
Swati Deep
Division of Plant Pathology, Indian Agricultural Research Institute, New Delhi, India
Pratibha Sharma
Division of Plant Pathology, Indian Agricultural Research Institute, New Delhi, India
Niranjan Behera
School of Life Sciences, Sambalpur University, Jyoti Vihar, Burla, Sambalpur, Odisha, India
Pallem Chowdappa
Division of Plant Pathology, Indian Institute of Horticultural Research, Bengaluru, India
Plant Pathology Journal
Year: 2014 | Volume: 13 | Issue: 4 | Page No.: 232-245







ABSTRACT
Morphological, cultural, pathogenic and genetic variability in thirty two Alternaria brassicicola (Schwein) Wiltshire isolates infecting cauliflower (Brassicae oleracea var botrytis) from different parts of India were studied here. Dark olivaceous black fungal colonies were observed with small, obpyrifom, septate, brown colored spores forming in chain having no beak. A significant (p<0.05) morphological variability was found within the isolates in respect to conidia length, width and number of septa whereas less cultural variability was seen with respect to colony colour and growth. High sporulation intensity was found in most of the isolates. RAPD (42) and ISSR (12) primers generated a total of 337 amplicons out of which 16 were monomorphic, indicating presence of high level of genetic diversity within the pathogenic A. brassicicola isolates. No major clusters were formed, rather isolates of mixed geographical regions of collection are found to be interlinked with each other. The internal transcribed spacer region of all the thirty two isolates showed 100% similarity with A. brassicicola accessions from NCBI.
PDF Abstract XML References Citation
Received: May 17, 2014;
Accepted: August 28, 2014;
Published: December 02, 2014
How to cite this article
Swati Deep, Pratibha Sharma, Niranjan Behera and Pallem Chowdappa, 2014. Diversity in Indian Isolates of Alternaria brassicicola (Schwein) Wiltshire Causing Black Leaf Spot Disease in Cauliflower. Plant Pathology Journal, 13: 232-245.
DOI: 10.3923/ppj.2014.232.245
URL: https://scialert.net/abstract/?doi=ppj.2014.232.245
DOI: 10.3923/ppj.2014.232.245
URL: https://scialert.net/abstract/?doi=ppj.2014.232.245
INTRODUCTION
As a second largest producer of vegetables next to China in the world, India accounts for about 15% of the world’s production of vegetables. Among vegetables, crucifers were reportedly affected with different fungal diseases specifically Alternaria blight which play a major role in production loss in many parts of the world (Hodgkin and MacDonald, 1986; Dillard et al., 1998; Pattanamahakul and Strange, 1999), including India (Meena et al., 2012; Sharma et al., 2013b). Severe damage of the foliage or seed germination in crucifers occurs due to Alternaria blight caused by Alternaria brassicae (Berk) (Kolte, 1985; Kolte et al., 1987; Tewari, 1991; Verma and Saharan, 1994; Sharma et al., 2013a, b) and A. brassicicola (Schw) (King, 1994; Jung et al., 2002; Sharma et al., 2013c). In vegetable brassica seeds, especially white cabbage and cauliflower, A. brassicicola is the dominant pathogen which causes more severe disease than A. brassicae (Maude and Humpherson-Jones, 1980; Humpherson-Jones, 1985; Maude et al., 1984; Kubota et al., 2006). Considerable damage occurs to cabbage and cauliflower in transit (Ellis, 2001). These species can survive in seeds for several months at different temperatures and relative humidity (Kumar and Gupta, 1994; Abul-Fazal et al., 1994) and the disease is spread during the growing season by wind-blown or rain-splashed spores (Rotem, 1994; MacKinnon et al., 1999; Oliver et al., 2001). The pathogen attacks most parts of the plant and it is thought to induce its chlorotic effect by release of phytotoxins (Jung et al., 2002).
Major taxonomic criteria for delimitation of fungal species including Alternaria are provided by morphological characteristics of conidia and conidiophores and sometimes host plant association (David, 1991; Pattanamahakul and Strange, 1999). All commercial cultivars of brassicas are susceptible to Alternaria brassicicola and no resistance reported till 1991 (Tewari, 1991). Many reports on the existence of morphological variability within the isolates of different Alternaria species have been reported by earlier workers (Verma and Saharan, 1994; Varma et al., 2006). Variability studies were done on the basis of morphology, sporulation, mycelial growth and other cultural characteristics (Ansari et al., 1989; Patni et al., 2005; Kaur et al., 2007). Morphological and cultural variability among the isolates of A. brassicae were reported from across the rapeseed-mustard and cauliflower growing region of India (Meena et al., 2005; Kaur et al., 2007; Singh et al., 2007; Goyal et al., 2011; Sharma et al., 2013a, b). However variability study in A. brassicicola infecting vegetable crop like cauliflower is still missing.
To understand host-pathogen co-evolution, epidemiology and resistance management, the study of genetic diversity of plant pathogen populations is also a basic need (Milgroom and Fry, 1997). The PCR based marker assisted genotypic identification of phytopathogens have been successfully going on at the species and subspecies level (Benali et al., 2011). Characterization of genetic variation in populations of Alternaria species pathogenic to crucifers by RAPD and nuclear ribosomal DNA sequences (Sharma and Tewari, 1995, 1998; Roberts et al., 2000; Tigano et al., 2003; Jasalavich et al., 1995) was reported with high level of similarity among the isolates of A. brassicae from different origin (Cooke et al., 1998; Sharma et al., 2013a, b). Occurrence of substantial genetic variability in A. solani and A. alternata infecting tomato and potato. Weir et al. (1998) and Morris et al. (2000) suggested diversity can be maintained by mutation, uniform host selection, extensive dispersal or the existence of a cryptic sexual stage. While resolving this, Bock et al. (2002, 2005) found moderate levels of genetic diversity in the form of substantial polymorphism between isolates from five populations of A. brassicicola attacking Cakile maritima along the New South Wales coast of Australia but the inter/intra species level variation among the Indian isolates of A. brassicicola (Schwein) Wiltshire infecting vegetable crucifers is still lacking. Therefore, the present study is aimed to study the morphological, cultural, pathogenic and genetic variability among the Alternaria brassicicola isolates collected from infected cauliflower (Brassicae oleracea L. var botrytis) from different part of India.
