
Hui Zhang
Key Laboratory at Universities of Xinjiang, Uygur Autonomous Region for Oasis,Agricultural Pest Management and Plant Protection Resource Utilization,Shihezi University, Shihezi, 832003, China
Li Chen
Key Laboratory at Universities of Xinjiang, Uygur Autonomous Region for Oasis,Agricultural Pest Management and Plant Protection Resource Utilization,Shihezi University, Shihezi, 832003, China
SiFeng Zhao
Key Laboratory at Universities of Xinjiang, Uygur Autonomous Region for Oasis,Agricultural Pest Management and Plant Protection Resource Utilization,Shihezi University, Shihezi, 832003, China
Juan Ren
Key Laboratory at Universities of Xinjiang, Uygur Autonomous Region for Oasis,Agricultural Pest Management and Plant Protection Resource Utilization,Shihezi University, Shihezi, 832003, China
XiaoLei Cao
Key Laboratory at Universities of Xinjiang, Uygur Autonomous Region for Oasis,Agricultural Pest Management and Plant Protection Resource Utilization,Shihezi University, Shihezi, 832003, China
Plant Pathology Journal
Year: 2014 | Volume: 13 | Issue: 2 | Page No.: 125-132







ABSTRACT
Bacillus subtilis S44 is a promising plant growth-promoting rhizobacterium and a potent biocontrol agent isolated from cotton rhizosphere. The ituD gene was cloned and identified from S44. A suicide vector containing ituD gene from B. subtilis S44 was recombined into the B. subtilis S44 chromosome to disrupt the ituD gene. Recombination was demonstrated by PCR, antifungal activity and HPLC confirmed the loss of the ituD gene. The growth rate of the recombinant is severely depressed compared with wild type and tetracycline had no effect on the growth rate the mutant strains. Antifungal activity in cell-free extracts of the recombinant has almost no antifungal activity compared with the wild strain. HPLC analysis show that the wild strian produce iturin A at an amount of 89.4 μg mL-1 while the ituD mutant only 3.41 μg mL-1. In pot and plot tests, the control effects of S44 treatment were 80.30 and 72.97%, respectively. The mutant strains S1 and S2 exhibit no control effect of cotton Rhizoctoniosis. These results suggest that the ituD gene was important for iturin A synthesis and biocontrol activity of S44.
PDF Abstract XML References Citation
Received: November 04, 2013;
Accepted: March 09, 2014;
Published: April 26, 2014
How to cite this article
Hui Zhang, Li Chen, SiFeng Zhao, Juan Ren and XiaoLei Cao, 2014. Knockout of ituD Gene of Bacillus subtilis S44 Strain and
Impact of its Biocontrol Effect to Cotton Rhizoctoniosis. Plant Pathology Journal, 13: 125-132.
DOI: 10.3923/ppj.2014.125.132
URL: https://scialert.net/abstract/?doi=ppj.2014.125.132
DOI: 10.3923/ppj.2014.125.132
URL: https://scialert.net/abstract/?doi=ppj.2014.125.132
INTRODUCTION
The use of microorganisms for biological purposes has become an effective alternative for the control of plant pathogens. Many examples of formulations that use bacteria or fungal strains possess significant potential for biocontrol applications. Among these strains, members of the genus Bacillus are well-known antibiotic producers (Stein, 2005). These strains possess an advantage over other biocontrol microorganisms for their inherent capability to form endospores and for their resistance to extreme conditions. Bacillus subtilis strains produce a broad spectrum of bioactive peptides with significant potential for biotechnological and biopharmaceutical applications. A known class of such compounds includes the lipopeptides surfactin, fengycin and iturin compounds. These compounds are amphiphilic membrane active-peptide antibiotics with potent antimicrobial activities that can be used as biopesticides for plant protection (Asaka and Shoda, 1996; Hiraoka et al., 1992; Hsieh et al., 2008). Antibiotics in the family iturin which are represented by iturin A, mycosubtilin and bacillomycin, are most commonly studied for their biocontrol activity. These antibiotics are heptapeptides with a β-amino fatty acid that exhibits strong antifungal activity (Kim et al., 2010; Crane et al., 2013; Pathak and Keharia, 2013). Iturin A is a cyclic lipopeptide antibiotic linked to a β-amino fatty acid chain with a length of 14-17 carbons (Ongena and Jacques, 2008). Seven amino acid residues of its structure are linked in sequence as follows: L-Asn-D-Tyr-D-Asn-L-Gln-L-Pro-D-Asn-L-Ser. The-COOH of 7-Serine and the -NH2 of the β-amino fatty acid chain are condensed to a cyclic structure. By relying on its membrane permeable properties (Bonmatin et al., 2003), the special structure endows this chain with strong antifungal action (Mizumoto et al., 2007; Arrebola et al., 2010; Kim et al., 2010; Yao et al., 2012; Ye et al., 2012). In Bacillus, the iturin A operon is 38- 40 kb in size and consists of four open reading frames: ituD, ituA, ituB and ituC. ituD encodes a putative malonyl coenzyme A transacylase, the disruption of which results in a specific deficiency or incapability to produce iturin A (Tsuge et al., 2001; Romero et al., 2007).
