
Khalid M. Ghoneem
Department of Seed Pathology Research, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
Said M. Ezzat
Department of Microbiology, Faculty of Science, Zagazig University, Zagazig, Egypt
Noha M. El-Dadamony
Department of Seed Pathology Research, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
Plant Pathology Journal
Year: 2014 | Volume: 13 | Issue: 4 | Page No.: 278-284







ABSTRACT
Mycobiota associated with seeds of twenty sunflower samples were studied in Egypt. A total of 20 genera and 31 species of fungi were recovered from the collected seed samples using Standard Blotter (SB) and Deep-Freezing Blotter (DFB) techniques. The two methods differed regarding to the frequency of recovered seed-borne fungi. Alternaria alternata, Aspergillus flavus, Rhizopus stolonifer, Penicillium spp. and Cladosporium spp. were the most abundant. Four fungi among those i.e., Macrophomina phaseolina, Fusarium oxysporum, F. solani and F. incarnatum, which are known as plant pathogens, were tested for their pathogenicity and transmission on sunflower seedlings. M. phaseolina caused the highest percentage (69.5%) of mortality of sunflower seedlings as compared with check treatment. The recovery rate of these pathogens gradually decreased from root up to the upper stem and did not reach to the stem apex.
PDF Abstract XML References Citation
Received: August 11, 2014;
Accepted: September 19, 2014;
Published: May 04, 2019
How to cite this article
Khalid M. Ghoneem, Said M. Ezzat and Noha M. El-Dadamony, 2014. Seed-Borne Fungi of Sunflower in Egypt with Reference to Pathogenic Effects and their Transmission. Plant Pathology Journal, 13: 278-284.
DOI: 10.3923/ppj.2014.278.284
URL: https://scialert.net/abstract/?doi=ppj.2014.278.284
DOI: 10.3923/ppj.2014.278.284
URL: https://scialert.net/abstract/?doi=ppj.2014.278.284
INTRODUCTION
Sunflower (Helianthus annuus L.), is an important member of the family Asteraceae and is one of the major oilseed crop grown for edible oil in the world. Sunflower seeds contain over more than 40% of good edible oil and 23% proteins. Beside this, it also constitute an excellent source of unsaturated fats, crude protein, fiber and important nutrients like vitamin E, selenium, copper, zinc and B-complex vitamin as well (Weiss, 2000; Gonzalez-Matute et al., 2002). In 2013, it was grown over an area of 9000 ha in Egypt with a total production of 22000 thousand tons while the annual Egyptian imports of sunflower oil was around 262764 thousand tons (FAO, 2014).
Sunflower is prone to be attacked by a variety of soil borne and seed borne fungal pathogens as well as by other phytopathogenic microorganisms and affected seeds may rot before emergence or emerging seedlings may be killed. Such seedlings exhibit damping off, root rot and downy mildew, leaf spot or stem rot symptoms (Mathur and Manandhar, 2003; Sangawan et al., 2005). Macrophomina phaseolina was reported as a most destructive soil and seed-borne fungal pathogens affecting sunflower in Egypt and worldwide. It cause charcoal-rot disease not only on sunflower but also on more than 500 plant species throughout the world (Purkayastha, 2006; Aboshosha et al., 2007). Of the fungal foliar diseases, leaf spot caused by Alternaria helianthi, Septoria helianth, Albugo tragopogonis and Plasmopara halstedii, inducing brown and grey spots, white rust and downy mildew, respectively, are relatively important (Masirevic and Jasnic, 2006; Van Wyk et al., 1999; Achbani et al., 2000). Fusarium wilt is caused by many species of Fusaria like Fusarium solani, F. oxysporum, F. helianthi, F. moniliforme, F. equestii and others (Masirevic and Jasnic, 2006). Sclerotina wilt and head rot of sunflower are caused by Sclerotinia sclerotiorum (El-Deeb et al., 2000). Several of these fungal species are reported to be seed borne. In addition, sunflower seeds are highly contaminated with fungi which attack the plants at different stages of development and subsequently during harvesting and storage (Afzal et al., 2010). In this respect, Rao (2006) surveyed the seed-borne fungi associated with sunflower by employing standard blotter method. The results of this study indicated the dominance of Alternaria helianthi (55.18%) followed by Rhizoctonia bataticola (14.71%) and A. alternata (8.79%). Other saprophytic fungi included Aspergillus flavus, A. niger and species of Penicillium, Rhizopus and Curvularia. Recently, Afzal et al. (2010) reported 13 phytopathogenic fungal species associated with seven cultivars of sunflower using agar and blotter paper methods including Alternaria alternata, Alternari helianthi, Aspergillus flavus, A. fumigatus, A. niger, Curvularia lunata, Drechslera tetramera, Fusarium solani, F. moniliforme, Macrophomina phaseolina, Mucor mucedo, Penicillium and Rhizopus spp. The isolated fungi were found to reduce seed germination by 10-20% and seedling mortality by 10-12%. Abdullah and Al-Mosawi (2010) reported 48 species of fungi belonging to 19 genera associated with sunflower seeds. A. niger, A. flavus, A. fumigatus, A. terreus, Chaetomium globosum, C. atrobrunneum, Alternaria alternata, Penicillium expansum, P. brevicompactum, Fusarium oxysporum, F. solani, Rhizopus stolonifer, Mucor hiemalis and A. ochraceus were the most frequent species.
