
Qi-Yao Yang
Key Laboratory of Pest Management in Crops, Ministry of Agriculture, Institute of Plant Protection,Chinese Academy of Agricultural Sciences, Beijing, 100081, People�s Republic of China
Ke Jia
Key Laboratory of Pest Management in Crops, Ministry of Agriculture, Institute of Plant Protection,Chinese Academy of Agricultural Sciences, Beijing, 100081, People�s Republic of China
Wen-Yi Geng
Key Laboratory of Pest Management in Crops, Ministry of Agriculture, Institute of Plant Protection,Chinese Academy of Agricultural Sciences, Beijing, 100081, People�s Republic of China
Rong-Jun Guo
Key Laboratory of Pest Management in Crops, Ministry of Agriculture, Institute of Plant Protection,Chinese Academy of Agricultural Sciences, Beijing, 100081, People�s Republic of China
Shi-Dong Li
Key Laboratory of Pest Management in Crops, Ministry of Agriculture, Institute of Plant Protection,Chinese Academy of Agricultural Sciences, Beijing, 100081, People�s Republic of China
Plant Pathology Journal
Year: 2014 | Volume: 13 | Issue: 3 | Page No.: 160-166







ABSTRACT
Bacillus subtilis B006 has been attempted in this study to control wilt disease of cucumber. In order to clear its suppressing mechanism in rhizosphere, population of Fusarium oxysporum f. sp. cucumerinum J. H. Owen (Foc) and B006 in cucumber rhizosphere were determined using real-time PCR. Real-time PCR program of B. subtilis B006 in soil was established for first time with the limit as low as 78.0 CFU g-1. Real-time PCR assays showed that combined use of B006 in substrate, drenching during transplantation and hole application with organic fertilizer can improve its colonization in rhizosphere by 10 times in comparison with its single use in substrate. And its presence resulted in the decline of Foc number over 10 times and 2 weeks delay of the disease development and reduction of disease severity in comparison with the non-B006 treatment. Greenhouse test showed that the addition of B006 powder in substrate can control cucumber wilt effectively with the control efficacy of 53.6%, combined use of 30 mL B006 suspension by drenching at the concentration of 106 CFU g-1 with hole application of 10 g organic fertilizer per seedling during transplantation resulted in the best control performance of 72.2%, while single use of organic fertilizer only resulted in the control performance of 29.4% in comparison with blank control. This study improves our understanding for the interaction of B006 and Foc in cucumber rhizosphere further and provides us a suitable application way of B. subtilis as a bioagent for cucumber wilt management.
PDF Abstract XML References Citation
Received: January 24, 2014;
Accepted: May 15, 2014;
Published: June 07, 2014
How to cite this article
Qi-Yao Yang, Ke Jia, Wen-Yi Geng, Rong-Jun Guo and Shi-Dong Li, 2014. Management of Cucumber Wilt Disease by Bacillus subtilis B006 Through
Suppression of Fusarium oxysporum f. sp. cucumerinum in Rhizosphere. Plant Pathology Journal, 13: 160-166.
DOI: 10.3923/ppj.2014.160.166
URL: https://scialert.net/abstract/?doi=ppj.2014.160.166
DOI: 10.3923/ppj.2014.160.166
URL: https://scialert.net/abstract/?doi=ppj.2014.160.166
INTRODUCTION
Vascular wilt of cucumber, caused by Fusarium oxysporum f. sp. cucumerinum J. H. Owen (Foc) often occurred in replanting field and leads to great loss (Owen, 1955; Jenkins and Wehner, 1983; Martinez et al., 2003). The disease is difficult to manage by chemical fungicide because F. oxysporum can grow along the xylem vessel and survive in soil as chlamydospores or saprophytes in plant debris over a year (Ogura, 1992). Grafting of cucumbers onto resistant rootstocks is an effective way to prevent the economic loss, however cucumber can lose the good taste because of the grafting. Soil fumigation using methyl bromide can provide satisfactory disease control in greenhouse but methyl bromide will be banned to use from 2015 (Ajwa et al., 2002). Therefore, biological control is becoming an alternative way to control the Fusarium wilt. Some antagonists showed potential to suppress this disease, such as mycorrhizal fungi and Trichoderma (Martinez-Medina et al., 2011; Chen et al., 2012), Penicillium (De Cal et al., 2009), Streptomyces (Zhao et al., 2012) and Bacillus (Chen et al., 2010). However, the control efficacies of antagonists are not always stable in fields due to the unsatisfied colonization in the rhizosphere to perform their function sustainably.
