
Emre Yoruk
Programme of Molecular Biology and Genetics, Institute of Science, Istanbul University, Vezneciler, Istanbul, Turkey
Gulruh Albayrak
Department of Molecular Biology and Genetics, Faculty of Science, Vezneciler, Istanbul University, Istanbul, Turkey
Plant Pathology Journal
Year: 2014 | Volume: 13 | Issue: 2 | Page No.: 133-138







ABSTRACT
The tri5 gene cluster is associated with production of trichothecene B type mycotoxins in Fusarium graminearum and F. culmorum. Multifunctional oxygenase and trichodiene synthase encoded by tri4 and tri5 genes, respectively, belonging to this cluster are responsible for production of trichothecenes. In this study, tri4 and tri5 gene expression levels were determined in 14 F. graminearum and 24 F. culmorum isolates causing head blight in wheat, maize and barley collected from different regions of Turkey. Isolates producing oxygenase and trichodiene synthase in high levels were distinguished from low-producers using two-step reverse-transcription comparative quantitative real-time PCR. Slope values were -3.63, -3.4 and -3.4 for amplification of tri4, tri5 and β-tubulin genes, respectively. Cp values were ranged from 17.08±0.0 to 38.82±1.66. The highest tri4 and tri5 expression levels were determined in 4F isolate of F. graminearum. ΔΔCT ratios were calculated as 0.1371 for tri4 gene and 0.865 for tri5 gene. However, maximum tri4 and tri5 expressions were detected in 20F (ΔΔCT = 0.2196) and 9F (ΔΔCT = 0.3761) isolates in F. culmorum, respectively. Quantification of definite toxin content can contribute to determination of pathogen infection grade and to gaining the information about fungal biomass on field crops and their products.
PDF Abstract XML References Citation
Received: December 16, 2013;
Accepted: March 31, 2014;
Published: April 26, 2014
How to cite this article
Emre Yoruk and Gulruh Albayrak, 2014. Tri4 and tri5 Gene Expression Analysis in Fusarium graminearum
and
F. culmorum Isolates by qPCR. Plant Pathology Journal, 13: 133-138.
DOI: 10.3923/ppj.2014.133.138
URL: https://scialert.net/abstract/?doi=ppj.2014.133.138
DOI: 10.3923/ppj.2014.133.138
URL: https://scialert.net/abstract/?doi=ppj.2014.133.138
INTRODUCTION
Mycotoxins are secondary metabolites produced by fungal organisms. Fusarium species produce several exotoxin types such as zearalenone, fumonisins, butenolide, trichothecenes etc. (Desjardins and Proctor, 2007). Class B trichothecenes include deoxynivalenol (DON), nivalenol (NIV) and their acetylated derivatives. Cereals which were infected by Fusarium sp. are also contaminated with them. Moreover, their acute toxicity is harmful to health of human and animal consuming the contaminated foods (Goswami and Kistler, 2004; Foroud and Eudes, 2009; Asran and Amal, 2011).
Currently, techniques based on amplification of eleven genes located in tri5 gene cluster are effectively used for chemotyping of Fusarium sp. producing trichothecene B (Chandler et al., 2003; Haratian et al., 2008; Yoruk and Albayrak, 2012). The eleven genes are separately responsible for catalysis of each step of the trichothecene B biosynthesis. Among them, tri5 and tri4 genes code trichodiene synthase and multifunctional oxygenase responsible for catalysing the first and second steps of biosynthetic pathway, respectively. Therefore, both of main genes are required for synthesis of trichothecenes. Determination of their expression levels is also essential for discrimination and identification of isolates (Doohan et al., 1999; Marin et al., 2010). Moreover, findings obtained from related studies contribute to understanding the importance of these genes in biochemical pathway. In addition, microbiological determination approaches such as HPLC (high performance liquid chromatography) or MS (mass spectrometry) are used to determine mycotoxins. Although results obtained from these approaches are reliable, cost of them is extremely higher than amplification based ones. Moreover, their throughputs are limited. Commercial ELISA (enzyme linked immunosorbent assay) based test systems are also used to determine toxin contents despite their low accuracy. Also, preparation of samples used in it is still time consuming (Ward et al., 2004; Gherbawy and Voigt, 2010).