In Egypt, Shahda et al. (1991), Khalil et al. (2014) and El-Wakil (2014) reported the association of large number of fungi with sunflower seeds and their list included; Alternaria alternata, Cladosporium spp., Fusarium oxysporum, F. moniliforme, F. proliferatum, F. semitectum, F. semitectum var majus, Macrophomina phaseolina, Rhizoctonia solani, Sclerotium rolfesii and Tricothecium rosum. In pathogenicity tests, Rhizoctonia solani and S. rolfesii were reported as the most destructive damping-off pathogens (97.5 and 92.5% seedling mortality) followed by M. phaseolina caused 48.5% seedlings mortality (Shahda et al., 1991).
The present study aimed to isolate and identify fungi from seeds of different cultivars of sunflower in Egypt, to determine the frequency of their occurrence and to study the pathogenicity and transmission of most prevalent seed-borne pathogenic fungi.
MATERIALS AND METHODS
Sample collection: Twenty seed samples of sunflower collected from growing fields in different regions of Dakahlia and Damiatta governorates during 2011-2012 were used in this study. The samples were collected in a 50x50 m area from each sampling site in a random zigzag pattern. Fully mature sunflower fruits were collected in cotton bags, labeled in the field, kept on ice until reached the lab and stored at 4°C until seed extraction. The extracted seeds were then spread out to dry on a porcelain plate at room temperature (25±2°C) for a few days. The seeds were then placed in a labeled envelope until testing.
Seed health testing: Detection of seed-borne fungi was done using recommended techniques by the International Seed Testing Association (ISTA, 1999) namely, Standard moist Blotter (SB) and Deep-Freezing Blotter (DFB) methods. A total number of 400 seeds from each sample were used. The percentage occurrence of each fungal species recovered by each method was calculated and compared.
Standard moist blotter method: Non-sterilized and surface-sterilized seeds (immersed into 1% Na(OCl)2 for 3 min) were plated in 9 cm diameter sterile petri dishes containing three layers of sterile blotter (filter paper) moistened with sterilized tap water. Ten seeds were placed in each petri dish and incubated at 20±2°C for 7 days under cool white fluorescent light with alternating cycles of 12 h light and 12 h darkness.
Deep freezing blotter method: The DFB method was used to detect a wide range of fungi which are able to arise easily from seeds in presence of humidity. After plating seeds as described in the SMB method, the dishes were incubated at 20±2°C for 24 h and transferred to a -20°C freezer for 24 h. This was followed by a 5 day incubation at 20±2°C under cool white fluorescent light with alternating cycles of 12 h light and 12 h darkness. Hyphal-tip and/or single-spore isolation techniques were followed to obtain pure cultures. Pure cultures of all fungi were maintained on potato carrot agar slants for further studies. Fungi were identified by recording their cultural properties, morphological and microscopic characteristics as described by Raper and Fennel (1965), Ellis (1971), Domsch et al. (1980), Booth (1977) and Burrges et al. (1988).
Pathogenicity test: Four fungal isolates (M. phseolina, F. oxysporum, F. solani and F. incarnatum) are most common in our surveys as well as worldwide known pathogenic fungi on sunflower were selected for pathogenicity test. Flasks containing 50 mL of potato dextrose broth were inoculated with disks (7 mm in diameter) taken from the growing edge of 5 day old colony of each fungus. The flasks were incubated in dark (without shaking) for 10 days at 25±2°C. Fifty grams of each mycelial mat were harvested and blended in 500 mL of sterile distilled water to produce fungal suspensions.
