
Hancheng Wang
Key Laboratory of Molecular Genetics, Guizhou Academy of Tobacco Sciences, Guiyang, 550081, People`s Republic of China
Jin Wang
College of Life Science, Yangtze University, Jingzhou, 434025, People`s Republic of China
Wenhong Li
Guizhou Institute of Plant Protection, Guiyang, 550006, People`s Republic of China
Jiehong Zhao
Key Laboratory of Molecular Genetics, Guizhou Academy of Tobacco Sciences, Guiyang, 550081, People`s Republic of China
Maosheng Wang
Key Laboratory of Molecular Genetics, Guizhou Academy of Tobacco Sciences, Guiyang, 550081, People`s Republic of China
Ning Lu
Key Laboratory of Molecular Genetics, Guizhou Academy of Tobacco Sciences, Guiyang, 550081, People`s Republic of China
Yushuang Guo
Key Laboratory of Molecular Genetics, Guizhou Academy of Tobacco Sciences, Guiyang, 550081, People`s Republic of China
Changqing Zhang
College of Agriculture, Yangtze University, Jingzhou, 434025, People`s Republic of China
Plant Pathology Journal
Year: 2014 | Volume: 13 | Issue: 3 | Page No.: 177-185







ABSTRACT
Tobacco black shank caused by Phytophthora nicotianae is one of the most destructive diseases suffered by tobacco in China. Streptomycin, calcium oxide and synthetic fertilizer are three important non-target chemicals used frequently during tobacco growing period. This study has evaluated the activities of the non-target chemicals streptomycin, calcium oxide and synthetic fertilizer on the mycelial growth, sporulation, zoospore formation and germination of cystospores of P. nicotianae. Metabolic fingerprints of P. nicotianae under pressures of those three chemicals are also compared with control treatments. Streptomycin inhibited mycelial growth, sporangia production, zoospore formation and cystospore germination more effectively than the other chemicals tested. Calcium oxide inhibited sporangia production, zoospore formation and cystospore germination at much higher concentration. Mycelial growth of P. nicotianae was not affected by this chemical. Synthetic fertilizer had negative or no effect on sporangia production and no effect on mycelial growth. Zoospore formation and cystospore germination was inhibited at high concentrations of this chemical. Metabolic fingerprint of P. nicotianae was not or poorly affected by calcium oxide and synthetic fertilizer. Streptomycin significantly changed the fingerprint of the pathogen, substrate of y-aminobutyric acid, fumaric acid, L-alaninamide, L-alanyl-glycine, L-proline, L-pyroglutamic acid and putrescine cannot be utilized by P. nicotianae. So it is no use to add calcium oxide to soil for black shank management in the field, while is useful for streptomycin. Synthetic fertilizer in some case may enhance the development of the disease. These new findings provide important information for black shank management in the future.
PDF Abstract XML References Citation
Received: April 17, 2014;
Accepted: July 23, 2014;
Published: August 09, 2014
How to cite this article
Hancheng Wang, Jin Wang, Wenhong Li, Jiehong Zhao, Maosheng Wang, Ning Lu, Yushuang Guo and Changqing Zhang, 2014. Activities of Three Agrochemicals against Phytophthora parasitica var. nicotianae and its Metabolic Fingerprints under Chemical Pressures. Plant Pathology Journal, 13: 177-185.
DOI: 10.3923/ppj.2014.177.185
URL: https://scialert.net/abstract/?doi=ppj.2014.177.185
DOI: 10.3923/ppj.2014.177.185
URL: https://scialert.net/abstract/?doi=ppj.2014.177.185
INTRODUCTION
Phytophthora parasitica var. nicotianae (P. nicotianae) Breda de Haan causes black shank of tobacco (Nicotiana tabacum L.) in most tobacco growing regions of the world. This pathogen has a host range that includes more than 72 species in the Solanaceae (Hickmann, 1958) and is a particularly important pathogen of tobacco (Dukes and Apple, 1961; Stokes and Litton, 1966; McCarter, 1967; Jacobi et al., 1983). In China, this disease is managed through the use of resistant cultivars, crop rotation, cultural practices and the effective systemic compound metalaxyl (Yuan et al., 2006). However, due to high aggressiveness and increasing resistance to metalaxyl of this pathogen (Cartwright and Spurr Jr., 1998; Wang et al., 2013a), it is still a serious problem for farmers in China.