Antagonists are often used in several ways including seed coating (Bennett and Whipps, 2008), drenching or root dipping during transplantation (Copping, 2004; Xue et al., 2009), applying in nursery pot or mixing with organic fertilizer (Cao et al., 2011; Latha et al., 2011). Among them, seed coating and drenching are convenient ways to get maximum inhibition of pathogen. However, in field, the application way of antagonists must be considered to meet the needs of crop planting. In cucumber planting, seedlings are usually sown in nursery pots and then transplanted to fields. Therefore, applying antagonists either in nursery pots or during transplantation make a good choice. Cao et al. (2011) reported the combination of B. subtilis SQR9 with fertilizer in nursery pot and reapplication during transplantation could effectively reduce the incidence of cucumber fusarium wilt about 49-61% in pot tests. However, the interaction of strain SQR9 with Foc in rhizosphere was not known clear. In Cao’s study, the colonization of SQR9 was not examined in cucumber roots in field. Therefore, the suppression of Foc population might be caused by the antibiotics produced previously during the fermentation process of compost inoculated with Bacillus SQR9.
B. subtilis strain B006 showed great potential to suppress Foc in vitro (Yang et al., 2012). In this study, its ability to manage cucumber wilt disease under different application ways was examined and the populations of strain B006 and Foc in rhizosphere were detected in the same samples to try to clear the control mechanism.
MATERIALS AND METHODS
Bacterial and fungal cultures: Antagonistic strain B006 was pre-isolated and identified against Foc in vitro (Yang et al., 2012). The B006 and Foc cultures were maintained on nutrient agar slants and 1/4 PDA slants, respectively and kept at 4°C. Dry powder of B. subtilis B006 was formulated by fermentation in 500 L fermenting apparatus at 30°C for 2 days. The fermentation liquid was adsorbed by diatomite, dried at 50°C and crashed into 40 μm powder. The final B006 number was at 109 CFU g-1 powder.
Materials for planting: Cucumber cultivar Zhongnong 8 was procured from Seed Corporation of Chinese Academy of Agricultural Sciences (Beijing) with middle resistance to wilt disease. Nursery substrate was obtained from Langfang Seed and Seedling Corporation (Langfang City, Hebei province). Nutrient components of the substrate were 2-4% NPK, 30% firmuletes, 42% organic materials and pH 5.5-6.5. Organic fertilizer was procured from Quanlinjia Organic Fertilizer Corporation (Shandong Province), nutrient components ≥40%.
Specificity and sensitivity of real-time PCR for B. subtilis B006 in rhizosphere: After incubation of strain B006 in Nutrient Broth (NB) at 30°C for 24 h, cells were collected by centrifugation at 11500 g and washed twice with sterile distilled water. Total DNA was extracted following the instruction of TIANamp bacteria DNA kit (TIANGEN, China). Oligonucleotide primers SCAR-A 5'-ACCCCCGAAGCTGATTA-3' and SCAR-S 5'-TGGCCTTTTCTTAGTTGTC-3' (Geng et al., 2011) were employed for amplification of specific DNA of strain B006. PCR reactions were carried out with a thermocycler (Bio-Rad) under the following conditions: 10 min at 94°C, 35 cycles of 30 sec at 94°C, 45 sec at 56°C, 50 sec at 72°C and one final step of 10 min at 72°C. The PCR product was purified on 1.2% agarose gel using Gel Extraction Kit (TIANGEN, China), ligated into pMD18-T vector (Takara, Japan) and transformed into Escherichia coli DH5α competent cells. Positive transformants were screened on LB plates containing 100 μg mL-1 of ampicillin and then amplified with M13 primers and sequenced to confirm the correct clone of the target fragment.