Healthy sunflower seeds (c.v. Giza 111) were surface sterilized in 1% sodium hypochlorite solution and soaked in fungal suspensions containing 2% Arabic gum for 15 min and left to dry at room temperature. Control treatment was carried out by soaking the disinfected seeds in tap water. Ten seeds per pot were planted in 20 cm diameter plastic pots containing sterile soil (2 kg soil/pot) and allowed to grow under greenhouse conditions. Ten replicates were used for each treatment. Data on pre-emergence damping off (% rotted seeds), post-emergence damping off (% infected seedlings) and plant survival were collected.
Transmission of seed borne fungi in sunflower plants: Sunflower plants surviving the challenge of the seed-borne fungi in the previous test were allowed to grow until maturity. Every two weeks, 10 plants were uprooted, washed, disinfected and dissected under sterile conditions. The various plant parts (roots, hypocotyls, basal stem, middle stem, upper stem, flowering branch top, inflorescence, flowers and seeds, if present) were plated on PDA and incubated at 24±2°C under cool white fluorescent light with alternating cycles of 12 h light and 12 h darkness for 4 days. Fungi recovered from each part were identified and the transmission rate and percentage were calculated.
Statistical analysis: Comparison of means was performed using LSD at p≤0.05 and the standard error was calculated using the statistical analysis software “CoStat 6.4” (CoStat, 2005).
RESULTS AND DISCUSSION
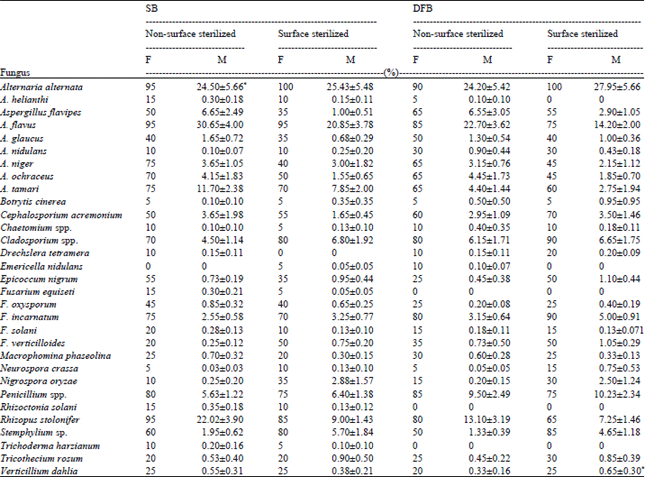
Occurrence of sunflower seed-borne mycoflora: Thirty-one fungal species belonging to 20 genera were isolated from the collected sunflower seed samples following Standard Blotter (SB) and Deep-Freezing Blotter (DFB) techniques. Considerable differences were observed between the SB and DFB techniques with regard to the frequency of the recovered seed-borne fungi (Table 1). Alternaria alternata, Aspergillus flavus, Rhizopus stolonifer, Penicillium spp. and Cladosporium spp. were most abundant. Alternaria alternata was the most frequent among those of the former and recovered from almost all samples while, A. helianthi was detected in lower percentages and have been documented causing characteristic leaf spots on sunflower in several parts of the world (Kumar and Sing, 1997; Bhutta et al., 1999). The results agree, at large, with many of the investigators (Shahda et al., 1991; Abdullah and Al-Mosawi, 2010; Afzal et al., 2010; Khalil et al., 2014) working on sunflower seeds. Large number of fungal species recovered from non-surface sterilized seeds were obtained by SB technique (19 genera and 30 species), as compared with DFB method (18 genera and 28 species) (Table 1). In addition, SB technique effectively detected the seed-borne saprophytes e.g, A. flavus (95%), A. niger (75%), A. ochraceus (70%), A. tamari (75%) and Rhizopus solonifer (95%). On the contrary, the DFB technique enhanced the recovery of A. flavipes (95%), A. glaucus (50%), Cephalosporium acremonium (70%), Cladosporium spp. (90%), F. incarnatum (90%), Macrophomina phaseolina (30%) and Tricothecium rosum (30%). It was observed that treated seeds yielded less population of seed-borne fungi in comparison to the untreated seeds indicating partial elimination of some contaminating fungi especially in DFB technique. These results are in close conformity with those of Afzal et al. (2010) who reported that chloral disinfection effectively reduced the microbial contamination. Similar findings were also reported from different seeds e.g., groundnut seeds by Rasheed et al. (2004) and legume seeds by Embaby and Abdel-Galil (2006).