China accounts for more than 39.6% of the total global tobacco production (Wang et al., 2013b). During tobacco production proceeding, farmers normally add calcium oxide to the soil to raise soil pH to the ideal range for tobacco production and use tobacco specific synthetic fertilizer with all three N: P: K plant nutrients (10:10:25) to provide nutrition for flue-cured tobacco plants (Table 1).
| Table 1: | Use dosage of three agrochemicals in flue-cured tobacco production seasons in Southwest China |
 | |
Meanwhile, another serious soil-borne disease of tobacco bacterial wilt, caused by Ralstonia solanacearum, is happened frequently together with black shank in most regions throughout China and also quite difficult to control. The antibiotic streptomycin has been frequently added to tobacco soil for bacterial wilt management (Hu, 2009).
During the lifecycle and epidemiology of P. nicotianae in the period of tobacco production, many factors could affect the infection of the pathogen and influence the disease development. Previous studies showed that black shank occurs in both acid and alkaline soils with optimum pH value for disease development between 6 and 7 (Kong et al., 2009). Calcium levels are positively correlated with severity of black shank disease (Von Broembsen and Deacon, 1997). However, whether the effect of calcium is on the oomycete or the host of tobacco is unclear. Mitigation strategies against these soil pathogens have primarily relied on metalaxyl and streptomycin. So far, limited information actually outlines the activities of calcium oxide, synthetic fertilizer and streptomycin to soil-borne pathogen of P. nicotianae. Lack of evidence for dosage use of calcium oxide and fertilizer guides black shank management in the field. Recently, the Biolog FF MicroPlate was introduced for characterization of filamentous fungi (FF MicroPlateTM Instruction for use). The MicroPlate contains 95 discrete substrates that could be used to test the substrate utilization and metabolite profiling of the pathogen P. nicotianae under different chemical pressures, which could help to know the metabolic activity of P. nicotianae when those chemicals were utilized in the field.
The objectives of this study were: (1) To investigate in vitro activities of three agrochemicals (streptomycin, calcium oxide and synthetic fertilizer) against P. nicotianae and (2) To evaluate the carbon metabolic fingerprint of P. nicotianae under different pressures of those chemicals. The compounds were chosen due to their importance for tobacco production in the field. The outcome of this study is be much valuable to assess the effect of non-target chemicals against the pathogen of P. nicotianae.
MATERIALS AND METHODS
Pathogen, media and chemical preparation: One isolate of P. nicotianae with wild-type sensitivity and pathogenicity to tobacco, was selected as the specie for assay in this study. The isolate was collected, isolated and identified from our early study (Wang et al., 2013a) in 2011 from an infected tobacco stem in a tobacco commercial field of the Guizhou province of China. Lima Bean Agar (LBA) was prepared by boiling 60 g lima bean for 1 h and filtering through 4 layers of gauze. The filtrate was added to deionized water and agar powder (16.0 g L-1) to a final volume of 1.0 L. Ten percent V8 liquid medium was prepared by adding 100 mL Campbell’s V8 juice to 900 mL sterile water containing 0.2 g CaCO3. All culture media were autoclaved prior to use. Stock chemical solutions were prepared by dissolving technical grade streptomycin (Sigma China Co., Ltd, Shanghai, China), technical grade calcium oxide (a.i. 98%; Jinfu Co., Ltd, Fuquan, China) and tobacco commercial grade synthetic fertilizer (a.i. 45%; N: P: K = 10: 10: 15; Jinfu Co., Ltd, Fuquan, China) in distilled water. Solutions were diluted as required and stored at 4°C to preserve chemical activity in the dark. Chemicals were added to LBA after autoclaving when the agar had cooled to approximately 50°C.
Sensitivities to mycelial growth and effect on morphology of hyphae: Sensitivities of mycelial growth to chemicals were assayed on LBA amended with different concentrations of tested compounds previously dissolved in distilled water. The final concentrations tested were for streptomycin 0, 1, 10, 100 and 200 mg L-1, for calcium oxide 0, 250, 500, 1000 and 2000 mg L-1 and for synthetic fertilizer 0, 31.25, 62.5, 125, 250 and 500 mg L-1. For each chemical at each concentration, three replicates were used. The diameters of the colonies were measured after incubation for 6 days at 25°C in darkness. The experiment was conducted twice.
The effect of chemicals on hyphal morphology was visualized as described previously (Kuhn et al., 1991). Briefly, LBA agar amended with test compounds was overlaid with a cellophane membrane (Bio-Rad Laboratories, Inc.), inoculated with a mycelia plug and incubated for 5 days at 25°C in darkness. Under these conditions, a relatively sparse hyphal monolayer was produced which could be observed directly under the light microscope.