Toxin biosynthetic key genes in tri5 cluster are used in amplification by polymerase chain reaction (PCR) for the quantification of trichothecene B. To determine their gene expression levels is more reliable, inexpensive, laborious and precise than microbiological determination approaches. Besides, it is possible to detect the mycotoxins even if pathogen produces only a little amount of them.
| Table 1: | Geographic origin, hosts and gene expression values of F. graminearum and F. culmorum isolates used in this study |
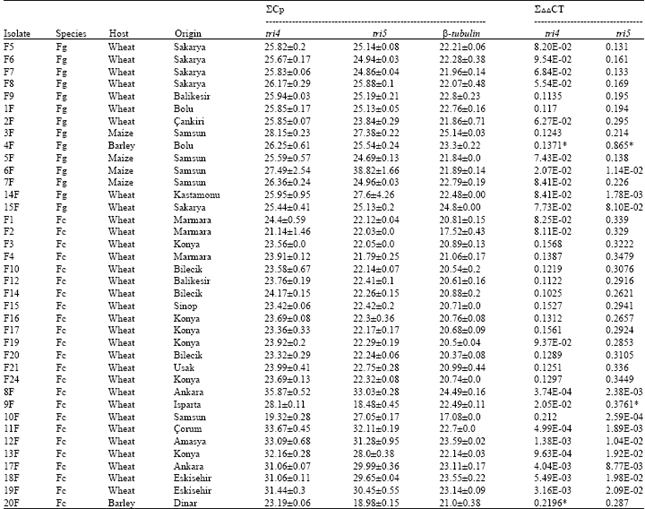 | |
| *Maximum gene expression, Fg: Fusarium graminearum, Fc: Fusarium culmorum | |
| Table 2: | Primer and prob sets used in this study |
 | |
| *Primers designed by Hue et al. (1999) | |
Therefore, quantification of trichothecene B type mycotoxins is a basic and powerful approach for getting knowledge about activity of genes located in that cluster (Kimura et al., 2003; Alexander et al., 2011).
In this study, tri4 and tri5 gene expression analysis were carried out in 14 F. graminearum and 24 F. culmorum isolates causing head blight in wheat, maize and barley collected from different regions of Turkey by using a two-step reverse-transcription comparative real-time PCR.
MATERIALS AND METHODS
14 F. graminearum and 24 F. culmorum monosporic isolates (Leslie et al., 2006) were kindly provided from Dr. Berna Tunali, Department of Plant Protection, Agricultural Faculty, Samsun On Dokuz Mayis University. Collection region, hosts and gene expression values are listed in Table 1.
Fungal isolates were grown in potato dextrose agar (PDA) plates at 25°C for 7 days. Total RNA extracted from 7-day-old mycelium by using TriPure isolation reagent s(Roche, Switzerland) including phenol and guanidine thiocyanate. Quantity of total RNA was assessed by Nanodrop 2000C.
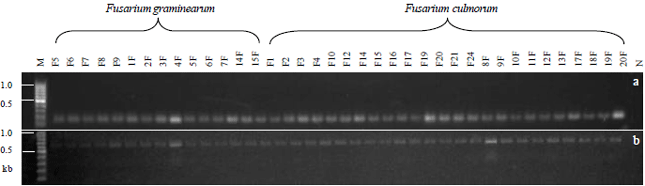 | |
| Fig. 1(a-b): | Tri4 gene amplicons by (a) tri4 primers and (b) tri5 gene bands using Tox5 primers in all isolates by RT-PCR. M: 100 bp DNA ladder (Thermo), N: Negative control (no template) |
Complementary DNA (cDNA) synthesis was carried out in a reaction volume of 25 μL containing: 1μg total RNA, 60 μM random hexamer, 1X reaction buffer, 5 μM DTT, 1U protector RNase inhibitor, 1 mM dNTPs and 1U transcriptor HF reverse transcriptase. cDNA synthesis was performed in thermal cycler (Bio-Rad, France) at 65°C for 10 min, 55°C for 30 min and 85°C for 5 min.
For gene expression analysis, primers and probe sets (Table 2) were designed using primer3 (Rozen and Skaletsky, 2000) and universal probe library online services (www.rocheappliedscience.com) to amplify tri4, tri5 and β-tubulin genes. qPCR assays were performed in Light Cycler 480 II system (Roche, Switzerland) using Taq-Man probes and conducted in a reaction volume of 20 μL containing: cDNA amount corresponding to 1μg total RNA, 1X probe master mix, 2pmol probe and 4pmol of each primers. β-tubulin gene was used as house keeping gene. Cycling conditions consisted of pre-denaturation at 95°C for 10 min; followed by 45 cycles at 95°C for 10 s, at 58°C for 30 s, at 72°C for 1 s. For each experiment set, standard series with a range of three dilutions were prepared in the determination of slope values. Crossing point values (Cp) were determined for three genes in all isolates. The tri4 and tri5 gene expression levels were normalized according to the β-tubulin gene and target/reference ratio results. Each experiment sets were replicated at least three times.