The bacterium B. subtilis S44 was described the isolation and partial characterization in previous study (Luo et al., 2010). This strain shows strong in vitro antifungal activity against various fungal phytopathogens and is capable of significantly decreasing the incidence of cotton Rhizoctoniosis, cotton Verticillium wilt and tomato root rot disease in the field (Luo et al., 2010, 2011). The strain S44 antifungi activity was determined, its antifungi ingredient is iturin A. This study primarily aims to reveal the function of iturin A in the biological control of soil-borne pathogens using B. subtilis S44 strain as well as to verify further the contribution of the ituD gene to the iturin A synthesis function. The results could be used for the commercial exploitation of the S44 strain for biofungicide production.
MATERIALS AND METHODS
Strains, plasmids and growth conditions: The bacterial culture S44 selected for this study was isolated from the rhizoplane of a field-grown cotton plant in Xinjiang in China and is identified as B. subtilis (unpublished data). Bacterial cultures were stored frozen in 10% glycerol at -70°C. Working cultures were maintained on tryptone yeast extract (TY; 5.0 g L-1 tryptone, 3.0 g L-1 yeast extract and 0.66 g L-1 CaCl2) agar slants at 4°C. The phytopathogenic fungal cultures were obtained from cottonroot in Xinjiang in China, maintained at 4°C and left to multiply (25±1°C) on Potato Dextrose Agar (PDA) medium. The S44 strain was grown in Nutrient Agar (NA) medium (3 g L-1 beef extract, 10 g L-1 peptone and 5 g L-1 NaCl) at 30°C with shaking (220 rpm). Luria Broth (LB) agar plates were prepared by adding 2% agar. Escherichia coli DH5α cells with plasmids were cultured aerobically at 37°C in LB medium (5 g L-1 yeast extract, 10 g L-1 tryptone, 5 g L-1 NaCl and 15 g L-1 agar) or on LB agar plates (1.5% agar).
Cloning of ituD gene: The ituD gene was amplified using the reported primer (Hsieh et al., 2008) of ituD-f: CGCGGATCCATGAACAATCTTGCCTTTTTA and ituD-r:ACATCGTACGTTATTTTAA AATCCGCAATT. B. subtilis S44 genomes were used as templates. Polymerase Chain Reaction (PCR) amplification was conducted in a 50 μL reaction mixture containing 2 μL of DNA, 10 μM dNTP [TaKaRa Biotechnology Co. Ltd., Japan], 10 μM primers and one unit ExTaq DNA polymerase [TaKaRa Biotechnology Co. Ltd., Japan]. Samples were denatured for 5 min at 94°C and then run for 31 cycles of 45 sec each at 94°C, 60 sec at 55°C and 90 sec at 72°C, with a final extension of 10 min at 72°C. The PCR products were separated through 1% agarose gel electrophoresis. The 1.2 kb band was excised and the gel was purified. The PCR product was purified using a Universal DNA purification kit [Tiangen Biotech (Beijing) Co. Ltd., China] and was ligated into the PMD18-T vector [TaKaRa Biotechnology Co. Ltd., Japan]. The ligation product was transformed into E. coli DH5α. After the cultures were grown, the constructs were purified and sequenced. Homology studies were conducted using the NCBI Blast program (Zhang et al., 2000).