MATERIALS AND METHODS
Collection of isolates: Thirty two isolates (Rajasthan-6, Delhi-2, Tamil Nadu-3, Haryana-2, Jammu and Kashmir-2, Bihar-1, Uttar Pradesh-3, Himachal Pradesh-1, West Bengal-3, Uttarakhand-1, Maharashtra-1 and Assam-1, Orissa-2, Gujarat-1, Jharkhand-1, Punjab-1, Karnataka-1) of Alternaria spp. were obtained from infected leaf samples of cauliflower collected from seventeen different regions of India during 2009-2012. Blighted leaf pieces (2 mm) were surface sterilized with 0.1% Mercuric chloride (HgCl2) for 1 min, rinsed in sterile water 3x for 10 min and placed on Potato Dextrose Agar (PDA) plates. Fungal growth was observed after 5-7 days of incubation at 25°C and was confirmed to be A. brassicicola by microscopic (Carl Zeiss, Germany) observation (Ellis, 2001). Single spore cultures of these isolates were purified on Potato Dextrose Agar (PDA) plates at 25±1°C which were further maintained on PDA slants at 4°C.
For all isolates, morphological characteristics of the colony and sporulation apparatus were determined from single-spore colonies. The nature of mycelia growth (colony colour and radial growth), shape of conidia was noted. The size and shape of conidia (length and width) was determined using ocular and stage micrometer. Numbers of septa were also recorded.
Effect of different media on growth and sporulation: Seven types of media were tested to determine their effect on growth and sporulation of the A. brassicicola isolates. Seven different growth media were used namely Potato dextrose agar (PDA), Cauliflower Leaf Extract Agar (CLEA) made of Dextrose (20 g), agar (20 g) and Cauliflower leaf infusion from 200 g in 1000 mL water, Carrot Potato Agar (CPA), Oat Meal Agar (OMA), Czapex Dox Agar (CDA), V8 Juice Agar (VJA) and Corn Meal Agar (CMA) from Himedia, India. The experiment was arranged in a completely randomized design with 3 replicates. Measurements on radial colony diameter (in cm) were taken on 7th day after inoculation whereby six plates were sampled each time for each media treatment.
To determine conidial concentration of each isolates, cultures grown on seven different media plates were considered. The 10 mL of sterile distilled water was added to culture plate and using a sterile glass slide, the culture surface was gently scrapped to make a conidial suspension. Conidial concentration was determined using a haemocytometer.
Pathogencity: Thirty two A. brassicicola isolates from cauliflower were tested for symptom production on a susceptible cultivar of cauliflower viz., DC-23000. The seeds of cauliflower were sown in nursery beds during October, 2010-11 and 2011-12 seasons. Cauliflower seedlings were transplanted after two weeks in fields with a spacing of 30x40 cm. After 60 days leaves from both the cauliflower plant were taken for the detached leaf inoculation method (Sharma et al., 2004, 2013b). Test leaves were properly washed under running tap water and then surface wiped off with 70% alcohol and 2 μL of 4x104 spores mL-1 spores were inoculated with a fine needle (Dispovan, 2.5 mL) while sterile distilled water was applied on control. The leaves were placed inside moist chambers in greenhouse conditions and were observed for appearance of disease symptoms on 5 day after inoculation. Experiments were conducted in Completely Randomized Design (CRD). Each treatment consisted of three replicates. Symptoms observed were ranked as minus (-) for no symptom and plus (+) for black leaf spot with yellow halos on inoculated leaves. Appearance of symptom again divided into three groups viz., black spot with diameter 0.2-0.5 cm ranked as single plus (+), spot with diameter 0.6-1.0 cm ranked as double plus (++) and spot diameter more than 1cm were ranked as three plus sign (+++).
Genomic DNA isolation: The thirty two A. brassicicola isolates were grown on Potato Dextrose Broth (PDB) medium for 7 days at 28°C. Mycelial mat were harvested by Whatman No.1 filter papers and lyophilized. DNA was isolated according to a modified Cetyl Trimethyl Ammonium Bromide (CTAB) method (Doyle and Doyle, 1990; Connolly et al., 1994; Sharma et al., 2013b). Around 300 mg of lyophilized fungal mat was ground in liquid nitrogen, dispersed in 800 μL of 2% CTAB extraction buffer at 65°C in water bath for 30 min. An equal volume of Chloroform:Isoamyl alcohol (24:1, v/v) was added, mixed and centrifuged (4000 rpm, 10 min). The upper aqueous phase was transferred to a fresh tube. Nucleic acids were precipitated by adding 0.6 volume of ice cold isopropanol and collected by centrifugation (14000 rpm for 30 min). Pellet was washed twice with 70% ethanol and then air dried. Then pellet were solubilized in 100 μL TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH- 8.0) and treated with RNase A (20 mg mL-1) for 1 h at 37°C. Equal volume of Phenol:Chloroform:Iso-amyl alcohol (25:24:1) was added to the DNA solution and mixed by swirling for 5 min. After centrifuge at 10,000 rpm for 5 min, supernatant was collected in a fresh tube. This was followed by extractions with chloroform: iso-amyl alcohol (24:1). The purified DNA was precipitated by adding 1/10 volume of 3 M sodium acetate (pH 5.6) and 2.5 times (v/v) along chilled ethanol (95%). Extra salts were removed by further washing with 70% ethanol and DNA was pelleted and dried under vacuum. The quality and quantity of DNA was analyzed both spectrophotometrically using Nanodrop (Thermo-Scientific Ltd.) and in 1% agarose gel. The pellet was dissolved in a minimum volume of TE (10:1) buffer at room temperature and stored at -20°C.
RAPD analysis: Forty two different 10-mer oligonucleotide primers (Operon Technologies, USA) were tested on a set of 32 A. brassicicola isolates to identify polymorphic amplicons. PCR was performed in 15 μL volumes containing 1 μL DNA (50 ng), 1.5 μL primer (10 pmol), 1.5 μL of 10xPCR buffer, 1 μL of 25 mM MgCl2, 0.5 μL Taq DNA polymerase (0.5 U), 1.5 μL dNTP mix (2 mM), 8 μL double distilled water. Amplification were performed on a Bioneer-Mygenie 32 thermocycler programmed for 5 min at 95°C (denaturation), 40 cycles each of 1 min at 94°C, 1 min at 32°C (annealing), 1 min at 72°C and final extension of 2 min at 72°C (Sharma et al., 2013b). The amplified products were loaded onto 1.5% agarose gels (Agarose T2 Low EEO, Biomatrix) containing 0.2 mg L-1 Ethidium bromide (Sigma, USA) and electrophoresed in 1xTAE buffer. All the PCRs were repeated thrice to check the reproducibility of the amplicons.
ISSR analysis: Twelve different ISSR primers obtained from SBS Genetech Co., Ltd. were also tested on the above thirty two isolates of A. brassicicola to identify polymorphism. PCR was performed in 15 μL volume reaction mixture same as for RAPD analysis but with different ISSR primers. The PCR amplifications were performed in the same thermocycler for 5 min at 95°C (denaturation), 40 cycles of 30 sec at 94°C, 30 sec at 51°C (annealing), 45 sec at 72°C and final extension of 7 min at 72°C. Again the amplified products were electrophoresed in 1.5% agarose in 1xTAE buffer after adding Ethidium bromide.