For confirmation of quantitative gene expression results, three times replicated RT-PCR assays were performed using cDNA molecules of 38 isolates to amplify each of three genes. PCR was carried out in a volume of 25 μL comprised of cDNA amount corresponding to 1 μg total RNA, 1X PCR buffer, 2.5 mM MgCl2, 0.25 mM of dNTPs, 10 pmol of each primer and 1U of Taq DNA polymerase (Promega, USA). Tox5 primer set designed by Hue et al. (1999) and Tri4 and FusTb1 primers designed (by us for the first time) in this study were used in the amplification of tri5, tri4 and β-tubulin genes, respectively (Table 2). PCR conditions were performed at 94°C for 5 min; 35 cycles at 94°C for 45 s, at 58°C for 45 s, at 72°C for 45 s and at 72°C for 5 min.
Furthermore, amplification of tri5, tri4 and β-tubulin genes in the genomic DNA’s was performed using Tox5, Tri4 and FusTb1 primers, respectively, in a volume of total 25 μL. The procedure was followed RT-PCR described previously, except for using genomic DNA as template (50 ng). RT-PCR and PCR amplification products were separated by electrophoresis in 1.5% agarose gels and visualized using gel visualization system (Avegene, X-lite 200).
RESULTS
Total RNA molecules with high quality (A260/A280 = 1.9-2) and quantity (1.5-2 μg μL-1) were isolated from 38 fungal samples. Expressions of tri4 and tri5 genes together with housekeeping gene (β-tubulin) in all isolates were detected by RT-PCR and qPCR techniques.
A 139 bp tri4 amplicon (Fig. 1a) and 658 bp fragment (Fig. 1b) belonging to tri5 gene were yielded in all isolates by RT-PCR analysis. The most strong amplification products were obtained from 4F and 20 F isolates for tri4 and also from 4F and 9F for tri 5 genes (Fig. 1a and 1b). In addition, 465 bp long common bands of β-tubulin were observed in 38 isolates with a similar banding pattern (data not shown).
In qPCR analysis, Cp and slope values and target/reference ratios were calculated according to signal peaks (Table 1). Slope values were -3.63, -3.4 and -3.4 for amplification of tri4, tri5 and β-tubulin genes, respectively. That the slope values reach to -3.3 indicated expression levels of three genes as valuable. ΣCp values for tri4 were calculated ranged from 19.32±0.28 to 35.87±0.52. ΣCp values for tri5 gene were found as 18.48±0.45-38.82±1.66 whereas values for β-tubulin were between 17.08±0.0 and 25.14±0.03. As shown in Table 1, the highest tri4 and tri5 expression levels were obtained from 4F isolate of F. graminearum. ΣΔΔCT ratios were calculated 0.1371 for tri4 and 0.865 for tri5 gene amplifications. However, maximum tri4 and tri5 levels were detected in 20F (ΣΔΔCT=0.2196) and 9F (ΣΔΔCT=0.3761) isolates of F. culmorum, respectively. It is clear that RT-PCR and qPCR findings verify each other.
Presence of the tri4, tri5 and β-tubulin genes were confirmed with standard PCR assay. Tri4 primer pair amplified 139 bp amplicons in all isolates (data not shown). Tox5 primer pairs produced 658 bp DNA fragment while FusTb1 yielded 465 bp amplicons (data not shown). Thus, the expression profile obtained in this study is reliable.
DISCUSSION
There is not any definite struggle strategy to be used in the Fusarium infections of field crops. Therefore, it seems to be difficult to overcome the Fusarium problems in agriculture in the near future. So, detection of mycotoxins via chemotyping has become an important strategy to avoid them in the food chain (Wilbert and Kemmelmeier, 2003; Gherbawy and Voigt, 2010). Activation of eleven genes shows differences from one strain to another even when they located in tri5 cluster. Expression of some tri genes is definitely required for production of mycotoxins. However, a number of strains contain them as pseudogene or completely deleted ones (Chandler et al., 2003; Kim et al., 2003; Kimura et al., 2003; Alexander et al., 2011). In that situation, mycotoxin can still continue to be produced. Scherm et al. (2011) determined an increase of trichothecene production in silenced tri6 gene samples when they are compared to control groups. The results of this study indicate that some genes found in that cluster are not essential in the production of mycotoxins. For trichothecene B production, determining the gene expression and expression levels of each gene is very important.