Construction of gene ituD disrupted strain: A 1.2 kb fragment of ituD was inserted into plasmid pUC18 [TaKaRa Biotechnology Co. Ltd., Japan] and digested with BamHI-Sph I [TaKaRa Biotechnology Co. Ltd., Japan] to yield pUC18-ituD. The Tcr gene cassette in plasmid pBR322 [TaKaRa Biotechnology Co. Ltd., Japan] was amplified as fragment using two primers of T-up: CGCATCGATTAGTTCTCAT GTTTGACAGCTTATCTT CGAT and T-down: TTAATCGATTCAGGTCGAGGT GGCCCGGCT (Zhang and Ge, 2008). Amplified fragment was inserted into the ClaI-digested pUC18-ituD plasmid to create the ituD-disruption vector pUC18-ituD::tet. The vector was transformed into E. coli DH5α and the plasmid preparations were obtained with the TIANprep mini plasmid kit [Tiangen Biotech (Beijing) Co. Ltd., China]. B. subtilis S44 was transformed according to the electroporation protocol (Cao et al., 2011), selecting LB agar plates containing tetracycline resistance at 5 μg mL-1. Integration into the targeted gene on the chromosome of B. subtilis S44 was confirmed by PCR and sequencing used primer ituD-f and ituD-r and the strains were called S1 and S2.
Antifungal activity assays with B. subtilis supernatants: The anti-fungal activity of the ituD knockout mutants (S1 and S2) of B. subtilis S44 was compared with that of wild-type cell on PDA agar plates using the dual culture technique (Yoshida et al., 2001). Bacterial inocula were prepared from cultures grown in Erlenmeyer flasks (250 mL) containing 100 mL of NA supplemented for 24 h at 37°C. Cell-free filtrates from different strain cultures were obtained through centrifugation at 13523 g for 15 min, followed by filtration through a 0.22 μm Polyethersulfone (PES) membrane disposable syringe filter unit. The Rhizoctonia solani cultures were grown on PDA for three days at 25°C. A 5 mm diameter mycelial section of R. solani composed of only a few mycelial sections was placed at the center of the dual-culture PDA plates and 100 μL cell-free filtrates from the different strain cultures were patched equidistantly along the perimeters of the plates at a distance of three cm from the fungus. Plates were incubated for three days at 25°C and the inhibition of fungal growth was assessed by the presence or absence of an inhibition zone and by comparison with NA controls.
Isolation and HPLC determination of iturin A: Strains were grown in NA medium for 48 h at 30°C. Iturin A was isolated according to the reported method (Yao et al., 2012) with some modifications. After cultivation, 1 L of the cultures was centrifuged at 13523Hg for 20 min and cell-free supernatant was precipitated by adding 6 mol L-1 HCl to a final pH of 2.0 and stored at 4°C overnight. The precipitate was collected by centrifugation at 13523Hg for 15 min at 4°C and then freeze-dried. The residue was extracted with 200 mL methanol under shaking for 24 h. The supernatant was obtained by centrifugation at 13523Hg for 15 min and then filtered through a 0.22 μm PES membrane disposable syringe filter unit. The filtrate was freeze-dried. The crude product was then dissolved in 50 mL deionized water, applied to a SupelcleanTM LC18 solid phase extraction cartridge (500 mg, 6 mL) and then eluted with 80% aqueous methanol. The collected elution was freeze-dried. The samples and iturin A I1774 [Sigma-Aldrich Co. Ltd., America] standards were dissolved in methanol [Merck Co. Ltd., Germany].