Real-time PCR was performed using the Bio-Rad iQTM 5 Multicolor RT-PCR Detection System (Bio-Rad) with SYBR Premix Dimer Eraser (Takara) containing SYBR Green I. The PCR conditions was 1 min at 95°C, followed by 45 cycles of denaturation at 94°C for 15 sec, 30 sec at 55°C, 30 sec at 72°C and 2 min at 72°C and melt curve condition was 81 cycles from 55.0 to 95.0°C with an increasing of 0.5°C per 15 sec.
For soil detection, B006 powder was mixed with 0.60 mm naturally dried garden soil and ten fold serially diluted to give the final concentration of B006 in range from 7.80x107 to 78.0 CFU g-1. Soil without B006 powder was taken as control. DNA was extracted with UltraCleanTM DNA extraction Kit (MoBio, Carlsbad, USA) as described above. The final volume of each DNA extract was 50 mL. And 2 mL of 10 fold dilution were used for real-time PCR. Ct values were obtained from Bio-Rad iQTM5 and transformed into values of representing DNA copies using plasmid DNA amplification curve. Then standard curve for B006 detection in soil was constructed using logarithm-transformed values of DNA copies plotted against logarithm-transformed values of B006 concentration.
Real-time PCR for detection of Foc in rhizosphere: Genomic DNA of Foc was extracted from fresh hyphal powder ground with liquid nitrogen by Biospin Fungus Genomic DNA Extraction Kit (BSC14S1, BioFlux). Oligonucleotide primer pair FocF3:5'-AAACGAGCCCGCTATTTGAG-3' and Foc R7:5'-TATTTCCTCCACATTGCCATG-3' (Lievens et al., 2007) was employed for amplification of specific DNA of Foc. Conditions for PCR reactions were same to the method described by Lievens et al. (2007). Real-time PCR program was 1 min at 94°C, followed by 45 cycles of denaturation at 94°C for 15 sec, 30 sec at 62°C, 30 sec at 72°C and 2 min at 72°C and melt curve condition was 81 cycles from 55.0 to 95.0°C with an increasing of 0.5°C per 10 sec. Ct value was measured and standard curve was set up by Bio-Rad iQTM 5 software.
For soil detection, one gram of Foc hyphal powder was suspended in 9 mL sterile water, 10 fold serially diluted and mixed with dry garden soil in a range of 20 μg to 2 ng in 1 g of soil. Triplicates of each dilution were performed. Soil (0.5 g) without hypha suspension was taken as control. Procedure of standard curve construction for Foc detection in soil was same to the detection method for B006.
Greenhouse experiments: Experiment was carried out in the Foc inoculated field (18x6 m ) in the greenhouse (Langfang city, Hebei Province). Foc was cultured in Czapek liquid medium (NaNO3 3 g, K2H (PO4)3 1 g, MgSO4.7H2O 0.5 g, KCl 0.5 g, FeSO4 0.01 g, sugar 30 g, distilled water 1 L) at 26°C for 3 days with shaking at 180 rpm. Forty milliliter liquid cultures were inoculated into 300 mL millet seed/rice hulls (50: 50) in flasks (Burgess et al., 2008) and incubated at 26°C for about 2 weeks to allow the complete colonization of Foc mycelium on the substrate. One gram of solid medium was suspended in 9 mL sterile water and the spore density was determined by counting under microscopy. The solid cultures were inoculated into field soil with an estimation of the spore density at 105 spores per gram surface soil (20 cm soil) and the actual Foc population in each block was determined by real-time PCR before transplantation. The population of B006 in the field was also determined because B006 was ever used in last crop.