Sensitivities to sporangia production: Sensitivities of sporangia production was assayed according to a previously described method for quantification of sporangia (Sun et al., 2010). Three individual agar discs (10 mm diameter) were removed from the edge of an actively growing culture and placed face up on the centre of a petri dish (9 cm diametere) containing 15 mL of 10% V8 liquid medium amended with a test chemical at various concentrations. The final concentrations tested were for streptomycin 0, 0.1, 1, 10 and 100 mg L-1, for calcium oxide 0, 206, 413, 825 and 1650 mg L-1 and for synthetic fertilizer 0, 63, 125, 250 and 500 mg L-1. Afterwards, the petri dishes were incubated at 25°C in light for 96 h, then the plugs were placed on a glass slide and the number of sporangia was counted at three sites along the margins of each agar plug with the help of a microscope. For each chemical at each concentration, three replicates were conducted. The experiment was conducted twice.
Sensitivities to zoospore formation and cystospore germination: Sporangia of P. nicotianae were induced on 10% V8 plates as mentioned above, the suspension was filtered through a double-layer of sterile cheesecloth to remove mycelial fragments and diluted with sterile water to a final concentration of 1x105 sporangia mL-1. Aqueous preparations of each chemical at different concentrations were added to an equal volume (500 μL) of the sporangia suspension. A control sporangia suspension received an equal volume of water only. The final concentrations tested for streptomycin, calcium oxide and synthetic fertilizer were for streptomycin 0, 0.1, 1, 10 and 100 mg L-1, for calcium oxide 0, 206, 413, 825 and 1650 mg L-1 and for synthetic fertilizer 0, 63, 125, 250 and 500 mg L-1. Sporangia suspensions were maintained at 4°C for 0.5 h and then at 25°C for 2 h in darkness to release the zoospores. A sporangium was regarded as released if the sporangium was empty which was easily to determine under the microscope. Empty sporangia were quantified on microscope slides at three sites by counting 100 sporangia per site and the percentages of released sporangia were calculated.
For sensitivity of the compounds on the germination of cystospores, zoospore suspensions of P. nicotianae were prepared as described above and diluted to a final concentration of 1x105 zoospores mL-1. Aqueous preparations of each compound at different concentrations were mixed with equal volume (500 μL) of the zoospore suspension. A control zoospore suspension was diluted with an equal volume of water only. The final concentrations tested for streptomycin were 0, 0.1, 1, 10 and 100 μg mL-1, for calcium oxide 0, 206, 413, 825 and 1650 μg mL-1 and for synthetic fertilizer 0, 62.5, 125, 250 and 500 μg mL-1. Afterwards, zoospore suspension were incubated at 25°C in darkness for 10 h. A cystospore was scored as germinated if the germ tube had reached at least the length of the cystospore. Germinated cystospores were quantified with the help of a microscope at three sites by counting 100 cystospores per site. The experiment was conducted twice with three replications.
Carbon metabolic fingerprint of P. nicotianae under pressure of chemicals: Carbon metabolic fingerprints of P. nicotianae under different pressures of the chemicals were conducted by the Biolog system (Biolog Inc., Hayward, CA). In preparation for analysis, zoospores of P. nicotianae were prepared as described above and diluted with sterile water to a final concentration of 1x105 sporangia mL-1. Streptomycin, calcium oxide and synthetic fertilizer were added into Filamentous Fungi Inoculating Fluid (FF-IF) to get a final concentration of 30, 1650 and 2000 μg mL-1, respectively. The FF MicroPlate test panels, containing 95 carbon source utilization assays, were used for analysis according to the manufacturer’s instructions throughout the experiment (Rice and Currah, 2005; Druzhinina et al., 2006; Singh, 2009; Papaspyridi et al., 2011). Readings were recorded for 6 days and kinetic data was analyzed with OmniLog-PM software. This software generates time course curves for respiration and calculates differences in the areas for test pathogen under different chemical pressures. This experiment was conducted twice.
Statistical analysis: Results from repeated experiments were combined for analysis since variances between experiments were homogeneous. All data was processed with the SIGMASTA Statistical Software Package (SPSS Science, Ver. 11). The concentration of each chemical causing 50% (EC50) or 90% (EC90) reduction in mycelial growth, sporulation, inhibition of zoospore release and germination of cystospores compared to the absence of the compound was estimated from the fitted regression line of the log-transformed percentage inhibition plotted against the log-transformed compound concentration (Brandt et al., 1988).
RESULTS
Sensitivities to mycelial growth and effect on morphology of hyphae: Inhibition of mycelial growth was highest with streptomycin with EC50 and EC90 values of 8.29 and 24.78 μg mL-1 (Table 2), respectively. In comparison, mycelial growth of P. nicotianae was less affected by calcium oxide and synthetic fertilizer in the test system on LBA plates with EC50 value >500 μg mL-1 for each chemical (Fig. 1). Moreover, in the absence of a compound, the average mycelial growth for P. nicotianae on LBA plates after 6 days at 25°C was about 65 mm.