The iturin A infiltrate was quantified using reverse-phase high-performance liquid chromatography (HPLC) column (Lichrospher C18, 250 9 4.6 mm, Agilent Technologies Corporate, USA) on a Waters 2695 HPLC system operated at a flow rate of 0.9 mL-1 min. A mixture of acetonitrile and 10 mM ammonium acetate (35:65, v/v) was used as the eluent and the elution was monitored at 210 nm. The four peaks corresponding to the major iturin A homologues were regarded as the total iturin A production (Asaka and Shoda, 1996).
Biocontrol assays with cotton in greenhouse: The biocontrol activity of S44, S1 and S2 against cotton Rhizoctoniosis caused by R. solani was tested in greenhouse experiments. The soil used in this study was collected from a cotton field at the Shihezi University experiment station. The soil was sieved and autoclaved for 3 h at 160°C before use. R. solani and the plates were incubated at 28°C for 72 h. The whole pan was covered and set aside. A mycelium plug (10.0 cm2) R. solani culture was sliced into approximately 1 mm sections and incorporated into 700 g sterilized field soil in each 1000 mL pot. S44, S1 and S2 cell suspension preserved at -80°C were inoculated to 20 mL of NA medium in an Erlenmeyer flask (nominal volume, 50 mL) and cultivated at 37°C at 220 strokes per minute (spm) for 24 h. This preculture was then inoculated in 100 mL of NA media mentioned above at 3% inoculation in the flask was incubated at 30°C and 220 rpm, On three days, each culture was serially diluted in 0.85% NaCl solution and plated to L agar plates. The plates were incubated at 37°C for 16 h, the numbers of colonies were counted and Log Colony-forming Units (CFU) per milliliter of culture were expressed as the cell number. And then the strains were diluted to 1H108 CFU mL-1. Cotton seeds (Gossypium hirsutum L.) were surface-disinfected for 1 min with 70% ethanol, rinsed five times with sterile distilled water and then surface disinfected again for 5 min with 0.5% sodium hypochlorite. After at least 10 rinses with sterile distilled water, the seeds were soaked in 1H108 CFU mL-1 concentrations of S44, S1 and S2 for 4 h, after which they were taken out to dry. Twenty soaked cotton seeds were sown per pot and grown in a greenhouse under a regime with light/dark cycle of 16/8 hours at 30°C in daytime and 24°C in night for 10 days. Eight treatments were used in each experiment: S44 only, S44+ R. solani, S1 only, S1+ R. solani, S2 only, S2+ R. solani, water only and water+ R. solani. Each experiment with five replicates (pots) was performed twice. After 10 days, the shoot length and dry weight were measured. The percentage of diseased seedlings, disease index and control effect were determined. Disease severity was classified in five grades, (0) Stem and roots without discoloration, (1) Stem and roots locally discolored, (2) One-third to one-half of the stem and roots discolored, (3) One-half to two-third of stem and roots discolored and (4) seedling death:
 |
Biocontrol assays with cotton in plot: Biocontrol of cotton Rhizoctoniosis by bacterial strains S44, S1 and S2 were evaluated under field conditions. The field trial was conducted in the spring of 2013 at the Shihezi University experiment station, Xinjiang, China The block was situated on well-drained clay loam soil where cotton plants have been grown continuously for four years. The following treatments were included in the experiment: (1) Water soaking (control), (2) S44 soaking, (3) S1 soaking and (4) S2 soaking. The experimental design was a randomized complete block with four replications. Individual plots were 4H4 m. Cotton seeds were surface disinfected for 1 min with 70% ethanol, rinsed five times with sterile distilled water and then surface disinfected again for 5 min with 0.5% sodium hypochlorite. After at least 10 rinses with sterile distilled water, the seeds were soaked in 1H108 CFU mL-1 concentrations of S44, S1 and S2 for 4 h, after which they were taken out to dry. Two hundred seeds were sown in different plots. After 20 days, the shoot length and dry weight were measured. The percentage of diseased seedlings, disease index and control effect were determined.
Statistical analysis: The data obtained were subjected to ANOVA using SPSS 8.0 software for Windows (SPSS Inc.). The mean values were compared using the least significant difference test at p<0.05.