Fusarium incarnatum was the most dominant species among all Fusarium species (75 and 80% in both SB and DFB techniques, respectively), followed by F. oxysporum and F. verticillioides (45, 20 and 25, 35%, in SB and DFB techniques, respectively) while, F. solani and F. equiseti were the least dominant among Fusarium species (20, 15 and 15, 0%, respectively). All of the isolated Fusarium species are known to be pathogenic to sunflower causing various symptoms in particular wilting and seedling rot (Qureshi et al., 2003; Sharfun-Nahar and Mushtaq, 2007).
Seven species of Aspergillus were isolated from seed samples are reported to produce different groups of aflatoxins which are natural toxins and hazardous to animals and human (Abdel-Mallek et al., 1994; Khalil et al., 2014). Among them, A. flavus, A. niger, A. ochraceus and A. tamari recorded highest occurrence, that may have affected seed germination and greatly lowered seed quality. Various threatening diseases including different types of carcinoma and immune system disorder, permanent damage of spleen, liver, kidney and brain functions in humans may develop, if such seeds are consumed (Sharfun-Nahar et al., 2005; Khalil et al., 2014).
In surface-sterilized seeds, high incidence of Cladosporium spp., Epicoccum nigrum, Fusarium, incarnatum, F. verticillioides, Nigrospora oryzae, Penicillium spp., Stemphylium sp. and Trichothecium rosum in both SB and DFB, Verticillium dahliae and Cephalosporium acremonium in DFB was observed while low incidence of Aspergillus species and Rhizopus stolonifer in both techniques was recorded. It was suggested that Cldosporium spp., N. oryzae, F. incarnatum and F. verticillioides typically internally seed-borne as compared with the other fungi were presumably externally seed-borne. On the other hand, seed surface sterilization led to complete absence of certain fungi (Drechslera tetramera in SB and Alternaria helianthi and Emericella nidulans in DFB method).
| Table 1: | Occurrence of sunflower seed-borne fungi using Standard Blotter (SB) and Deep-Freezing Blotter (DFB) methods |
 | |
| *Means±standard error | |
 |
This means that these fungi are externally seed-borne. Removal of externally seed-borne fungi by surface sterilization provided a chance for the internally seed-borne fungi to appear in greater numbers (Singh and Mathur, 2004).
Results of the present study revealed that sunflower seeds were infected with several pathogenic fungi such as Botrytis cinerea, F. incarnatum, F. oxysporum, F. solani, F. verticillioides, V. dahliae, M. phaseolina and R. solani. So, there is a serious need to increase public awareness on aspects related to seed health and to develop suitable management practices for improving the quality of the seeds. Seed health testing of major crops should be introduced in the national seed quality control system.
Pathogenicity tests: Four fungal species, i.e., M. phaseolina, F. oxysporum, F. solani and F. incarnatum were tested for their pathogenicity on sunflower seeds and seedlings. Pathogenicity tests were conducted in pots using surface-sterilized seeds of sunflower. Growing-on test showed that the disease symptoms were similar in all treatments of Fusarium species and were in form of rotted seeds and root, collar, stem and seedling rots, damping-off, stunting, wilting, tip burning and reduction in growth. Wilting and seedling rot were recorded as most prominent symptoms exhibited by all Fusarium spp. Infection with M. phaseolina produced disease symptoms of seedling blight damping-off, root rot and basal stem rot (Table 2). M. phaseolina caused the highest percentage of rotted seeds (46.7%), followed by F. solani (40%), F. oxysporum (37%) and F. incarnatum (31%) as compared with the check (2.5%). After 60 days, plants raised from the infected seed showed 23.97% seedling damping off due to infection of roots by M. phaseolina. Among Fusarium species tested, F. solani caused 19.7% wilting on seedlings, followed by F. oxysporum and F. incarnatum (15.2 and 12%, respectively). Two months after planting, results indicated that most tested fungi caused mild to severe infection on sunflower plants. M. phaseolina caused 69.5% mortality to sunflower seedlings while, F. solani exhibited seedlings mortality of 59.7%, followed by F. oxysporum (51.3%) and F. incarnatum (43%) as compared with check treatment. M. phaseolina was reported as a worldwide-distributed pathogen and is responsible for several diseases of sunflower, including seedling blight, damping-off, root rot, basal stem rot and charcoal rot (Khan, 2007). These results are in agreement with finding of Dawar (1994) and Sharfun-Nahar and Mushtaq (2007), who reported F. solani and F. oxysporum as major casual pathogens of collar and seedling rots, wilting and tip burning symptoms of sunflower seedlings. The results are also comparable to findings of Qureshi et al. (2003), where some strains of F. solani showed pathogenicity on their original as well as other hosts. It is interesting to note that F. incarnatum is not reported as pathogens of sunflower (as far as we know) caused significant diseases including wilting, root and seedling damping off, stunting and reduced the growth of plants.