Internal Transcribed Spacer (ITS) analysis: Thirty two A. brassicicola isolates were analyzed by amplifying the regions of the rDNA repeat from the 3’end of the 18 s and the 5’ end of the 28 s gene using PCR conditions with the two universal primers, ITS1-3’TCCGTA GGTGAACCTGCGG5’ and ITS4-3’TCCTCC GCT TA TT GATATGC5’ which were synthesized on the basis of conserved regions of the eukaryotic rRNA gene (White et al., 1990; Jasalavich et al., 1995). The PCR- amplification reactions were performed in a 25 μL mixture containing 50 mM KCl, 20 mM Tris HCl (pH 8.0), 2.0 mM MgCl2, 2 mM of dNTP mix, 20 mM of each primer with 50 ng μL-1 of template and 2.5 U of Taq polymerase. These reactions were subjected to an initial denaturation at 94°C for 4 min, followed by 40 cycles of denaturation at 94°C for 35 sec, primer annealing at 60°C for 1 min 30 sec and primer extension at 72°C for 3 min and a final extension for 10 min at 72°C in the thermal cycler. Aliquots (10 μL) of the amplified products were analyzed by electrophoresis in 1.5% (w/v) agarose gel in 1xTAE buffer (40 mM Tris, 20 mM acetic acid, 1 mM EDTA [pH 8]), stained with ethidium bromide (1 μg mL-1) and electrophoresis was carried out at 70 V for 2 h in TAE buffer. The molecular marker was 100 bp ladder (Biomatrix Co. Ltd.). The desired bands were cut from the gel with minimum quantity of gel portion and the amplified PCR product was eluted using QIAGEN gel extraction kit as per instruction manual.
Nucleotide sequencing and in silico analysis: The sequencing of the PCR product was carried out in automated Sequencer at Xcelris Lab., Ahmadabad, India. Related sequences were searched for homology using BLAST bioinformatic search tool available at the Gen-Bank database (http://www.ncbi.nlm.nih.gov/blast/) (Altschul et al., 1997). The multiple sequence alignment and pairwise alignment were performed using the ClustralW algorithm in Bioedit (Hall, 1999) and Phylogenetic analysis was done using MEGA 5.0 (Tamura et al., 2011) software. To assess the possible phylogenetic relationship neighbour-joining tree (Saitou and Nei, 1987) was created using maximum composite likelihood method (Tamura et al., 2004) and Parsimony tree was created using Close-Neighbor-Interchange algorithm (Nei and Kumar, 2000) with search level 0 in which the initial trees were obtained with the random addition of sequences (10 replicates). The sequences were aligned among themselves as well as with other published sequences available in GenBank using BlastN.
Statistical analysis: Averages of colony diameter on each media plates and number of conidia per milliliter for each isolates were taken for subsequent data analysis. Analysis of variance and separation of means to determine differences in growth rates on media types was analyzed by performing two-way ANOVA using statistical software PRISM version 3.0 at p<0.0001. Similarly the conidial characters were also statistically analyzed at p<0.05 by performing one way ANOVA and least significant difference was calculated by student’s t-test.
Dendrogram were constructed employing nested un-weighted pair group method using the arithmetic averages (UPGMA) and the sequential, agglomerative, hierarchical, non-overlapping (SAHN) clustering was obtained by NTSYS-PC Version 2.02 h (Rohlf, 1997). Resolving Power (RP) of the RAPD and ISSR primer was calculated as per (Prevost and Wilkinson, 1999). RP = ΣIB, where IB (Band informativeness) = 1-{2x (0.5-P)}; P is the proportion of the 32 isolates of A. brassicicola analyzed that contains the band.
RESULTS AND DISCUSSION
Single spore culture of thirty two isolates of Alternaria brassicicola (Schweinitz, Wiltshire) were isolated from seventeen different cauliflower growing states of India (Table 1). These isolates were further maintained on PDA slants at 4°C for further study.
Colony colour and radial growth were observed among thirty two isolates of Alternaria brassicicola infecting cauliflower in India. The colour of A. brassicicola colonies on PDA plates varies between olive gray to dark olivaceous black. Mycelia colour of all tested isolates was found to be brown. Conidial characteristics of each isolate were found similar to each other viz., obpyriform conidia having brown colour with smooth surface and short beak (Table 1). A Significant (p<0.05) variation was observed in conidia length and breadth (Table 2) of the thirty two single spore cultures of A. brassicicola. Average conidia length varied between 32.57 μm in CaAbcP1 from Punjab to 40.08 μm in CaAbcUP2 from Uttar Pradesh. Similarly average conidial breadth varied from 6.23 μm in CaAbcJK1 from Jammu and Kashmir to 9.40 μm in CaAbcWB3 from West Bengal. Average number of horizontal septa varied between 1.5-3. No vertical septum was found in any of the conidia. From the conidia size and shape it was found that all the isolates of A. brassicicola are almost similar to each other. Similar results were also showed by Ellis (1971), Pattanamahakul and Strange (1999) and Jung et al. (2002). The fungus differs from A. brassicae which is also majorly responsible for the black leaf spot in crucifers in India (Sharma et al., 2013a; Meena et al., 2012; Singh et al., 2012) in its beakless and smaller conidia (Cho et al., 2001). All the thirty two isolates of A. brassicicola showed significant (p<0.05) morphological variability in respect of conidia length, conidia width and number of septa.
Seven different nutrient media brought slight variation in the mycelial colony growth of thirty two isolates of A. brassicicola. According to average radial growth of different isolates on different media, Cauliflower Leaf Extract Agar (CLEA) and Potato Dextrose Agar (PDA) was found optimum for all isolates. On CLEA most of the isolates attended maximum radial growth (>6 cm) with CaAbcO1 from Orissa having maximum 7.5 cm except nine isolates viz., CaAbcR1, CaAbcR3 (Rajasthan), CaAbcH1 (Haryana), CaAbcJK1 (Jammu and Kashmir), CaAbcWB2 (West Bengal), CaAbcMH1 (Maharashtra), CaAbcO2 (Orissa), CaAbcD1 (Delhi) and CaAbcK1 (Karnataka) with the least in CaAbcWB2 (4.83 cm). On PDA, highest radial growth was obtained in CaAbcUP3 from Uttar Pradesh (6 cm) and least in CaAbcR6 from Rajasthan (4.40 cm). On CDA, PCA, CMA and VJA highest growth were obtained in CaAbcR4 (5.63 cm), CaAbcR5 (5 cm), CaAbcWB2 (5.03 cm) and CaAbcP1 (5.97 cm), respectively whereas least growth was found in CaAbcH1 (2.17 cm), CaAbcUP2 (3.23), CaAbcR1 (3.53 cm) and CaAbcJK1 and CaAbcUP1 (2.97 cm each), respectively.