RESULTS
Sequence analysis of the ituD gene: The ituD gene contained 1,203 bp was deposited in GenBank under accession number KF381340 and encoded a 400-amino acid polypeptide. Similar to other ituD genes, ituD exhibits high G+C average content (42%). An alignment of the deduced amino acid sequence of ituD in the GenBank database show this ituD with the reported ituD amino acid sequence homology of 85%.
Disruption of gene ituD: The two colonies were selected randomly after the double crossover gene knockout process and the isolated genomic DNAs of the two colonies were confirmed by PCR and sequencing. PCR and sequencing appeared with the expected sizes of 2.5 kb (ituD knock-out mutant) and 1.2 kb (wild type strain) (Fig. 1). Thus, the colonies with Tcr had integrated the tetracycline resistance cassette into the ituD gene which was thus disrupted. Two strains were called B. subtilis S1 (ituDG) and B. subtilis S2 (ituDG) which would be used for further validation.
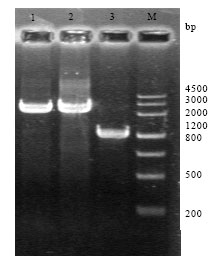 | |
| Fig. 1: | Result of 1% agarose gel electrophoresis of ituD gene PCR product of S44 and mutant strain. Lanes 1-4 contain, respectively. PCR-amplified products of ituD:tet and ituD gene: Lanes 1-2, ituD:tet gene, lanes 3, ituD gene |
Anti-fungal activity in ituD knockout mutant of B. subtilis The antifungal activity of S1 and S2 was compared with that of B. subtilis S44 wild-type on PDA agar plates, as shown in Fig. 2. Compared with the antifungal activity in the case of B. subtilis S44 wild-type (Fig. 2a), S1 and S2 (Fig. 2b, c) showed no anti-fungal activity. This result indicates that ituD disruption resulted in iturin A deficiency.
Growth rates of S44, S1 and S2 B. subtilis: To determine whether inhibition of antifungal activity had any effect on growth of B. subtilis S1 and S2, cells were grown in NA medium containing. Growth was measured as a change in OD600. The results (Fig. 3a) show that growth of the S1, S2, was significantly less than that of the wild type. To eliminate the possibility that the presence of tetracycline may be affecting growth, S1 and S2 were grown in varying concentrations of tetracycline from 0 to 10 μg mL-1. The results (Fig. 3b, c) show that tetracycline had no effect on the growth rate of S1 and S2.
Isolation and HPLC determination of iturin A: The crude extracts of the wild-type strain and mutants were analyzed using HPLC (Fig. 4). The wild type exhibited one major peak and several minor peaks at 210 nm (Fig. 4a). Results indicated that the wild type showed a retention time of 8.147, 11.432, 12.593 and 13.590 min (Fig. 4a) and could produce iturin A at an amount of 89.4 μg mL-1. HPLC analysis monitored at 210 nm confirmed that the produced iturin A was only 3.41 μg mL-1 in the ituD mutant (Fig. 4b).