| Table 2: | Pathogenicity of selected fungi isolated from sunflower seeds and type of symptoms they produced under greenhouse conditions1 |
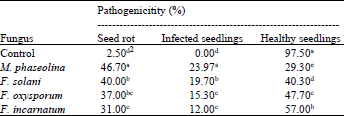 | |
| 1Affected plants with different fungi in the pathogenicity test were determined during seedling stage (1-6 weeks) as: Pre-emergence damping-off (rotted seeds) and Post-emergence damping-off (infected seedlings), 2Values are means of 15 replicates (pots), 10 seeds each. Values within a column followed by the same letters are not significantly different according to least significant difference test (p≤0.05) | |
Transmission of seed-borne fungi in sunflower plants: Sunflower plants surviving the challenge of the introduced seed-borne fungi (in the pathogenicity test) were left to grow until maturity. The rate of recovery of each fungus from various plant parts, including roots, crown, basal stem (from soil surface up to 10 cm height), middle stem (10-15 cm) and upper stem (15-20 cm) and stem apex at intervals of 60 days, was determined. Among the tested pathogens, M. phaseolina and F. solani showed the highest incidence on roots and crown parts of sunflower plants (95, 93.3 and 90, 80%, respectively), followed by F. oxysporum, F. incarnatum (90, 82.3 and 75, 74%, respectively). Isolation trials from basal, middle and upper stem parts showed that M. phaseolina, F. solani, F. oxysporum and F. incarnatum were restricted to basal stem part at incidence of 31.67, 10, 8.3 and 5.67%, respectively. Macrophomina phaseolina was the only fungus recovered from middle stem parts at infection percentage of 12.3%. However, the recovery percentages of the tested pathogens gradually decreased from root up to the middle stem and none of the pathogens except M. phaseolina has reached to the middle stem (Table 3). Pathogens are either extra-embryonal or embryonal, since infection was able to cause seed rot, seedling mortality and finally death of seedlings. In this case, the pathogen may spread from seeds (primary infection) to stems, petioles and leaves. The germ tube may penetrate the host and produce local infection (e.g., A. alternata) or live saprophytically for a period of time, persist in a resting stage in the soil or in plant residues and infect the host at a later time (e.g., Macrophomina phaseolina and Fusarium spp.) (Singh and Mathur, 2004). These results are in agreement with that of Khan (2007) who studied the location and transmission of M. phaseolina in naturally infected sunflower seeds. The results revealed that pathogen was located in seed coat, cotyledons and in embryonic axis of sunflower seedlings at various concentrations. These pathogens showed the disease cycle pattern of extra-embryal infection followed local infection.
| Table 3: | Incidence of pathogenic fungi in different parts of sunflower plants |
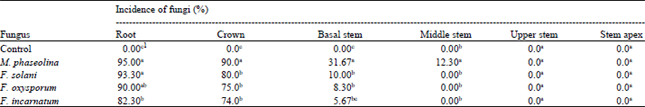 | |
| 1Each value represents the mean of 7 replicates, Values within a column followed by the same letters are not significantly different according to least significant difference test (p≤0.05) | |
The mechanism of seed germination may have a bearing upon the mode of transmission of inoculum from seed to seedling. In this respect, there are two types of host; epigeal in which the cotyledons are carried above ground and hypogeal in which the cotyledons still being covered by the seed coat. Epigeal cotyledons become green and may function like true leaves; hypogeal cotyledons remain pale and serve as storage and absorption organs. In hypogeal hosts, e.g., sunflower, the fleshy cotyledons act as a starting point, often as the food base, for invasion into roots and stem of the seedling. Often these exemplify intra-embryal infection followed by either local or systemic infection (Neergaard, 1979).
