/122 (isolate LM), 100/100 (isolate SP), 86/86 (isolates BB and ML) and 86/100 (isolate GP). For Pep allozymes, the band with the lowest electrophoretic mobility migrated a distance of 6 mm from the line of origin and this was assigned allelic identity 100. The band with the highest electrophoretic mobility migrated a distance of 21 mm from the line of origin and this was assigned allelic identity of 92. Therefore, for Pep allozyme analysis, three different allozyme genotypes were found: 100/100 (isolates GP, SP and NF), 92/92 (isolates LM and BB) and 92/100 (isolates ML and PS). Although the exact genotype of the seven tested P. infestans isolates may not be absolutely accurate (Forbes et al., 1998) list seven alleles for Gpi and five alleles for Pep), what this data indicates here is that those seven isolates are most likely to be genetically distinct because they possess unique combinations of alleles at the Gpi and Pep locus.
Taken together, the results of aggressiveness characteristics, metalaxyl resistance and allozyme analysis at the Gpi and Pep locus collectively indicate that the seven isolates of Phytophthora infestans collected over the island of Mauritius in 2012 and 2013 are genetically distinct from each other and do not represent the same isolate, even those collected in geographically close locations. This is quite a surprising observation because Mauritius is a small island and it has long been assumed, in both the scientific and grower community, that there was a single, or at most, two, isolate(s) of P. infestans that were causing all the recorded blight outbreaks over the island. This study demonstrates that this is not the case, therefore blight management tactics will need to be adapted to each situation, for instance, it would be useless to use metalaxyl as systemic protectant fungicide in those fields, where the isolate is metalaxyl-resistant but can be used in those fields where the isolates show sensitivity to metalaxyl. The liberalization of the potato import market, coupled with a lack of seed tuber testing for potential pathogens before cultivation, could possibly have contributed to the entry of a wide diversity of P. infestans spores through seed tubers onto the island. Now that this wide diversity in the local P. infestans population is known, it is necessary to re-enforce or ensure proper implementation of existing phytosanitary measures regarding potato seed certification.
Despite the large amount of work that has been and is currently being, done on understanding population characteristics and dynamics of Phytophthora infestans in many areas of the world, we are still a long way from a firm assessment of the exact factors that contribute to the evolution of populations of the fungus in the different geographical and ecological situations where the pathogen prospers. Several studies have shown that the characteristics of the groups of genotypes of the pathogen present in the various parts of the world, while different from those of the original set of clones (US-1 and related sub-clones), are also markedly distinct from one another; and this is also what is observed in this study. For instance, while populations present in North America tend to be quite simple, with one genotype dominating at each location in a given year (Goodwin et al., 1998), the setup of European populations is often much more complex, with a number of genotypes coexisting at any single time (Drenth et al., 1993; Lebreton and Andrivon, 1998; Lebreton et al., 1998; Cooke et al., 2003). This is also the case here on the island of Mauritius, where, within a very small area of 2,040 km2, we have at least seven distinct isolates of P. infestans that co-exist.
Forbes et al. (1998) compiled a global marker database for isolates of Phytophthora infestans originating from 41 locations which include 31 countries plus 10 regions within Mexico. The database contains information on 1,776 isolates for one or more of the following markers: Restriction Fragment Length Polymorphism (RFLP) "fingerprint" consisting of 23 bands, mating type, dilocus allozyme genotype, mitochondrial DNA haplotype, sensitivity to the fungicide metalaxyl and virulence. The authors performed a cluster analysis of genotypes from clonally propagated populations worldwide which generally confirmed a previously published classification of "old" and "new" genotypes. Genotypes from geographically distant countries were found to be frequently clustered and several old and new genotypes were found in two or more distant countries. Once additional genotypic characterization is performed on the seven isolates reported here, it would be interesting to find out where they cluster among existing populations registered in the global database. This would give us important clues as to the possible origin of the isolates found on the island and can help to reveal global migration patterns of the pathogen towards the region of Southern Africa and the Indian Ocean.
ACKNOWLEDGMENTS
The authors would like to acknowledge Dr. Salem Saumtally, Director of the Mauritius Sugarcane Industry Research Institute, for kindly reviewing this article. This work has been funded by the University of Mauritius.
REFERENCES
- Andrivon, D., R. Corbiere, L. Lebreton, F. Pilet, J. Montarry, R. Pelle and D. Ellisseche, 2004. Host adaptation in Phytophthora infestans: A review from a population biology perspective. Plant Breed. Seed Sci., 50: 15-27.
Direct Link - Andrivon, D., 1995. Biology, ecology and epidemiology of the potato late blight pathogen Phytophthora infestans in soil. Phytopathology, 85: 1053-1056.
Direct Link - Carter, G.A., R.M. Smith and K.J. Brent, 1982. Sensitivity to metalaxyl of Phytophthora infestans populations in potato crops in south-west England in 1980 and 1981. Ann. Applied Biol., 100: 433-441.