Streptomycin at the final test concentration of 100 μg mL-1 significantly inhibited the colony growth of P. nicotianae. It also induced major changes in the morphology of hyphae and the most characteristic effect of streptomycin was the development of periodic swellings along the hyphae (Fig. 2).
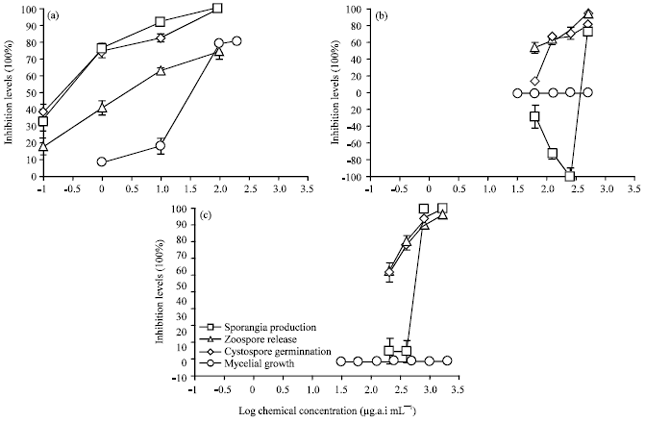 | |
| Fig. 1(a-c): | Influence of dose of (a) Streptomycin, (b) Synthetic fertilizer and (c) Calcium oxide on inhibition of mycelia growth, sporangia production, zoospore release and cystospore germination. |
| Table 2: | EC50 and EC90 values of three agrochemicals for mycelial growth, sporangia production, zoospore formation and germination of cystospores of Phytophthora nicotianaea |
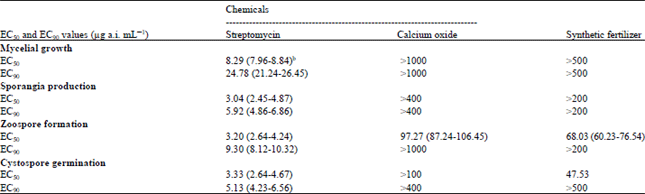 | |
| aEC50 and EC90 values are the concentrations of each chemical causing 50 and 90% reduction, respectively, in mycelial growth, sporangia production, zoospore formation and germination of cystospores of Phytophthora nicotianae compared to the absence of a chemical. bMeans values in probability level | |
In comparison, the morphology of the hyphae of the pathogen was not affected by calcium oxide and synthetic fertilizer at their highest test concentrations.
Sensitivities to sporangia production: Among the three test compounds, streptomycin exhibited highest activity on sporangia production with EC50 and EC90 values of 3.04 and 5.92 μg mL-1 (Table 2), respectively. Calcium oxide and synthetic fertilizer presented lower inhibition on sporangia production with each EC50 values >200 μg mL-1. Additionally, calcium oxide completely inhibited sporangia production when treated at concentration of > 800 μg mL-1 Fig. 1c. Synthetic fertilizer enhanced sporangia production at the concentrations between 62.5 and 250 μg mL-1 Fig. 1b.
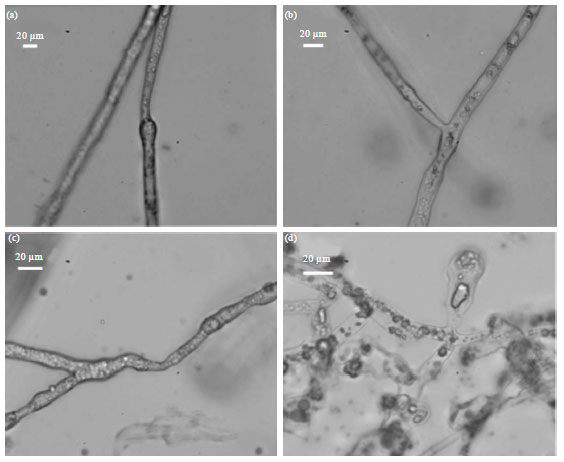 | |
| Fig. 2(a-d): | Morphology of cell materials in the hyphae of Phytophthora nicotianae after exposure to various concentrations of streptomycin. The bar markers indicate 20 μm in all 4 parts with (a) No streptomycin (b) 1 μg mL-1 (c) 10 μ mL-1 and (d) 100 μ mL-1 streptomycin |
Reduction of sporangia production in of streptomycin at 1 μg mL-1 was significantly higher than that recorded for 400 μg mL-1 of calcium oxide and 500 μg mL-1 of synthetic fertilizer. Production of sporangia was totally suppressed by 100 μg mL-1 of streptomycin. The average number of sporangia developing on V8 media plate in the absence of a chemical after 96 h inoculation at 25°C was about 2.5x102 per site.