| Table 1: | Morphological and cultural characteristics of thirty two isolates of Alternaria brassicicola collected from Cauliflower growing states of India with their accession numbers |
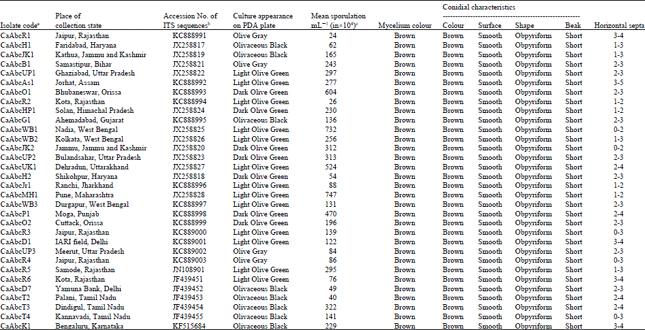 | |
| aIsolate coding was done as Ca: Cauliflower, Abc: Alternaria brassicicola, Alphabets and numbers in the Isolate code: State of collection and number of sample isolated. bAccession numbers of ITS sequences of the A. brassicicola isolates deposited to NCBI database. cMean Sporulation mL¯1 (in×104) of A. brassicicola isolates on different nutrient media | |
| Table 2: | Variation in A. brassicicola isolates based on colony growth on different nutrient media, conidial size and pathogenicity response on susceptible cauliflower cultivar (DC-23000) |
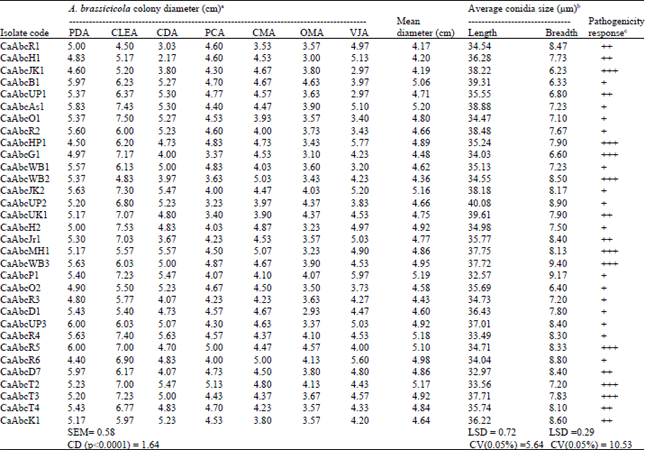 | |
| aMean radial growth (cm) of A. brassicicola isolates measured on different nutrient media in six replications. bThe size of conidia (length and width in μm) was determined using ocular and stage micrometer. cPathogenicity response of A. brassicicola isolates on susceptible cauliflower cultivar (DC-23000) where symptoms observed were ranked as minus (-) for no symptom and plus (+) for black leaf spot with yellow halos on inoculated leaves. Appearance of symptom again divided into three groups viz., black spot with diameter 0.2-0.5 cm ranked as single plus (+), spot with diameter 0.6-1.0 cm ranked as double plus (++) and spot diameter more than 1cm were ranked as three plus sign (+++) | |
The OMA showed the lowest radial growth ranging from 2.93 cm (in CaAbcD1) to 4.63 cm (in CaAbcB1) (Table 2).
Sporulation pattern of each isolates of A. brassicicola on 7th day on seven different nutrient media was similar to each other. The lowest sporulation (20-50x104 mL-1) was observed for the isolates from Rajasthan (CaAbcR1, CaAbcR2), Delhi (CaAbcD7), and Tamil Nadu (CaAbcT2). Moderately sporulating (50-100x104 mL-1) were found for the isolates from Haryana (CaAbcH2), Jharkhand (CaAbcJR1), Uttar Pradesh (CaAbcUP3) and Rajasthan (CaAbcR4 and CaAbcR6). Rest of the isolates were found highly sporulating (>100x104 mL-1), among which the Orissa isolate CaAbcO1 was found to be the highly sporulating with 604x104 mL-1 followed by CaAbcUK1 (524x104 mL-1) from Uttarakhand, CaAbcP1 (470x104 mL-1) from Punjab and CaAbcT3 (322x104 mL-1) from Tamil Nadu (Table 1).
All the A. brassicicola isolates from cauliflower were found pathogenic in nature. Among these nine isolates viz., CaAbcJK1, CaAbcHP1, CaAbcG1, CaAbcWB2, CaAbcWB3, CaAbcMH1, CaAbcR5, CaAbcT2 and CaAbcT3 were highly pathogenic as the spot size produced by them are >1cm in diameter. Eight isolates namely CaAbcR1, CaAbcH1, CaAbcUP1, CaAbcUK1, CaAbcJr1, CaAbcD7, CaAbcT4 and CaAbcK1 were found moderately pathogenic as they producing spot size of 0.6-1 cm and rest fifteen isolates were found lowest pathogenic as the spot size were 0.2-0.5 cm in diameter (Table 2). These results are in agreement with earlier workers on different Alternaria species (Awasthi and Kolte, 1989; Varma et al., 2006; Meena et al., 2005, 2012; Kaur et al., 2007; Singh et al., 2007, 2012; Reis and Boiteux, 2010; Sofi et al., 2013; Sharma et al., 2013b; Sun and Zhang, 2008), who observed morphological, cultural, pathogenic variability in different geographical isolates.
Forty two random decamer oligonucleotide primers were utilized for RAPD analysis but two primers did not give satisfactory amplification, so were not considered further. Thirty nine primers resulted in the amplification of distinct and reproducible bands in the present investigation. All the primers produced wide range of fragments/amplicons ranging from 100-1500 bp. The highest number of fragments (199) was amplified by the primer OPA8 and the lowest by the primer OPX13 (7). The number of polymorphic bands produced were maximum with the primers OPA3 and OPA17 (9 each), while minimum (3) with the primers OPA9 and OPC5. In case of monomorphic bands the maximum number of bands (3) was produced by the primer OPA9 followed by OPA6 and OPA13 with two bands each. Among the rest, seven primers viz., OPA4, OPA5, OPA8, OPC7, OPE3, OPE4 and OPE6 yielded each single monomorphic band. The Resolving Power (RP) was highest in case of OPA8 (12.4375), whereas the lowest RP was observed in OPX13 (0.4375). Total 14 monomorphic bands were amplified with all the RAPD primers used in the present study. The details of RAPD banding pattern was presented in Table 3 and Fig. 1a. Jaccard’s similarity coefficient was calculated from the RAPD data showed that all the isolates were correlated with each other with an average similarity of 0.56.