CrossRefDirect Link - Cohen, Y. and M. Reuveni, 1983. Occurrence of metalaxyl-resistant isolates of Phytophthora infestans in potato fields in Israel. Phytopathology, 73: 925-927.
Direct Link - Carlisle, D.J., L.R. Cooke, S. Watson and A.E. Brown, 2002. Foliar aggressiveness of Northern Ireland isolates of Phytophthora infestans on detached leaflets of three potato cultivars. Plant Pathol., 51: 424-434.
CrossRefDirect Link - Cooke, D.E.L., V. Young, P.R.J. Birch, R. Toth and F. Gourlay et al., 2003. Phenotypic and genotypic diversity of Phytophthora infestans populations in Scotland (1995-97). Plant Pathol., 52: 181-192.
CrossRefDirect Link - Daggett, S.S., E. Gotz and C.D. Therrien, 1993. Phenotypic changes in populations of Phytophthora infestans from eastern Germany. Phytopathology, 83: 319-323.
Direct Link - Davidse, L.C., D. Looijen, L.J. Turkensteen and D. van der Wal, 1981. Occurrence of metalaxyl-resistant ttrains of Phytophthora infestans in Dutch potato fields. Eur. J. Plant Pathol., 87: 65-68.
CrossRefDirect Link - Deahl, K.L., D.A. Inglis and S.P. DeMuth, 1993. Testing for resistance to metalaxyl in Phytophthora infestans isolates from northwestern Washington. Am. Potato J., 70: 779-795.
CrossRefDirect Link - Drenth, A., S.B. Goodwin, W.E. Fry and L.C. Davidse, 1993. Genotypic diversity of Phytophthora infestans in the Netherlands revealed by DNA polymorphisms. Phytopathology, 83: 1087-1092.
Direct Link - Elansky, S.N., V.P. Apryshko, D.I. Milyutina and B.E. Kozlovsky, 2007. Resistance of Russian strains of Phytophthora infestans to fungicides Metalaxyl and Dimethomorph. Moscow Univ. Biol. Sci. Bull., 62: 11-14.
CrossRefDirect Link - Flier, W.G., G.B.M. van den Bosch and L.J. Turkensteen, 2003. Stability of partial resistance in potato cultivars exposed to aggressive strains of Phytophthora infestans. Plant Pathol., 52: 326-337.
CrossRefDirect Link - Fontem, D.A., O.M. Olanya, G.R. Tsopmbeng and M.A.P. Owona, 2005. Pathogenicity and metalaxyl sensitivity of Phytophthora infestans isolates obtained from garden huckleberry, potato and tomato in Cameroon. Crop Protect., 24: 449-456.
CrossRefDirect Link - Forbes, G.A., S.B. Goodwin, A. Drenth, P. Oyarzun, M.E. Ordonez and W.E. Fry, 1998. A global marker database for Phytophthora infestans. Plant Dis., 82: 811-818.
CrossRefDirect Link - Fraser, D.E., P.B. Shoemaker and J.B. Ristaino, 1999. Characterization of isolates of Phytophthora infestans from tomato and potato in North Carolina from 1993 to 1995. Plant Dis., 83: 633-638.
CrossRefDirect Link - Fry, W.E., A. Drenth, L.J. Spielman, B.C. Mantel, L.C. Davidse and S.B. Goodwin, 1991. Population genetic structure of Phytophthora infestans in the Netherlands. Phytopathology, 81: 1330-1336.
Direct Link - Gomez-Alpizar, L., I. Carbone and J.B. Ristaino, 2007. An andean origin of Phytophthora infestans inferred from mitochondrial and nuclear gene genealogies. Proc. Nat. Acad. Sci. USA., 104: 3306-3311.
CrossRefDirect Link - Goodwin, S.B., B.A. Cohen and W.E. Fry, 1994. Panglobal distribution of a single clonal lineage of the Irish potato famine fungus. Proc. Nat. Acad. Sci. USA., 91: 11591-11595.
PubMedDirect Link - Goodwin, S.B., R.E. Schneider and W.E. Fry, 1995. Use of cellulose-acetate electrophoresis for rapid identification of allozyme genotypes of Phytophthora infestans. Plant Dis., 79: 1181-1185.
Direct Link - Goodwin, S.B., C.D. Smart, R.W. Sandrock, K.L. Deahl, Z.K. Punja and W.E. Fry, 1998. Genetic change within populations of Phytophthora infestans in the United States and Canada during 1994 to 1996: Role of migration and recombination. Phytopathology, 88: 939-949.
CrossRefDirect Link - Goodwin, S.B., L.J. Spielman, J.M. Matuszak, S.N. Bergeron and W.E. Fry, 1992. Clonal diversity and genetic differentiation of Phytophthora infestans populations in northern and central Mexico. Phytopathology, 82: 955-961.
Direct Link - Hermansen, A., A. Hannukkala, H. Naerstad and M.B. Brurberg, 2000. Variation in populations of Phytophthora infestans in Finland and Norway: Mating type, metalaxyl resistance and virulence phenotype. Plant Pathol., 49: 11-22.