REFERENCES
- Abdullah, S.K. and K.A. Al-Mosawi, 2010. Fungi associated with seeds of sunflower (Helianthus annuus) cultivars grown in Iraq. Phytopathologia, 57: 11-20.
Direct Link - Aboshosha, S.S., S.I. Attaalla, A.E. El-Korany and E. El-Argawy, 2007. Characterization of Macrophomina phaseolina isolates affecting sunflower growth in El-Behera Governorate, Egypt. Int. J. Agric. Biol., 9: 807-815.
Direct Link - Achbani, E.H., A. Lamrhari, N. Laamaraf, M. Bahsine, M.N. Serrhini, A. Douira and D.T. de Labrouche, 2000. Downy mildew (Plasmopara halstedii): Importance and geographical distribution on sunflower in Morocco. Phytopathol. Mediterr., 39: 283-288.
Direct Link - Afzal, R., S.M. Mughal, M. Munir, K. Sultana, R. Qureshi, M. Arshad and M.K. Laghari, 2010. Mycoflora associated with seeds of different sunflower cultivars and its management. Pak. J. Bot., 42: 435-445.
Direct Link - Burrges, L.W., C.M. Liddell and B.A. Summerell, 1988. Laboratory Manual for Fusarium Research: Incorporating a Key and Descriptions of Common Species Found in Australia. 2nd Edn., Fusarium Research Laboratory, Department of Plant Pathology and Agricultural Entomology, University of Sydney, Sydney, Australia, ISBN-13: 9780949269560, Pages: 156.
- Domsch, K.W., W. Gams and T.H. Anderson, 1980. Compendium of Soil Fungi. Academic Press, Cambridge, Massachusetts, ISBN: 9780122204012, Pages: 1264.
Direct Link - El-Deeb, A.A., S.M. Abdallah, A.A. Mosa and M.M. Ibrahim, 2000. Sclerotinia diseases of sunflower in Egypt. Arab Univ. J. Agric. Sci., 8: 779-798.
Direct Link - El-Wakil, D.A., 2014. Seed-borne fungi of sunflower (Helianthus annuus L.) and their impact on oil quality. IOSR J. Agric. Vet. Sci., 6: 38-44.
Direct Link - Embaby, E.M. and M.M. Abdel-Galil, 2006. Seed borne fungi and mycotoxins associated with some legume seeds in Egypt. J. Applied Sci. Res., 2: 1064-1071.
Direct Link - Khalil, A.A., D.A. Elwakil and M.I. Ghonim, 2014. Mycoflora association and contamination with aflatoxins in sunflower (Helianthus annuus L.) seeds. Int. J. Plant Soil Sci., 3: 685-694.
CrossRef - Khan, S.N., 2007. Macrophomina phaseolina as causal agent for charcoal rot of sunflower. Mycopath, 5: 111-118.
Direct Link - Purkayastha, S., B. Kaur, N. Dilbaghi and A. Chaudhury, 2006. Characterization of Macrophomina phaseolina, the charcoal rot pathogen of cluster bean, using conventional techniques and PCR-based molecular markers. Plant Pathol., 55: 106-116.
CrossRefDirect Link - Rasheed, S., S. Dawar, A. Ghaffar and S.S. Shaukat, 2004. Seed borne mycoflora of groundnut. Pak. J. Bot., 36: 199-202.
Direct Link - Shahda, W.T., A.M. Tarabeih, S.H. Michail and A.H. Hemeda, 1991. Fungi associated with sunflower seeds in Egypt with reference to chemical control measures. J. King Saud Univ., 3: 287-293.
Direct Link - Qureshi, S.A., R. Riaz, V. Sultana, S. Ehteshamul-Haque and J. Ara, 2003. Pathogenicity and antimicrobial activity of seed-before Fusarium solani (Mart.) appel and wollenw. emend. snyd and hans strains. Pak. J. Biol. Sci., 6: 1183-1186.
CrossRefDirect Link - Sharfun-Nahar, M. Mushtaq and M.H. Hashmi, 2005. Seed-borne mycoflora of sunflower (Helianthus annuus L.). Pak. J. Bot., 37: 451-457.
Direct Link