Sensitivities to zoospore formation and cystospore germination: Among the three chemicals tested, streptomycin was the most sensitive on zoospore formation with EC50 and EC90 values of 3.20 and 9.30 μg mL-1, respectively (Table 2). Followed by synthetic fertilizer with EC50 and EC90 values of 68.03 and >200 μg mL-1, respectively. In comparison, calcium oxide was least effective with EC50 and EC90 values of 97.27 and >1000 μg mL-1, respectively.
Germination of cystospores of P. nicotianae was seriously inhibited by streptomycin with EC50 and EC90 values of 3.33 and 5.13 μg mL-1, respectively. Less inhibition was noted for calcium oxide and synthetic fertilizer with EC50 and EC90 values of >100, 47.53 μg mL-1 and >400 and >500 μg mL-1, respectively. Complete inhibition of cystospore germination was achieved with 100 μg mL-1 streptomycin (Fig. 1).
Carbon metabolic fingerprint of P. nicotianae under pressure of chemicals: Carbon metabolic fingerprint of P. nicotianae was conducted by the Biolog system. When incubated on FF MicroPlate (Fig. 3) without chemicals, P. nicotianae utilized 47 carbons among the 95 carbon source utilization assays (Fig. 4). The typical carbons metabolized by P. nicotianae included D-gluconic acid, D-glucuronic acid, 2-Keto-D-gluconic acid, D-xylose, fumaric acid, L-lactic acid, D-malic acid, L-malic acid, quinic acid, D-saccharic acid, succinic acid, L-alanine, L-alanyl-glycine, L-asparagine, L-aspartic acid, L-glutamic acid, L-proline, L-pyroglutamic acid and L-serine. In comparison, when incubated under the pressure of streptomycin at 30 μg mL-1, only 27 carbons were metabolized by the pathogen.
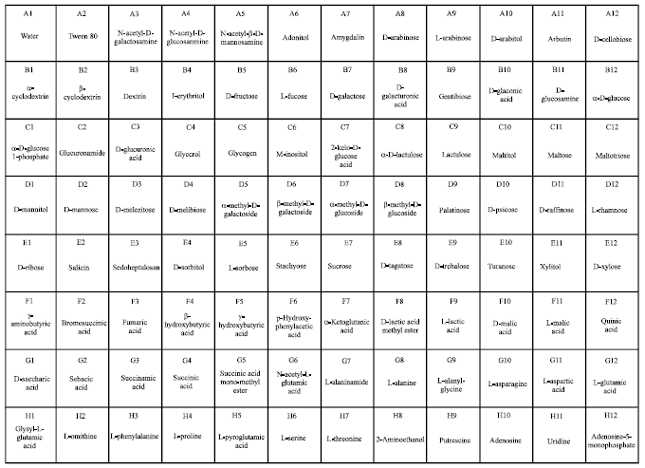 | |
| Fig. 3: | Layout of assays in the Biolog FF MicroPlate |
Streptomycin significantly inhibited the utilization of y-aminobutyric acid, fumaric acid, L-alaninamide, L-alanyl-glycine, L-proline, L-pyroglutamic acid and putrescine by the pathogen (Fig. 4a). When treated with synthetic fertilizer at final test concentration of 500 μg mL-1, P. nicotianae had similar metabolic fingerprint as that of control, except the carbon of bromosuccinic acid (Fig. 4b). Additionally, when treated with calcium oxide at final test concentration of 1650 μg mL-1, the pathogen had the same metabolic fingerprint as that of control (Fig. 4c).
DISCUSSION
To establish an efficient disease management strategy for the control of tobacco black shank it is important to know the interaction between environmental factors and the pathogen of P. nicotianae. As a first step, the activities of agrochemicals on the different life cycle stages have to be established. While some chemicals might inhibit some stages of pathogen development (e.g., by inhibition mycelia growth), others may enhance the epidemic of black shank (e.g., stimulate sporulation). Current analysis in this study has shown the strengths and weaknesses of three major agrochemicals during the various development stages of P. nicotianae and also carbon metabolic fingerprint of this pathogen under pressures of those chemicals.
Among the agrochemicals tested, the mycelial growth, the sporangia production, zoospore formation and the germination of cystospores of P. nicotianae was more sensitive to streptomycin than that of calcium oxide and synthetic fertilizer. Corresponding findings have been previously reported for streptomycin in Phytophthora, Pythium and Peronophythora species (Eckert and Tsao, 1962; Rooke and Shattock, 1983; Liu, 2001; Lee et al., 2005). In our test system, highest sensitivity was noted by sporangia formation, followed by that of cystospores germination and zoospore formation and less sensitivity was accorded for mycelial growth. Though in certain cases, toxicity may be different due to discrepancies between culture media, test concentrations, time of dosage and duration of contact with the pathogen. Results from this study still provided useful information for our understanding the activity of streptomycin against P. nicotianae.