CrossRefDirect Link - Holmes, S.J.I. and A.G. Channon, 1984. Studies on metalaxyl-resistant Phytophthora infestans in potato crops in south-west Scotland. Plant Pathol., 33: 347-354.
CrossRefDirect Link - Kato, M., E.S. Mizubuti, S.B. Goodwin and W.E. Fry, 1997. Sensitivity to protectant fungicides and pathogenic fitness of clonal lineages of Phytophthora infestans in the United States. Phytopathology, 87: 973-978.
CrossRefDirect Link - Koh, Y.J., S.B. Goodwin, A.T. Dyer, B.A. Cohen, A. Ogoshi, N. Sato and W.E. Fry, 1994. Migrations and displacements of Phytophthora infestans populations in East Asian countries. Phytopathology, 84: 922-927.
Direct Link - Lebreton, D. and D. Andrivon, 1998. French isolates of Phytophthora infestans from potato and tomato differ in phenotype and genotype. Eur. J. Plant Pathol., 104: 583-594.
CrossRefDirect Link - Lebreton, L., C. Laurent and D. Andrivon, 1998. Evolution of Phytophthora infestans populations in the two most important potato production areas of France during 1992-96. Plant Pathol., 47: 427-439.
CrossRefDirect Link - Lebreton, L., J.M. Lucas and D. Andrivon, 1999. Aggressiveness and competitive fitness of Phytophthora infestans isolates collected from potato and tomato in France. Phytopathology, 89: 679-686.
CrossRefDirect Link - Legard, D.E., T.Y. Lee and W.E. Fry, 1995. Pathogenic specialization in Phytophthora infestans: Aggressiveness on tomato. Phytopathology, 85: 1356-1361.
Direct Link - Miller, J.S., D.A. Johnson and P.B. Hamm, 1998. Aggressiveness of isolates of phytophthora infestans from the Columbia basin of Washington and Oregon. Phytopathology, 88: 190-197.
CrossRefPubMedDirect Link - Olanya, O.M., E. Adipala, J.J. Hakiza, P. Ojiambo, J.M. Mujalazi, G. Forbes and R. Nelson, 2001. Epidemiology and population dynamics of Phytophthora infestants in sub-Saharan Africa: Progress and constrains. Afr. Crop Sci. J., 9: 185-193.
Direct Link - Oyarzun, P.J., A. Pozo, M.E. Ordonez, K. Doucett and G.A. Forbes, 1998. Host specificity of Phytophthora infestans on tomato and potato in Ecuador. Phytopathology, 88: 265-271.
CrossRefDirect Link - Reis, A., C.D. Smart, W.E. Fry, L.A. Maffia and E.S. Mizubuti, 2003. Characterization of isolates of Phytophthora infestans from southern and southeastern Brazil from 1998 to 2000. Plant Dis., 87: 896-900.
CrossRefDirect Link - Revelo, E., G. Dorado, L.E. Lagos and O. Burbano-Figueroa, 2011. Foliar virulence of isolates of Phytophthora infestans sensu lato on detached leaves of two Solanum Betaceum cultivars. Trop. Plant Pathol., 36: 367-373.
CrossRefDirect Link - Shattock, R.C. and D.S. Shaw, 1975. Mutants of Phytophthora infestans resistant to and dependent upon, antibiotics. Trans. Br. Mycol. Soc., 64: 29-41.
CrossRefDirect Link - Shattock, R.C., D.S. Shaw, A.M. Fyfe, J.R. Dunn, K.H. Loney and J.A. Shattock, 1990. Phenotypes of Phytophthora infestans collected in England and Wales from 1985 to 1988: Mating type, response to metalaxyl and isoenzyme analysis. Plant Pathol., 39: 242-248.
CrossRefDirect Link - Singh, P.H. and B.P. Singh, 2006. Population dynamics of metalaxyl sensitive and resistance strains of Phytophthora infestans under controlled environment. Potato J., 33: 162-163.
Direct Link - Spielman, L.J., B.J. McMaster and W.E. Fry, 1992. Relationships among measurements of fitness and disease severity in Phytophthora infestans. Plant Pathol., 41: 317-324.
CrossRefDirect Link - Therrien, C. D., P.W. Tooley, L.J. Spielman, W.E. Fry, D.L. Ritch and S.E. Shelly, 1993. Nuclear DNA content, allozyme phenotypes and metalaxyl sensitivity of Phytophthora infestans from Japan. Mycol. Res., 97: 945-950.
CrossRefDirect Link - Vega‐Sanchez, M.E., L.J. Erselius, A.M. Rodriguez, O. Bastidas and H.R. Hohl et al., 2000. Host adaptation to potato and tomato within the US-1 clonal lineage of Phytophthora infestans in Uganda and Kenya. Plant Pathol., 49: 531-539.
CrossRefDirect Link - Zhang, S., D.G. Panaccione and M.E. Gallegly, 1997. Metalaxyl stimulation of growth of isolates of Phytophthora infestans. Mycologia, 89: 289-292.
Direct Link