Dry B006 powder was mixed with nursery substrate at a final concentration of 106 CFU g-1 substrate and nursery substrate without B006 was taken as control. Cucumber seeds were soaked in distilled water over night at 28°C and plump seeds were selected and seeded in nursery pots, one seed in each pot. Seedlings at 3 or 4 leaf stage were transplanted into four field blocks with different treatments of B006 powder as follows: CK1, substrate with non-B006, direct transplantation of seedlings; CK2, substrate with non-B006, seedlings were transplanted to soil with addition of 10 g organic fertilizer per seedling; T1, single use of B006 by mixing with nursery substrate followed by direct transplantation; T2, addition of B006 in nursery substrate followed by drenching with B006 suspension at 106 CFU mL-1 (30 mL per seedling) during transplantation and 1 week after, respectively; T3, in addition of same treatments with T2, 10 g organic fertilizer/seedling was applied during transplantation. Fifteen seedlings in a row for each treatment were planted in every block.
Three to four cucumber roots and rhizosphere soil were randomly collected from each treatment at 0, 1, 3, 5 and 7 weeks after transplantation. Total DNA was extracted from soil as described above. Population of Bacillus B006 and Foc were determined by real-time PCR and analyzed by iQTM5 software. Disease index of cucumber wilt were investigated at the 3rd and 5th week after transplantation, respectively according to the method described by Zhao and Wu (2001).
Statistical analysis: The data was statistically analyzed by SPSS 17.0 software and significant differences were determined according to Duncan’s new multiple range test at p<0.05.
RESULTS
Specificity of real-time PCR of B. subtilis B006 and Foc: Genomic DNA of strain B006 and Foc were amplified using their specific primers SCAR-A/SCAR-R and FocF3/FocR7, respectively and specific DNA fragments of 523 and 244 bp were amplified. Intense fluorescent signal from genomic DNA extract of strain B006 and Foc were detected. Primer dimer could be detected in the same but the melt temperature of target DNA of strain B006 and primer dimer was at 86.0°C and 74.0°C, respectively. This difference showed that the amplification of strain B006 was not affected by the existence of primer dimer. The melt temperature of target DNA of Foc strain and primer dimer was at 82.0 and 76.0°C, respectively indicating the effective amplification for Foc.
Standard curve and sensitivity of real-time PCR assay of B. subtilis B006 and Foc: Perfect amplification curve was obtained by real-time PCR assay of B. subtilis B006 in soil and a standard curve for B. subtilis B006 detection in soil was constructed using logarithm-transformed values of DNA copies plotted against logarithm-transformed values of B006 concentration. The standard curve was linear over the B006 concentration in soil ranged from 7.80x107~78.0 CFU g-1 and the regress equation of linearity Y = 0.9726 X-1.0988 (r2 = 0.9981) was obtained.
The standard curve for Foc detection in soil was linear over the Foc concentration in soil ranged from 2 ng g-1 to 20 μg g-1 and the regress equation of linearity Y = 0.8446 X + 2.145 (r2 = 0.9959) was obtained.
Real-time PCR of B. subtilis B006 and Foc in cucumber rhizosphere: Detection of B006 and Foc by real-time PCR in the infected soil before transplantation showed that the population of Foc was averaged to be 5.47±0.27x105 copies g-1 soil and the number of B006 was at 4.37±1.85x103 CFU g-1 soil. B. subtilis B006 and Foc populations were determined, respectively in same samples by real-time PCR after seeds germinating and dynamics were shown in Fig. 1.