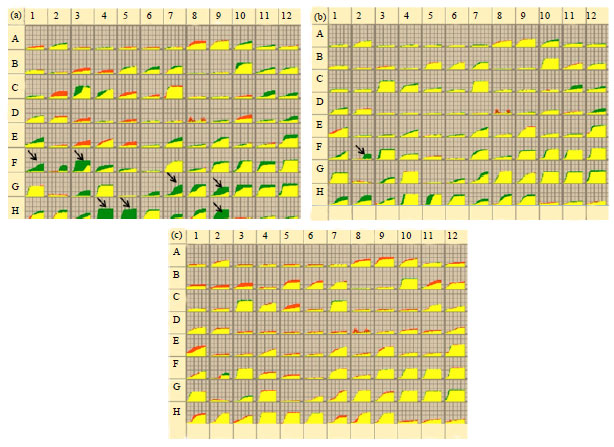 | |
| Fig. 4(a-c): | Carbon metabolic fingerprint comparisons of P. nicotianae between control treatment and three agronomic chemical treatments. Comparison between control and (a) 30 μg mL-1 of streptomycin, (b) 500 μg mL-1 of synthetic fertilizer and (c) 1650 μg mL-1 of calcium oxide. Green presented metabolic fingerprint of control treatments; red in (a), (b) and (c) presented metabolic fingerprint of streptomycin, synthetic fertilizer and calcium oxide, respectively; yellow presented common fingerprints of both control and chemical treatments |
Streptomycin, an aminocyclitol glycoside antibiotic, was first discovered in the 1950s (Sundin and Bender, 1993) and then used to control bacterial pathogens of plants, including tobacco bacterial wilt (caused by R. solanacearum) and tobacco wildfire (caused by Pseudomonas syringae pv. tabaci). Its mode of action to bacteria is presumably the binding to the 30S ribosomal subunit, thereby interfering with polypeptide synthesis and thus inhibiting translation (Sundin, 2002). While activities of streptomycin have been investigated extensively to oomycete pathogens (Rooke and Shattock, 1983; Liu, 2001; Lee et al., 2005), the exact mechanism by which streptomycin influences Phytophthora species is seldom reported (Chabane et al., 1996; Whittaker et al., 1996). After exposure to streptomycin, zoospores of P. nicotianae were able to switch into cystospores accompanied with the formation of a cell wall, whereas cystospores did not produce germ tubes when treated at concentration of 100 μg mL-1. In streptomycin-treated hyphae, the most characteristic change was the development of periodic swelling and the disruption of tip growth. Further research is needed to know the distribution of the newly synthesized cell wall materials in streptomycin-treated hyphae to get the mode of action of this chemical to oomycete pathogens.
Tobacco in southwest China is mainly located in semi-arid areas over acid or neutral soils. The CaO and CaCO3 are sometimes applied in the soil to enhance botanical composition and production of flue-cured tobacco. For some soilborne pathogens (Von Broembsen and Deacon, 1997; Campanella et al., 2002; Heyman et al., 2007; Serrano et al., 2012), extracellular calcium is necessary for production of zoosporangia by Phytophthora spp. (Halsall and Forrester, 1977), for adhesion of Phytophthora zoospore cysts to host or other surfaces (Gubler et al., 1989; Donaldson and Deacon, 1992) and for germination of Phytophthora (Von Broembsen and Deacon, 1996). It had also demonstrated that Ca2+ interfere with P. nicotianae zoospore biology at multiple stages with compounding effects on epidemiology and management of the pathogen (Halsall and Forrester, 1977; Von Broembsen and Deacon, 1997). In comparison, in our test system, sensitivity of P. nicotianae from tobacco to CaO agreed with the data reported in P. cinnamomi (Erwin and Ribeiro, 1996), P. nicotianae (Campanella et al., 2002), P. nicotianae (Von Broembsen and Deacon, 1997) and P. sojae (Sugimoto et al., 2005). Further research is needed to confirm the ability of calcium oxide to enhance black shank management by field trials. Additionally, to our best knowledge, few researches have been conducted to the fungicidal value of synthetic fertilizer to phytophthora oomycete. In our test system, while high concentrations of synthetic fertilizer produced an inhibition on zoospore release and cystospore germination it presented a direct negative effect on mycelial growth and sporangia production of the pathogen. Synthetic fertilizer could enhance the epidemic of black shank in the field; assays in planta trial should be conducted to investigate the relationship between fertilizer and disease incidence.