Fungal microorganisms producing high levels of definite mycotoxins can be discriminated from non-producers and also low-producers. For this purpose, quantification of target genes encoding those mycotoxins and their expression levels can now be carried out by qPCR assay. For the first time, Schnerr et al. (2001) applied this assay for quantification of trichodiene synthase in 30 wheat samples. They showed PCR based method was as useful as microbiological determination approaches.
In Turkey, genotyping of F. graminearum and F. culmorum isolates was done by different research groups (Arici and Koc, 2010; Gurel et al., 2010; Yoruk et al., 2011, 2012). Chemotypes for trichothecene B in both of two species, which are associated with especially head blight, were studied with HPLC and PCR analysis (Tunali et al., 2006; Yoruk and Albayrak, 2012; Mert-Turk and Gencer, 2013).
In this study, expression of tri4 and tri5 genes, encoding oxygenase and trichodiene synthase respectively, were analyzed from 38 local Fusarium isolates using two-step transcription comparative quantitative real-time PCR. The results indicated that different amount of mycotoxins were produced by different isolates, although they may contain the same gene(s). Also, isolates have maximum gene expression profiles were distinguished from low-producers. In some countries, proposed mycotoxin limits are determined in food and feed products. Our results could be useful for that purpose if there were legislative limitations for trichothecenes in Turkey. Quantification of definite toxin content can be also useful in the evaluation of infection grade. Mycotoxin quantification can inform us about fungal biomass and their products on field crops. Moreover, results obtained from similar studies can be used in routine monitoring programs associated with disease controlling.
Expression analysis of genes responsible for class B trichothecene production in phytopathogenic Fusarium species was carried out in this study. Trichothecene producer isolates were easily distinguished from low-producers by qPCR assays in a short time. This kind of studies allow pre-selection of isolates which produce high level of trichothecenes in order to use further investigation such as gene disruption, gene silencing etc. Although all isolates possess the tri4 and tri5 genes, significant differences in gene expression profiles were shown. Isolate selection by distinguishing producers and also high-producers from low- and non-producers is so important in plant pathology. Therefore, selection of trichothecene producers and determination of gene expression levels are efficient strategies especially in quelling. This paper report will useful for wide range of researchers in plant pathology.
ACKNOWLEDGEMENT
This study was supported by Research Fund of the Istanbul University, project number: T23489/11052012. The authors are grateful to Prof. Dr. Berna Tunali from Department of Plant Protection, Agricultural Faculty, Samsun On Dokuz Mayis University for providing fungal material.
REFERENCES
- Alexander, N.J., S.P. McCormick, C. Waalwijk, T.V.D. Lee and R.H. Proctor, 2011. The genetic basis for 3-ADON and 15-ADON trichothecene chemotypes in Fusarium. Fungal Genet. Biol., 48: 485-495.
CrossRefPubMedDirect Link - Arici, S.E. and N.K. Koc, 2010. RAPD-PCR analysis of genetic variation among isolates of Fusarium graminearum and Fusarium culmorum from wheat in Adana Turkey. Pak. J. Biol. Sci., 13: 138-142.
CrossRefDirect Link - Asran, M.R. and M.I.E. Amal, 2011. Aggressiveness of certain Fusarium graminearum isolates on wheat seedlings and relation with their trichothecene production. Plant Pathol. J., 10: 36-41.
CrossRefDirect Link - Chandler, E.A., D.R. Simpson, M.A. Thomsett and P. Nicholson, 2003. Development of PCR assays to Tri7 and Tri13 trichothecene biosynthetic genes and characterisation of chemotypes of Fusarium graminearum, Fusarium culmorum and Fusarium cerealis. Physiol. Mol. Plant Pathol., 62: 355-367.
CrossRefDirect Link - Desjardins, A.E. and R.H. Proctor, 2007. Molecular biology of Fusarium mycotoxins. Int. J. Food Microbiol., 119: 47-50.