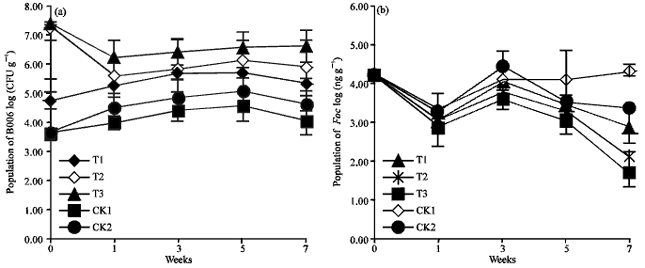 | |
| Fig. 1(a-b): | Population of B. subtilis, (a) B006 and (b) Foc in cucumber rhizosphere determined by real-time PCR at different growing time following three different application ways of B006. CK1, substrate with non-B006, direct transplantation of seedlings at 3 or 4 leaf stage; CK2 substrate with non-B006, seedlings were transplanted to soil with addition of 10 g organic fertilizer per seedling; T1, addition of B006 in nursery substrate followed by direct transplantation; T2, addition of B006 in nursery substrate followed by drenching with B006 suspension at 106 CFU mL-1 (30 mL per seedling) during transplantation and 1 week after, respectively; T3, in addition of same treatments with T2, 10 g organic fertilizer per seedling was applied during transplantation |
| Table 1: | Efficacy of different application ways of B. subtilis B006 in suppressing of cucumber fusarium wilt during different growth period |
 | |
| *Data in the same column followed by the different letters are significantly different (p<0.05); 1CK1, substrate with non-B006, direct transplantation of seedlings at 3 or 4 leaf-stage; CK2, substrate with non-B006, seedlings were transplanted to soil with addition of 10 g organic fertilizer per seedling; T1, addition of B006 in nursery substrate followed by direct transplantation; T2, addition of B006 in nursery substrate followed by drenching with B006 suspension at 106 CFU mL-1 (30 mL per seedling) during transplantation and 1 week after, respectively; T3, in addition of the same treated method with; T2, 10 g organic fertilizer per seedling was applied during transplantation, 2and3 was the control efficacy of each treatment, compared with CK1 and CK2, respectively | |
Although the initial populations of B006 were different among different treatments, the colonization dynamics of B006 in cucumber rhizosphere were similar except for declines in treatments T2 and T3 at the first week. Application of B006 powder strongly suppressed Foc growth and propagation in rhizosphere as demonstrated by a great reduction of Foc number by 10-100 times in comparison with non B006 treatment. Among three B006 treatments, T3 treatment with a re-application of B006 in combination with organic fertilizer resulted in the best suppressing performance. At the 7th week after transplantation, the Foc number decreased to 1.68 log (ng) per gram of soil, while it remained to be 4.33 log (ng) per gram and 3.37 log (ng) per gram of soil in treatments without B006 and single use of fertilizer, respectively. These results showed that application of B006 performed good suppressing of Foc in cucumber rhizosphere.
Efficacy of B. subtilis B006 powder in suppressing disease development: Investigation of disease index at the 3rd and the 5th week after transplantation revealed that application of B006 suppressed cucumber fusarium wilt effectively at the 5th week in comparison with non B006 treatment. The average control efficacy in T1 and T2 reached 53.6 and 58.6% (Table 1), respectively. Combined use of organic fertilizer with B006 in T3 treatment resulted in the best control efficacy of 72.2% and 58.5% (Table 1), compared with treatments without B006 and single use of organic fertilizer, respectively. While single use of organic fertilizer only led to the suppression of 29.4% (Table 1). These results indicated that amendment of B006 powder in nursery substrate was more effective than use of fertilizer on the control of cucumber fusarium wilt.