Phytophthora pathogens are known to have unique biochemical pathways to assimilate a vast array of simple and complex nutrients available to them. In our study, the Biolog FF MicroPlate which had 95 discrete substrates, was introduced to test the metabolites of P. nicotianae under three agronomic chemical pressures (Singh, 2009). Culture growth of the pathogen was monitored by change in absorbance in each well. Fingerprints were created with Spotfire visualization software, data of each well were collected automatically during incubation and variations between control and treatments were compared. Under the test pressure of calcium oxide, P. nicotianae shared the same fingerprints as that of control. Under nutrient replete conditions, only the utilization of the utilization of the artificial substrate bromosuccinic acid was affected. Those finding agreed with the results gotten in vitro for the two chemicals. So it is no use to add calcium oxide and synthetic fertilizer to soil for tobacco black shank management in the field. In contrast, the addition of streptomycin inhibited the ability to utilize several substrates including y-aminobutyric acid, fumaric acid, L-alaninamide, L-alanyl-glycine, L-proline, L-pyroglutamic acid and putrescine which were significantly inhibited by the chemical. So it could be useful to use streptomycin to control black shank when using it for bacterial wilt management. Although a few chemicals are available to control black shank in tobacco, our findings are valuable to assess the three agronomic chemical treatments to decrease the impacts of the disease.
ACKNOWLEDGMENT
This study was supported by National Natural Science Foundation of China (31360448), Guizhou Tobacco Company (201305, 201336, 201436) and Guizhou Science Technology Foundation ([2011]2337). We also thank the anonymous reviewers for critical reviews of the manuscript.
REFERENCES
- Brandt, U., H. Schagger and G. von Jagow, 1988. Characterisation of binding of the methoxyacrylate inhibitors to mitochondrial cytochrome c reductase. Eur. J. Biochem., 173: 499-506.
CrossRefDirect Link - Campanella, V., A. Ippolito and F. Nigro, 2002. of calcium salts in controlling Phytophthora root rot of citrus. Crop Prot., 21: 751-756.
CrossRefDirect Link - Cartwright, D.K. and H.W. Spurr Jr., 1998. Biological control of Phytophthora parasitica var. nicotianae on tobacco seedlings with non-pathogenic binucleate Rhizoctonia fungi. Soil Biol. Biochem., 30: 1879-1884.
CrossRefDirect Link - Chabane, K., P. Leroux and G. Bompeix, 1996. Resistance to fungicides and streptomycin in Phytophthora parasitica: Genetic determinism and use in hybrid determination. Phytopathol. Mediterr., 35: 82-90.
Direct Link - Donaldson, S.P. and J.W. Deacon, 1992. Role of calcium in adhesion and germination of zoospore cysts of Pythium: A model to explain infection of host plants. J. Gen. Microbiol., 138: 2051-2059.
CrossRefDirect Link - Druzhinina, I.S., M. Schmoll, B. Seiboth and C.P. Kubicek, 2006. Global carbon utilization profiles of wild-type, mutant and transformant strains of Hypocrea jecorina. Applied Envron. Microbiol., 72: 2126-2133.
CrossRefDirect Link - Eckert, J.W. and P.H. Tsao, 1962. A selective antibiotic medium for isolation of Phytophthora and Pythium from plant roots. Phytopathology, 52: 771-777.
Direct Link - Halsall, D.M. and R.I. Forrester, 1977. Effects of certain cations on the formation and infectivity of Phytophthora zoospores. 1. Effects of calcium, magnesium, potassium and iron ions. Can. J. Microbiol., 23: 994-1001.
CrossRefDirect Link - Heyman, F., B. Lindahl, L. Persson, M. Wikstrom and J. Stenlid, 2007. Calcium concentrations of soil affect suppressiveness against Aphanomyces root rot of pea. Soil Biol. Biochem., 39: 2222-2229.
CrossRefDirect Link - Hickmann, C.J., 1958. Phytophthora-plant destroyer. Trans. Br. Mycol. Soc., 41: 1-13.
CrossRefDirect Link - Hu, S.Q., 2009. Preliminary study on control effect of effervescent troche of agricultural streptomycin against tobacco bacterial wilt. Crop Res., 23: 27-29.
Direct Link - Jacobi, W.R., C.E. Main and N.T. Powell, 1983. Influence of temperature and rainfall on the development of tobacco black shank. Phytopathology, 73: 139-143.
Direct Link - Kong, P., G.W. Moorman, J.D. Lea-Cox, D.S. Ross, P.A. Richardson and C. Hong, 2009. Zoosporic tolerance to pH stress and its implications for Phytophthora species in aquatic ecosystems. Applied Environ. Microbiol., 75: 4307-4314.
CrossRefDirect Link - Kuhn, P.J., D. Pitt, S.A. Lee, G. Wakley and A.N. Sheppard, 1991. Effects of dimethomorph on the morphology and ultrastructure of Phytophthora. Mycol. Res., 95: 333-340.