| Table 3: | Details of RAPD and ISSR banding pattern in different isolates of Alternaria brassicicola of Indian origin |
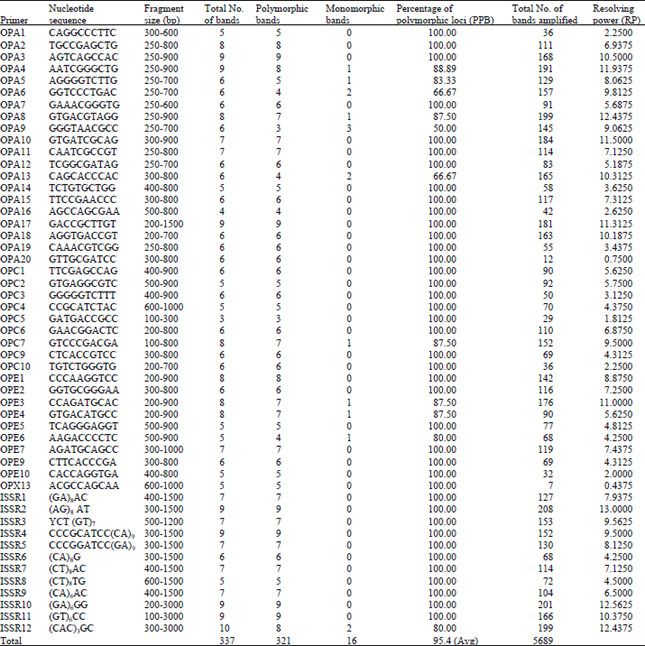 | |
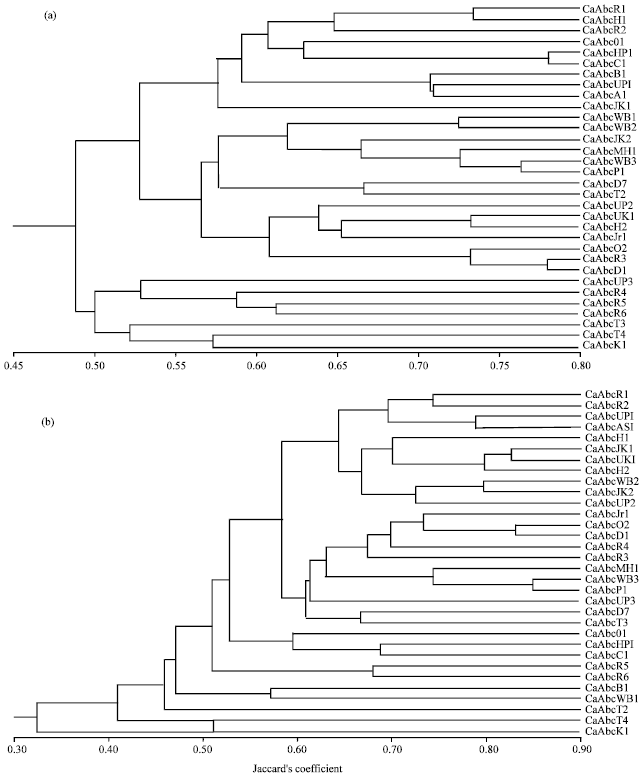 | |
| Fig. 1(a-b): | Dendrogram generated by the unweighted pair group method with arithmetic means (UPGMA) of Jaccard’s similarity coefficients based on the data amplified from the 32 isolates of Alternaria brassicicola using (a) RAPD and (b) ISSR primers |
The isolates CaAbcD1 and CaAbcR3 were the most closely related, while CaAbcR4 and CaAbcR1 were widely apart with similarity coefficient 0.78 and 0.35, respectively (Table 1).
Twelve ISSR primers yielded a total of 92 reproducible bands. Among these 92 loci only two were found to be monomorphic, whereas the rest were polymorphic. The Resolving Powers (RP) for ISSR were comparatively higher than the RAPD markers which proved ISSR a microsatellite based marker can produce extremely high variability and high mapping density. The highest RP was observed for the primer (AG)8AT (13.00) (Table 3, Fig. 1b). Jaccard’s similarity coefficient calculated from the ISSR data showed that all the isolates were correlated with each other with an average similarity of 0.53. The isolates CaAbcP1 and CaAbcWB3 were closely associated with similarity coefficient of 0.85 while the isolates CaAbcK1 and CaAbcH1 were most distantly separated having similarity coefficient of 0.22. In the present study, both RAPD and ISSR could successfully differentiate among the isolates of A. brassicicola showing high level of polymorphism. Gherbawy (2005) reported considerable genetic variability among Alternaria isolates, even within the same species group. This result was also well endorsed by findings of other species of Alternaria infecting crucifer (Sharma et al., 2013b; Meena et al., 2012; Sangwan and Mehta, 2007) and other crops (Kale et al., 2012). The phylogenetic relationship could not group the isolates based on geographical origin. Rather a high level of genetic variability among the studied A. brassicicola isolates was found using RAPD and ISSR markers. This result was similar to the findings of Bock et al. (2002) who got genetic differences among a small sample of isolates of A. brassicicola using AFLP markers and Bock et al. (2005) who detected substantial polymorphism between isolates from five populations of A. brassicicola attacking Cakile maritima along the New South Wales coast of Australia with a maximum of two genotypes being shared between population pairs. Varma et al. (2006) reported similar results while studying genetic diversity among the Alternaria solani isolates infecting tomato crop. The RAPD and ISSR analysis proved to be an efficient method for detecting genetic variability amongst different Alternaria spp.viz., Alternaria brassicae, A. brassicicola and A. raphani (Sharma and Tewari, 1998), A. cassiae and A. alternata (Tigano et al., 2003; Iram and Ahmad, 2005; Pusz, 2009), A. alternata, A. arborescens and A. tenuissima (Hong et al., 2005, 2006), A. solani (Varma et al., 2006; Kumar et al., 2008; Leiminger et al., 2010), A. radicina and A. carotiincultae (Park et al., 2008). Conserved primers ITS1 and ITS4, directed the amplification of an approximately 600 bp ITS rDNA fragments from all isolates of Alternaria brassicicola. DNA sequencing revealed that these fragments ranged in size from 584-624 bp. The rDNA revealed presence of partial sequences of 18S and 28S rRNA and complete sequences of ITS1, ITS2 and 5.8S rRNA gene. The GenBank accession numbers for all the amplified ITS sequences determined in this study are provided in Table 1. In addition to the sequences used in this study, the alignment included 15 ITS sequences of different Alternaria spp. (including A. brassicae (Accession: HQ674659), A. brassicicola clone Abra0318 (Accession: AY781078), A. brassicicola strain P4838 (Accession: GU983654), A. alternata (Accession: AB369904), A. carotiincultae (Accession: AF229465), A. mimicula (Accession: FJ266477), A. radicina (Accession: EU781949), A. japonica (Accession: AF229474), A. dianthi (Accession: AY154702), A. compacta (Accession: AF314573), A. gaisen (Accession: EU520078), A. mali (Accession: AY154683), A. citri (Accession: DQ339104), A. porri (Accession: AB026159), A. solani (Accession: HQ270462) obtained from GenBank. The sequence alignment revealed all the 32 A. brassicicola isolates are closely associated with each other with bootstrap value of 100% identity.