DISCUSSION
As a fungicide, antagonists are often used in seed treatment, drenching or dipping. Seed treatment provides unique opportunities for beneficial microbes to antagonize the pathogen and protects crop against soil-borne fungal pathogens (Harman et al., 1991; Mao et al., 1997; Ayoubi et al., 2012). However, single seed treatment with antagonists is not always effective (Georgakopoulos et al., 2002), this might attribute to the rapid infection by pathogen in a very short time as 2 h in a diseased soil (Nelson, 1987; Osburn et al., 1989). This phenomenon was also described in other research (Savchuk and Fernando, 2004) as indicated by the obvious variation of suppressing in ascospores germination of S. sclerotiorum by application time of Pseudomonas spp. (DF-41 and PA-23). In China, farmers usually plant commercial cucumber seeds coating with chemical fungicide or buy commercial seedlings from Seedlings Corporation. This situation makes seed treatment with beneficial microbes inconvenient. In order to effectively prevent seed and root invasions by the pathogen, addition of the antagonists into nursery substrate was used as an alternative way to help antagonists colonize in cucumber rhizosphere prior to pathogen infection. Cao et al. (2011) improved the application methods of antagonistic Bacillus spp. by mixing of Bacillus with manure followed by fermentation to form a bio-fertilizer and used the bio-fertilizer in substrate followed by furrow application. This combined use of bio-fertilizer effectively controlled cucumber Fusarium wilt. However the biocontrol efficacy usually varied with the different components of soil amendments and pathogenic fungi (Bonanomi et al., 2010). In this study, we compared the control efficacy of three B006 powder application ways. Investigation of the disease index showed that addition of B006 powder in nursery substrate played more important role than the other application ways of B006 poder, although the combined use of in nursery substrate with drenching resulted in the best control of fusarium wilt.
The success of biological control might be predicted with the understanding of the dynamics between a pathogen and a biocontrol agent (BCA) in rhizosphere and clearly know how the BCA and pathogen influence each other's population and life cycles and how the biological and physical environment influence this relationship (Paulitz, 2000). Many researches reported the colonization of Bacillus on plant seedlings using gfp marker in laboratory conditions (Cao et al., 2011; Fan et al., 2011) or recovery of the gfp marked cells from field (Hao et al., 2010). However, due to the difficult transformation of our wild B. subtilis strain B006, an alternative way must be used for the examination of its population dynamics in field. Real-time PCR has been proved to be an effective method to detect the changing of benefits or pathogens in rhizosphere. Based on our previous screening of specific markers to detect strain B006 in soil or in soil added with fertilizer (Geng et al., 2011), we developed a real-time PCR method to detect strain B006 in field soil. Using this method, we can inspect the interaction of strain B006 and Foc on the base of studying the population density fluxes over time. Real-time PCR analysis indicated that nursery substrate and organic fertilizer could improve B. subtilis B006 multiplication and reduced the Foc number for one order of magnitude or more. This suggests the importance of colonization of B006 in seedlings rhizosphere and persistence at a high population during plants growing season in disease suppressing. However, colonization of antagonists or pathogens in rhizosphere was affected by many physical characters, including root exudates (Yaryura et al., 2008), soil water pressure (Walker et al., 2002; Hussain et al., 2005) and soil type (Zheng and Sinclair, 2000) etc. Ecological research about the interaction of antagonistic Bacillus and Foc with plants will be carried out in our further work.
It is concluded that B. subtilis B006 is an effective strain which grows quickly and produces metabolites including surfactin and fengycin in vitro and in rhizosphere (Yang et al., 2012). Many researchers have demonstrated surfactin and fengycin involved in the suppressing of pathogen infection (Asaka and Shoda, 1996; Bais et al., 2004; Liu et al., 2011; Yi et al., 2011), little is known about the function of surfactin and fengycin produced by strain B006 in rhizosphere, further work is on progress in our study.
ACKNOWLEDGMENTS
This study was supported by the Program of China Agriculture Research System (CARS-25-06B) and "five-year" plan of national science and technology projects for the rural development in China (2012BAD19B0103).
REFERENCES
- Asaka, O. and M. Shoda, 1996. Biocontrol of Rhizoctonia solani damping-off of tomato with Bacillus subtilis RB14. Applied Environ. Microbiol., 62: 4081-4085.
Direct Link - Ayoubi, N., D. Zafari and M. Mirabolfathy, 2012. Combination of Trichoderma species and Bradyrhizobium japonicum in control of Phytophthora sojae and soybean growth. J. Crop Prot., 1: 67-79.
Direct Link - Bais, H.P., R. Fall and J.M. Vivanco, 2004. Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol., 134: 307-319.
Direct Link - Bennett, A.J. and J.M. Whipps, 2008. Beneficial microorganism survival on seed, roots and in rhizosphere soil following application to seed during drum priming. Biol. Control, 44: 349-361.