CrossRef - Doohan, F.M., G. Weston, H.N. Rezanoor, D.W. Parry and P. Nicholson, 1999. Development and use of a reverse transcription-PCR assay to study expression of tri5 by Fusarium species in vitro and in planta. Applied Environ. Microbiol., 65: 3850-3854.
PubMed - Foroud, N.A. and F. Eudes, 2009. Trichothecenes in cereal grains. Int. J. Mol. Sci., 10: 147-173.
Direct Link - Goswami, R.S. and H.C. Kistler, 2004. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol., 5: 515-525.
CrossRefDirect Link - Gurel, F., G. Albayrak, O. Diken, E. Cepni and B. Tunali, 2010. Use of Rep-PCR for genetic diversity analyses in Fusarium culmorum. J. Phytopathol., 158: 387-389.
CrossRefDirect Link - Haratian, M., B. Sharifnabi, A. Alizadeh and N. Safaie, 2008. PCR analysis of the Tri13 gene to determine the genetic potential of Fusarium graminearum isolates from Iran to produce nivalenol and deoxynivalenol. Mycopathologia, 166: 109-116.
CrossRef - Hue, F.X., M. Huerre, M.A. Rouffault and C. de Bievre, 1999. Specific detection of Fusarium species in blood and tissues by a PCR technique. J. Clin. Microbiol., 37: 2434-2438.
Direct Link - Kim, H.S., T. Lee, M. Dawlatana, S.H. Yun and Y.W. Lee, 2003. Polymorphism of trichothecene biosynthesis genes in deoxynivalenol-and nivalenol-producing Fusarium graminearum isolates. Mycol. Res., 107: 190-197.
CrossRefPubMedDirect Link - Kimura, M., T. Tokai, K. O'Donnell, T.J. Ward and M. Fujimura et al., 2003. The trichothecene biosynthesis gene cluster of Fusarium graminearum F15 contains a limited number of essential pathway genes and expressed non-essential genes. FEBS Lett., 539: 105-110.
CrossRefPubMedDirect Link - Marin, P., M. Jurado, N. Magan, C. Vazquez and M.T. Gonzalez-Jaen, 2010. Effect of solute stress and temperature on growth rate and tri5 gene expression using real time RT-PCR in Fusarium graminearum from Spanish wheat. Int. J. Food Microb., 140: 169-174.
CrossRefPubMedDirect Link - Mert-Turk, F. and R. Gencer, 2013. Distribution of the 3-AcDON, 15-AcDON and NIV chemotypes of Fusarium culmorum in the North-West of Turkey. Plant Protect. Sci., 49: 57-64.
Direct Link - Rozen, S. and H. Skaletsky, 2000. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol., 132: 365-386.
CrossRefDirect Link - Scherm, B., M. Orru, V. Balmas, F. Spanu and E. Azara et al., 2011. Altered trichothecene biosynthesis in TRI6-silenced transformants of Fusarium culmorum influences the severity of crown and foot rot on durum wheat seedlings. Mol. Plant Pathol., 12: 759-771.
CrossRefPubMedDirect Link - Schnerr, H., L. Niessen and R.F. Vogel, 2001. Real time detection of the tri5 gene in Fusarium species by LightCyclerTM-PCR using SYBR®Green I for continuous fluorescence monitoring. Int. J. Food Microbiol., 71: 53-61.
CrossRefPubMedDirect Link - Tunali, B., J. Nicol, F.Y. Erol and G. Altiparmak, 2006. Pathogenicity of Turkish crown and head scab isolates on stem bases on winter wheat under greenhouse conditions. Plant Pathol. J., 5: 143-149.
Direct Link - Ward, E., S.J. Foster, B.A. Fraaije and H.A. Mccartney, 2004. Plant pathogen diagnostics: Immunological and nucleic acid-based approaches. Ann. Applied Biol., 145: 1-16.
CrossRefDirect Link - Wilbert, F.M. and C. Kemmelmeier, 2003. Identification of deoxynivalenol, 3-acetyldeoxynivalenol and zearalenone in galactose oxidase-producing isolates of Fusarium graminearum. J. Basic Microbiol., 43: 148-157.
CrossRefPubMedDirect Link - Yoruk, E. and G. Albayrak, 2012. Chemotyping of Fusarium graminearum and F. culmorum isolates from Turkey by PCR assay. Mycopathologia, 173: 53-61.
CrossRefPubMedDirect Link