CrossRefDirect Link - Lee, H.B., Y. Kim, J.C. Kim, G.J. Choi, S.H. Park, C.J. Kim and H.S. Jung, 2005. Activity of some aminoglycoside antibiotics against true fungi, Phytophthora and Pythium species. J.Applied Microbiol., 99: 836-843.
CrossRefDirect Link - Liu, A.Y., 2001. Inhibition of streptomycin on some oomycetes. Acta Phytophylacica Sinica, 28: 254-258.
Direct Link - Papaspyridi, L.M., P. Katapodis, Z. Gonou-Zagou, E. Kapsanaki-Gotsi and P. Christakopoulos, 2011. Growth and biomass production with enhanced β-glucan and dietary fibre contents of Ganoderma austral ATHUM 4345 in a batch-stirred tank bioreactor. Eng. Life Sci., 11: 65-74.
CrossRefDirect Link - Rice, A.V. and R.S. Currah, 2005. Profiles from Biolog FF plates and morphological characteristics support the recognition of Oidiodendron fimicola sp. nov. Stud. Mycol., 53: 75-82.
CrossRef - Rooke, D.M. and R.C. Shattock, 1983. Effect of chloramphenicol and streptomycin on developmental stages of Phytophthom infestans. J. Gen. Microbiol., 129: 3401-3410.
CrossRefDirect Link - Serrano, M.S., P. De Vita, P. Fernandez-Rebollo and M.E.S. Hernandez, 2012. Calcium fertilizers induce soil suppressiveness to Phytophthora cinnamomi root rot of Quercus ilex. Eur. J. Plant Pathol., 132: 271-279.
CrossRefDirect Link - Singh, M.P., 2009. Application of Biolog FF Microplate for substrate utilization and metabolite profiling of closely related fungi. J. Microbiol. Meth., 77: 102-108.
CrossRefDirect Link - Sugimoto, T., M. Aino, M. Sugimoto and K. Watanabe, 2005. Reduction of Phytophthora stem rot disease on soybeans by the application of CaCl2 and Ca(NO3)2. J. Phytopathol., 153: 536-543.
CrossRefDirect Link - Sundin, G.W., 2002. Distinct recent lineages of the strA-strB streptomycin-resistance genes in clinical and environmental bacteria. Curr. Microbiol., 45: 63-69.
CrossRefDirect Link - Sundin, G.W. and C.L. Bender, 1993. Ecological and genetic analysis of copper and streptomycin resistance in Pseudomonas syringae pv. syringae. Applied Environ. Microbiol., 59: 1018-1024.
Direct Link - Von Broembsen, S.L. and J.W. Deacon, 1996. Effects of calcium on germination and further zoospore release from zoospore cysts of Phytophthora parasitica. Mycol. Res., 100: 1498-1504.
CrossRefDirect Link - Von Broembsen, S.L. and J.W. Deacon, 1997. Calcium interference with zoospore biology and infectivity of Phytophthora parasitica in nutrient irrigation solutions. Phytopathology, 87: 522-528.
CrossRefDirect Link - Wang, H.C., X.J. Chen, L.T. Cai, Y. Cao and N. Lu et al., 2013. Race distribution and distribution of sensitivities to mefenoxam among isolates of Phytophthora parasitica var. nicotianae in Guizhou Province of China. Crop Prot., 52: 136-140.
CrossRefDirect Link - Wang, H.C., M.S. Wang, H.Q. Xia, S.J. Yang and Y.S. Guo et al., 2013. First report of Fusarium wilt of tobacco caused by Fusarium kyushuense in China. Plant Dis., 97: 424-424.
CrossRefDirect Link - Whittaker, S.L., S. Assinder and D.S. Shaw, 1996. Inheritance of streptomycin and chloramphenicol resistance in Phytophthora infestans: Evidence for co-segregation of mitochondrial DNA and streptomycin resistance. Mycol. Res., 100: 87-92.
CrossRefDirect Link - Yuan, S.K., X.L. Liu, B.G. Gu, J. Dong and H. Jiang, 2006. Sensitivity of Phytophthora infestans to flumorph: In vitro determination of baseline sensitivity and the risk of resistance. Plant Pathol., 55: 258-263.
CrossRefDirect Link - Sun, H.Y., H.C. Wang, G. Stammler, J.X. Ma and M.G. Zhou, 2010. Baseline sensitivity of populations of Phytophthora capsici from China to three Carboxylic Acid Amide (CAA) fungicides and sequence analysis of cholinephosphotranferases from a CAA-sensitive isolate and CAA-resistant laboratory mutants. J. Phytopathol., 158: 244-252.
CrossRefDirect Link