CrossRefDirect Link - Bonanomi, G., V. Antignani, M. Capodilupo and F. Scala, 2010. Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem., 42: 136-144.
CrossRefDirect Link - Cao, Y., Z.H. Zhang, N. Ling, Y.J. Yuan, X.Y. Zheng, B. Shen and Q.R. Shen, 2011. Bacillus subtilis SQR 9 can control Fusarium wilt in cucumber by colonizing plant roots. Biol. Fertil. Soils, 47: 495-506.
CrossRefDirect Link - Chen, F., M. Wang, Y. Zheng, J. Luo, X. Yang and X. Wang, 2010. Quantitative changes of plant defense enzymes and phytohormone in biocontrol of cucumber Fusarium wilt by Bacillus subtilis B579. World J. Microbiol. Biotechnol., 26: 675-684.
CrossRefDirect Link - Chen, L.H., X.Q. Huang, F.G. Zhang, D.K. Zhao, X.M. Yang and Q.R. Shen, 2012. Application of Trichoderma harzianum SQR-T037 bio-organic fertiliser significantly controls Fusarium wilt and affects the microbial communities of continuously cropped soil of cucumber. J. Sci. Food Agric., 92: 2465-2470.
CrossRef - Fan, B., X.H. Chen, A. Budiharjo, W. Bleiss, J. Vater and R. Borriss, 2011. Efficient colonization of plant roots by the plant growth promoting bacterium Bacillus amyloliquefaciens FZB42, engineered to express green fluorescent protein. J. Biotechnol., 151: 303-311.
CrossRefPubMedDirect Link - Georgakopoulos, D.G., P. Fiddaman, C. Leifert and N.E. Malathrakis, 2002. Biological control of cucumber and sugar beet damping-off caused by Pythium ultimum with bacterial and fungal antagonists. J. Applied Microbiol., 92: 1078-1086.
Direct Link - Hao, B.Q., L.P. Ma and X.W. Qiao, 2010. Colonization ability of plant growth promoting Bacillus B96-II-gfp labeled with GFP. Chin. J. Eco-Agric., 8: 861-865.
Direct Link - Harman, G.E., X. Jin, T.E. stasz, G. Peruzzotti, A.C. Leopold and A.G. Taylor, 1991. Production of conidial biomass of Trichoderma harzianum for biological control. Biol. Control, 1: 23-38.
CrossRef - Hussain, S., M. Powelson, N. Christensen and S.J.A. Shah, 2005. Soil water pressure affects population dynamics of biocontrol agents of Verticillium dahliae, the cause of potato early dying. Plant Pathol. J., 4: 113-121.
CrossRefDirect Link - Jenkins Jr., S.F. and T.C. Wehner, 1983. Occurrence of Fusarium oxysporum f. sp. cucumerinum on greenhouse-grown Cucumis sativus seed stocks in North Carolina. Plant Dis., 67: 1024-1025.
CrossRefDirect Link - Latha, P., T. Anand, V. Prakasam, E.I. Jonathan, M. Paramathma and R. Samiyappan, 2011. Combining Pseudomonas, Bacillus and Trichoderma strains with organic amendments and micronutrient to enhance suppression of collar and root rot disease in physic nut. Applied Soil Ecol., 49: 215-223.
CrossRefDirect Link - Lievens, B., L. Claes, D.J. Vakalounakis, A.C. Vanachter and B.P. Thomma, 2007. A robust identification and detection assay to discriminate the cucumber pathogens Fusarium oxysporum f. sp. cucumerinum and f. sp. radicis-cucumerinum. Environ. Microbiol., 9: 2145-2161.
CrossRefPubMedDirect Link - Mao, W., J.A. Lewis, P.K. Hebbar and R.D. Lumsden, 1997. Seed treatment with a fungal or a bacterial antagonist for reducing corn damping-off caused by species of Pythium and Fusarium. Plant Dis., 81: 450-454.