Neighbour-joining analysis of ITS sequences showed minor variations where the optimal tree was found with sum of branch length of 0.40. The neighbour-joining tree (Fig. 2a) was drawn to scale with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the maximum composite likelihood method, in the units of the number of base substitution per site. The analysis involved 47 nucleotide sequences containing the 32 A. brassicicola isolates along with other 15 Alternaria sp. from GenBank. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated. There were a total of 454 positions in the final dataset. All the 47 Alternaria spp. were grouped into one large clade in which 32 A. brassicicola isolates of the present study revealed 100% identity with the A. brassicicola clone Abra0318 (Accession: AY781078) and A. brassicicola strain P4838 (Accession: GU983654) obtained from Genbank. Moreover these isolates showed maximum similarity with the rDNA sequence of A. mimicula isolate from NCBI. The A. brassicicola isolates showed slight variation from two other species A. brassicae and A. japonica which were also highly responsible for causing leaf spot in crucifers, whereas the neighbour joining tree revealed A. japonica is more closely associated with the A. brassicicola rather than A. brassicae. Under the large Alternaria clade, distinct small species clades were revealed which include the other Alternaria species showing 95-100% identity with each other.
Parsimony analysis of ITS data set revealed 123 most parsimonious trees, out of which one tree (Fig. 2b) having length = 196, the consistency index = 0.642857 (0.595376), the retention index = 0.825871 (0.825871) and the composite index is 0.530917 (0.491703) for all sites and parsimony-informative sites (in parentheses) had near identical topology to the tree generated with neighbor-joining analysis (Fig. 2a).
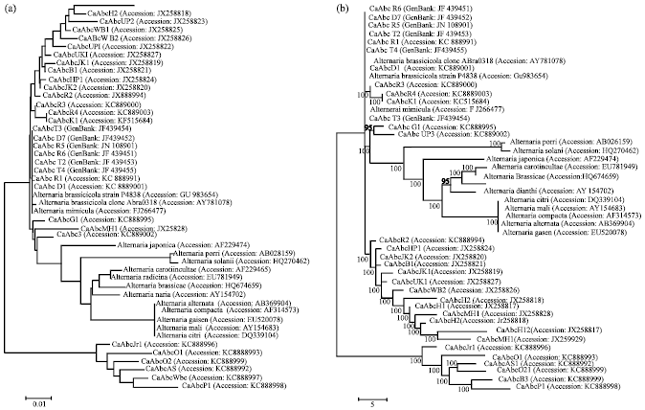 | |
| Fig. 2(a-b): | Evolutionary relationships among thirty two Alternaria brassicicola isolates of India by aligning rDNA sequences using neighbour joining method in MEGA5 software, (a) The phylogenetic tree having sum of branch length 0.40 in the unit of number of base substitutions per site is constructed using maximum composite likelihood method and (b) The phylogenetic tree was obtained using the close neighbour interchange algorithm with search level 0 in which the initial trees were obtained with the random addition of sequences (10 replicates). The analysis included 15 nucleotide sequences of other Alternaria spp. obtained from NCBI GenBank. The name of the isolates and Alternaria spp. along with the GenBank accession numbers are indicated on the termini of branches. The scale bar indicates the number of nucleotide substitutions. The consensus values (>50%) are mentioned below the branches |
The large Alternaria clade was again supported by a consensus value of 100% with all 32 A. brassicicola isolates having 100% identity with each other and the A. brassicicola isolates of NCBI.
The inter transcribed spacer region is widely used in classifying fungi (Chillali et al., 1998) due to its variability nature among species (Jung et al., 2002). This region is suitable for PCR amplification, restriction analysis and sequencing procedure (Pryor and Gilbertson, 2000). Molecular systematic studies among Alternaria spp. based on nuclear ribosomal DNA have been previously analysed (Kusaba and Tsuge, 1994, 1995; Pryor and Gilbertson, 2002; Chou and Wu, 2002; Pryor and Bigelow, 2003). Alternaria species pathogenic to crucifer varied from each other by rDNA sequence analysis (Jasalavich et al., 1995). In the present study, all the thirty two A. brassicciola isolates were found closely associated with each other with bootstrap value of 100% identity. Both the neighbor joining and parsimony tree analysis showed A. brassicicola isolates were forming a large group showing 95-100% similarity with other A. brassicicola isolates considered from NCBI. They were closely related to the A. mimicula and A. japonica species under the big Altermnaria clade. Pryor and Gilbertson (2000) also found 98% similarity A. brassicicola and A. japonica species. A. brassicae which is also majorly responsible for black leaf spot in cauliflower grouped distantly apart from A. brassicicola (Sharma et al., 2013b).
CONCLUSION
In conclusion the overall morphological, cultural, pathogenic and genetic variability showed high level of polymorphism among the A. brassicicola isolates infecting cauliflower in India with no clear grouping of isolates on the basis of their geographical origin. Morphological characteristics are correlated with the RAPD and ISSR analysis in showing polymorphism. Both the RAPD and ISSR primers proved to be efficient for detecting genetic variability among the A. brassicicola isolates. ITS analysis clearly indicates 99-100% similarity among the A. brassicicola isolates grouping them in a large group of species, different from other Alternaria spp. under the Alternaria genus/clade.
ACKNOWLEDGMENT
This study has been carried out with financial support from the Indian Council of Agricultural Research under the project Outreach programme on Diagnosis and management of leaf spot diseases of field and horticultural crops. The authors are grateful to Director, Indian Agricultural Research Institute and Head, Plant Pathology IARI for providing necessary facilities for the research.
REFERENCES
- Abul-Fazal, M., M.I. Khan and S.K. Saxena, 1994. The incidence of Alternaria species ID different cultivars of cabbage and cauliflower seeds. Indian Phytopathol., 47: 419-421.
Direct Link - Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Ansari, N.A., M.W. Khan and A. Muheet, 1989. Effect of some factors on growth and sporulation of Alternaria brassicae causing Alternaria blight of rapeseed and mustard. Acta Botanica Indica, 17: 49-53.
Direct Link - Bock, C.H., P.H. Thrall and J.J. Burdon, 2005. Genetic structure of populations of Alternaria brassicicola suggests the occurrence of sexual recombination. Mycol. Res., 109: 227-236.
CrossRefDirect Link - Bock, C.H., P.H. Thrall, C.L. Brubaker and J.J. Burdon, 2002. Detection of genetic variation in Alternaria brassicicola using AFLP fingerprinting. Mycol. Res., 106: 428-434.
CrossRefDirect Link - Chillali, M., H. Idder-Ighili, J.J. Guillaumin, C. Mohammed, B.L. Escarmant and B. Botton, 1998. Variation in the ITS and IGS regions of ribosomal DNA among the biological species of European Armillaria. Mycol. Res., 102: 533-540.
Direct Link - Cho, H.S., B.R. Kim and S.H. Yu, 2001. Taxonomic studies on Alternaria in Korea (1). Mycobiology, 29: 27-42.
Direct Link - Chou, H.H. and W.S. Wu, 2002. Phylogenetic analysis of internal transcribed spacer regions of the genus Alternaria and the significance of filament-beaked conidia. Mycol. Res., 106: 164-169.
CrossRefDirect Link - Connolly, A.G., I.D. Godwin, M. Cooper and I.H. DeLacy, 1994. Interpretation of randomly amplified polymorphic DNA marker data for fingerprinting sweet potato (Ipomoea batatas L.) genotypes. Theor. Applied Genet., 88: 332-336.
CrossRefDirect Link - Cooke, D.E.L., J.W. Forster, P.D. Jenkins, D.G. Jones and D.M. Lewis, 1998. Analysis of intraspecific and interspecific variation in the genus Alternaria by the use of RAPD-PCR. Ann. Applied Biol., 132: 197-209.
CrossRefDirect Link - David, J.C., 1991. IMI descriptions of fungi and bacteria: Set 108 nos. 1071-1080. Mycopathologia, 116: 45-64.
CrossRefDirect Link - Dillard, H.R., A.C. Cobb and J.S. Lamboy, 1998. Transmission of Alternaria brassicicola to cabbage by flea beetles (Phyllotreta cruciferae). Plant Dis., 82: 153-157.
CrossRefDirect Link - Gherbawy, Y.A.M.H., 2005. Genetic variation among isolates of Alternaria spp. from select Egyptian crops. Arch. Phytopathol. Plant Prot., 38: 77-89.
CrossRef - Goyal, P., M. Chahar, A.P. Mathur, A. Kumar and C. Chattopadhyay, 2011. Morphological and cultural variation in different oilseed Brassica isolates of Alternaria brassicae from different geographical regions of India. Indian J. Agric. Sci., 81: 1053-1058.
Direct Link - Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser., 41: 95-98.
Direct Link - Hodgkin, T. and M.V. MacDonald, 1986. The effect of a phytotoxin from Alternaria brassicicola on Brassica pollen. New Phytol., 104: 631-636.
CrossRefDirect Link - Hong, S.G., R.A. Cramer, C.B. Lawrence and B.M. Pryor, 2005. Alt a 1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol., 42: 119-129.
CrossRefDirect Link - Hong, S.G., M. Maccaroni, P.J. Figuli, B.M. Pryor and A. Belisario, 2006. Polyphasic classification of Alternaria isolated from hazelnut and walnut fruit in Europe. Mycol. Res., 110: 1290-1300.
CrossRefDirect Link - Humpherson-Jones, F.M., 1985. The incidence of Alternaria spp. and Leptosphaeria maculans in commercial brassica seed in the United Kingdom. Plant Pathol., 34: 385-390.
CrossRefDirect Link - Iram, S. and I. Ahmad, 2005. Analysis of variation in Alternaria alternata by pathogenicity and RAPD study. Pol. J. Microbiol., 54: 13-19.
PubMed - Jasalavich, C.A., V.M. Morales, L.E. Pelcher and G. Seguin-Swartz, 1995. Comparison of nuclear ribosomal DNA sequences from Alternaria species pathogenic to crucifers. Mycol. Res., 99: 604-614.
CrossRefDirect Link - Jung, D.S., Y.J. Na and K.H. Ryu, 2002. Phylogenic analysis of Alternaria brassicicola producing bioactive metabolites. J. Microbiol., 40: 289-294.
Direct Link - Kale, S.M., V.C. Pardeshi, G.S. Gurjar, V.S. Gupta, R.T. Gohokar, P.B. Ghorpade and N. Kadoo, 2012. Inter-simple sequence repeat markers reveal high genetic diversity among Alternaria alternata isolates of Indian origin. J. Mycol. Plant Pathol., 42: 194-200.
Direct Link - Kolte, S.J., R.P. Awasthi and Vishwanath, 1987. Assessment of yield losses due to Alternaria blight in rapeseed and mustard. Indian Phytopathol., 40: 209-211.
Direct Link - Kubota, M., K. Abiko, Y. Yanagisawa and K. Nishi, 2006. Frequency of Alternaria brassicicola in commercial cabbage seeds in Japan. J. Gen. Plant Pathol., 72: 197-204.
CrossRefDirect Link - Kumar, R. and P.P. Gupta, 1994. Survival of Alternaria brassicae, A. brassicicola and A. alternata in the seeds of mustard (B. juncea) at different temperatures and relative humidities. Ann. Biol., 10: 55-58.
Direct Link - Kumar, V., S. Haldar, K.K. Pandey, R.P. Singh, A.K. Singh and P.C. Singh, 2008. Cultural, morphological, pathogenic and molecular variability amongst tomato isolates of Alternaria solani in India. World J. Microbiol. Biotechnol., 24: 1003-1009.
CrossRefDirect Link - Kusaba, M. and T. Tsuge, 1994. Nuclear ribosomal DNA variation and pathogenic specialization in Alternaria fungi known to produce host-specific toxins. Applied Environ. Microbiol., 60: 3055-3062.
Direct Link - Kusaba, M. and T. Tsuge, 1995. Phologeny of Alternaria fungi known to produce host-specific toxins on the basis of variation in internal transcribed spacers of ribosomal DNA. Curr. Genet., 28: 491-498.
CrossRefDirect Link - MacKinnon, S.L., P. Keifer and W.A. Ayer, 1999. Components from the phytotoxic extract of Alternaria brassicicola, a black spot pathogen of canola. Phytochemistry, 51: 215-221.
CrossRefDirect Link - Maude, R.B. and F.M. Humpherson-Jones, 1980. Studies on the seed-borne phases of dark leaf spot Alternaria brassicicola and grey leaf spot Alternaria brassicae of brassicas. Ann. Applied Biol., 95: 311-319.
CrossRefDirect Link - Maude, R.B., F.M. Humpherson-Jones and C.G. Shuring, 1984. Treatments to control Phoma and Alternaria infections of brassica seeds. Plant Pathol., 33: 525-535.
CrossRefDirect Link - Meena, P.D., A. Rani, R. Meena, P. Sharma, R. Gupta and P. Chowdappa, 2012. Aggressiveness, diversity and distribution of Alternaria brassicae isolates infecting oilseed Brassica in India. Afr. J. Microbiol. Res., 6: 5249-5258.