CrossRefDirect Link - Martinez, R., M.I. Aguilar, M.L. Guirado, A. Alvarez and J. Gomez, 2003. First report of fusarium wilt of cucumber caused by Fusarium oxysporum in Spain. Plant Pathol., 6: 410-410.
CrossRefDirect Link - Martinez-Medina, A., A. Roldan and J.A. Pascual, 2011. Interaction between arbuscular mycorrhizal fungi and Trichoderma harzianum under conventional and low input fertilization field condition in melon crops: Growth response and Fusarium wilt biocontrol. Applied Soil Ecol., 47: 98-105.
CrossRefDirect Link - Nelson, E.B., 1987. Rapid germination of sporangia of Pythium species in response to volatiles from germinating seeds. Phytopathology, 77: 1108-1112.
CrossRefDirect Link - Osburn, R.M., M.N. Schroth, J.G. Hancock and M. Hendson, 1989. Dynamics of sugar beet seed colonization by Pythium ultimum and Pseudomonas species: Effects on seed rot and damping-off. Phytopathology, 79: 709-716.
Direct Link - Paulitz, T.C., 2000. Population dynamics of biocontrol agents and pathogens in soils and rhizospheres. Eur. J. Plant Pathol., 106: 401-413.
CrossRefDirect Link - Savchuk, S. and W.G.D. Fernando, 2004. Effect of timing of application and population dynamics on the degree of biological control of Sclerotinia sclerotiorum by bacterial antagonists. FEMS Microbiol. Ecol., 49: 379-388.
CrossRefPubMedDirect Link - Walker, R., S. Rossall and M.J.C. Asher, 2002. Colonization of the developing rhizosphere of sugar beet seedlings by potential biocontrol agents applied as seed treatments. J. Applied Microbiol., 92: 228-237.
CrossRefPubMedDirect Link - Xue, Q.Y., Y. Chen, S.M. Li, L.F. Chen, G.C. Ding, D.W. Guo and J.H. Guo, 2009. Evaluation of the strains of Acinetobacter and Enterobacter as potential biocontrol agents against Ralstonia wilt of tomato. Biol. Control, 48: 252-258.
CrossRef - Yang, Q., Y. Suo, R. Guo, S. Li and X. Xu, 2012. Antifungal activities and principal component analysis of Bacillus subtilis B006 against Fusarium oxysporum f. sp. cucumerinum and Phytophthora capsici. Chin. J. Biol. Control, 8: 235-242.
Direct Link - Yaryura, P.M., M. Leon, O.S. Correa, N.L. Kerber, N.L. Pucheu and A.F. Garcia, 2008. Assessment of the role of chemotaxis and biofilm formation as requirements for colonization of roots and seeds of soybean plants by Bacillus amyloliquefaciens BNM339. Curr. Microbiol., 56: 625-632.
CrossRefPubMedDirect Link - Yi, H.S., J.W. Yang, S.Y. Ghim and C.M. Ryu, 2011. A cry for help from leaf to root: Above ground insect feeding leads to the recruitment of rhizosphere microbes for plant self-protection against subsequent diverse attacks. Plant Signal. Behav., 6: 1192-1194.
PubMed - Zhao, S., C.M. Du and C.Y. Tian, 2012. Suppression of Fusarium oxysporum and induced resistance of plants involved in the biocontrol of Cucumber Fusarium Wilt by Streptomyces bikiniensis HD-087. World J. Microbiol. Biotechnol., 28: 2919-2927.
CrossRefPubMedDirect Link - Zhao, X. and D. Wu, 2001. Studies on techniques of in vitro evaluation of fusaric wilt-resistance in cucumber. J. South Chin. Agric. Univ., 22: 39-41, 45.
Direct Link - Zheng, X.Y. and J.B. Sinclair, 2000. The effects of traits of Bacillus megaterium onseed and root colonization and their correlation withthe suppression of Rhizoctonia root rot of soybean. BioControl, 45: 223-243.
CrossRefDirect Link