Direct Link - Milgroom, M.G. and W.E. Fry, 1997. Contributions of population genetics to plant disease epidemiology and management. Adv. Bot. Res., 24: 1-30.
CrossRefDirect Link - Morris, P.F., M.S. Connolly and D.A. St Clair, 2000. Genetic diversity of Alternaria alternata isolated from tomato in California assessed using RAPDs. Mycol. Res., 104: 286-292.
Direct Link - Nei, M. and S. Kumar, 2000. Molecular Evolution and Phylogenetics. Oxford University Press, United Kingdom, ISBN-13: 9780195350517, Pages: 352.
Direct Link - Oliver, E.J., P.H. Thrall, J.J. Burdon and J.E. Ash, 2001. Vertical disease transmission in the Cakile-Alternaria host-pathogen interaction. Aust. J. Bot., 49: 561-569.
CrossRefDirect Link - Park, M.S., C.E. Romanoski and B.M. Pryor, 2008. A re-examination of the phylogenetic relationship between the causal agents of carrot black rot, Alternaria radicina and A. Carotiincultae. Mycologia, 100: 511-527.
CrossRefDirect Link - Pattanamahakul, P. and R.N. Strange, 1999. Identification and toxicity of Alternaria brassicicola, the causal agent of dark leaf spot disease of Brassica species grown in Thailand. Plant Pathol., 48: 749-755.
CrossRefDirect Link - Prevost, A. and M.J. Wilkinson, 1999. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet., 98: 107-112.
CrossRefDirect Link - Pryor, B.M. and D.M. Bigelow, 2003. Molecular characterization of Embellisia and Nimbya species and their relationship to Alternaria, Ulocladium and Stemphylium. Mycologia, 95: 1141-1154.
Direct Link - Pryor, B.M. and R.L. Gilbertson, 2000. Molecular phylogenetic relationships amongst Alternaria species and related fungi based upon analysis of nuclear ITS and mt SSU rDNA sequences. Mycol. Res., 104: 1312-1321.
CrossRefDirect Link - Pryor, B.M. and R.L. Gilbertson, 2002. Relationships and taxonomic status of Alternaria radicina, A. carotiincultae and A. petroselini based upon morphological, biochemical and molecular characteristics. Mycologia, 94: 49-61.
PubMedDirect Link - Pusz, W., 2009. Morpho-physiological and molecular analyses of Alternaria alternata isolated from seeds of Amaranthus. Phytopathologia, 54: 5-14.
Direct Link - Reis, A. and L.S. Boiteux, 2010. Alternaria species infecting brassicaceae in the Brazilian neotropics: Geographical distribution, host range and specificity. J. Plant Pathol., 92: 661-668.
Direct Link - Roberts, R.G., S.T. Reymond and B. Andersen, 2000. RAPD fragment pattern analysis and morphological segregation of small-spored Alternaria species and species groups. Mycol. Res., 104: 151-160.
Direct Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Sangwan, M.S. and N. Mehta, 2007. Pathogenic variability in isolates of Alternaria brassicae (Berk.) Sacc. from different agro-climatic zones of India. Plant Dis. Res., 22: 101-107.
Direct Link - Sharma, M., S. Deep, D.S. Bhati, P. Chowdappa, R. Selvamani and P. Sharma, 2013. Morphological, cultural, pathogenic and molecular studies of Alternaria brassicae infecting cauliflower and mustard in India. Afr. J. Microbiol. Res., 7: 3351-3363.
Direct Link - Sharma, P., S. Deep, M. Sharma and D.S. Bhati, 2013. Genetic variation of Alternaria brassicae (Berk.) Sacc., causal agent of dark leaf spot of cauliflower and mustard in India. J. Gen. Plant Pathol., 79: 41-45.
CrossRef - Sharma, P., S. Deep, M. Sharma, D. Singh and P. Chowdappa, 2013. PCR based assay for the detection of Alternaria brassicicola in Crucifers. Indian Phytopathol., 66: 263-268.
Direct Link - Sharma, P., S.R. Sharma and M. Sindhu, 2004. A detached leaf technique for evaluation of resistance in cabbage and cauliflower against three major pathogens. Indian Phytopathol., 57: 315-318.
Direct Link - Sharma, T.R. and J.P. Tewari, 1995. Detection of genetic variation in Alternaria brassicae by RAPD fingerprints. J. Plant Biochem. Biotechnol., 4: 105-107.
CrossRefDirect Link - Sharma, T.R. and J.P. Tewari, 1998. RAPD analysis of three Alternaria species pathogenic to crucifers. Mycol. Res., 102: 807-814.
Direct Link - Singh, D., R. Singh, H. Singh, R.C. Yadav and N. Yadav et al., 2007. Cultural and morphological variability in Alternaria brassicae isolates of Indian mustard (Brassica juncea L. Czern and Coss.). Proceeding of the 12th International Rapeseed Congress, Volume 4, March 26-30, 2007, Wuhan, China, pp: 158-160.
- Singh, R., C. Mohan and V.K. Sharma, 2012. Morphological and pathological variations in isolates of Alternaria brassicae causing leaf blight of rapeseed and mustard. Plant Dis. Res., 27: 22-27.
Direct Link - Sofi, T.A., M.A. Beig, G.H. Dar, M. Ahmad and A. Hamid et al., 2013. Cultural, morphological, pathogenic and molecular characterization of Alternaria mali associated with Alternaria leaf blotch of apple. Afr. J. Biotechnol., 12: 370-381.
Direct Link - Sun, X. and T.Y. Zhang, 2008. Morphological and molecular characterization of Alternaria isolates on fruits of Pyrus bretschneideri Rehd Ya Li. Mycosystema, 27: 105-117.
Direct Link - Tamura, K., M. Nei and S. Kumar, 2004. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. U.S.A., 101: 11030-11035.
CrossRefDirect Link - Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei and S. Kumar, 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance and maximum parsimony methods. Mol. Biol. Evol., 28: 2731-2739.
CrossRefPubMedDirect Link - Tigano, M.S., S. Aljanabi and S.C.M. De Mello, 2003. Genetic variability of Brazilian Alternaria spp. isolates as revealed by RAPD analysis. Braz. J. Microbiol., 34: 117-119.
CrossRef - Varma, P.K., S. Singh, S.K. Gandhi and K. Chaudhary, 2006. Variability among Alternaria solani isolates associated with early blight of tomato. Commun. Agric. Applied Biol. Sci., 71: 37-46.
PubMedDirect Link - Weir, T.L., D.R. Huff, B.J. Christ and C.P. Romaine, 1998. RAPD-PCR analysis of genetic variation among isolates of Alternaria solani and Alternaria alternata from potato and tomato. Mycologia, 90: 813-821.
Direct Link - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link