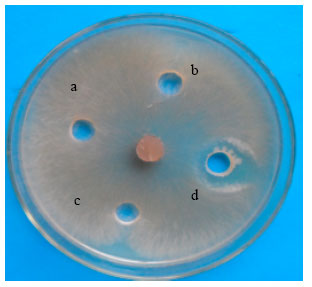 | |
| Fig. 2: | Effect of different strains of extraction of lipopeptides against R. solani, (a) CK, (b-c) Lipopeptide crude extract of mutant strain and (d) Lipopeptide crude extract of wild-type |
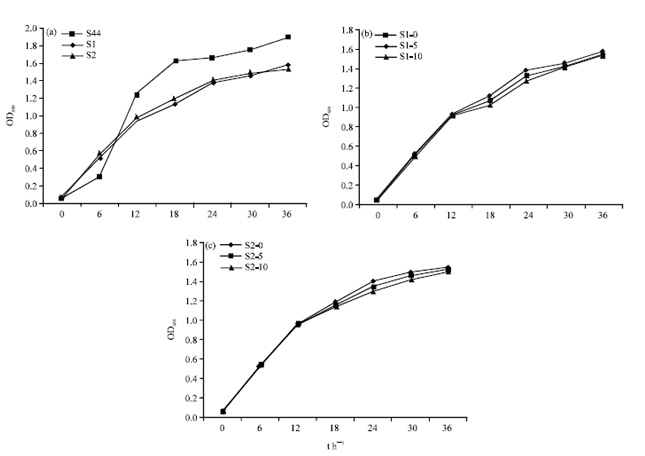 | |
| Fig. 3(a-c): | Growth rates of S44, S1 and S2 B. subtilis, (a) S4, S1, S2 grown in NA media, (b) S1 0, 5 and 10: 0, 5 and 10 μg mL-1 tetracycline and (c) S2 0, 5 and 10: 0, 5 and 10 μg mL-1 tetracycline, respectively |
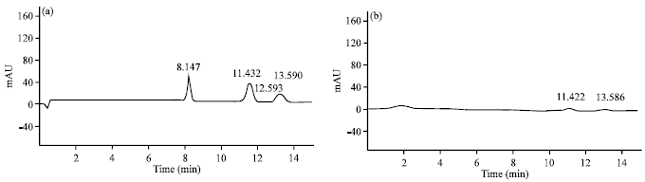 | |
| Fig. 4(a-b): | HPLC analysis of iturin A in B. subtilis strains, elution profiles at 210 nm for the (a) Wild type strain and (b) ituD knockout mutant |
Biocontrol assays with cotton in greenhouse: Seed treatment with S44, S1 and S2 can affect cotton seedling growth (Table 1). For S44, the shoot length decreased by 29.7%, whereas fresh and dry weight improved by 19.4 and 15.0%, respectively, compared with CK (water soaked cotton seeds). For S1, the shoot length decreased by 9.2%, whereas fresh and dry weight improved by 4.1 and 7.4%, respectively, compared with CK. For S2, the shoot length decreased by 0.8%, whereas fresh and dry weight improved by 4.7 and 9.2%, respectively, compared with CK. For S44, the shoot length decreased by 29.0 and 41.0% compared with S1 and S2, respectively.
| Table 1: | Effects of different treatments on cotton seedlings in pot tests |
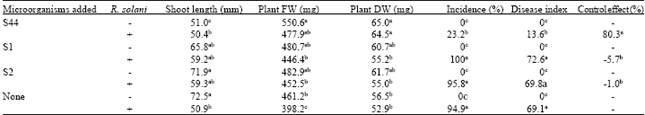 | |
| For each treatment, each data point is a mean of the results obtained from three replications, different letters represent significant difference (p<0.05) according to Duncan=s multiple range test, FW: Fresh weight, DW: Dry weight | |
| Table 2: | Control effects of different treatments on cotton seedlings in plot tests |
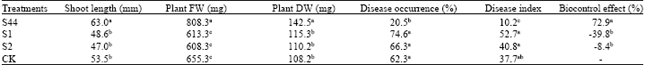 | |
| For each treatment, each data is a mean of the results obtained from three replications, different letters represent significant difference (p<0.05) according to Duncan=s multiple range rest, FW: Fresh weight, DW: Dry weight | |
The fresh and dry weight of S44 improved by 12.7, 12.3, 6.6 and 5.1% compared with S1and S2, respectively. These results which were obtained from the greenhouse tests, indicated that S44 inhibited cotton seeding growth but promoted plant accumulation of organic matter.
The control effects of S44, S1 and S2 exhibited significant differences (Table 1). The incidences of S44, S1 and S2 treatment were 23.2, 100.0 and 95.8%, respectively and the corresponding disease indexes were 13.6, 72.6 and 69.8. The control effect of S44 treatment was 80.3%. The mutant strains S1 and S2 have no control effect to cotton Rhizoctoniosis.
Biocontrol assays with cotton in plots: Seed treatment with S44, S1 and S2 can affect cotton seedling growth in field (Table 2). S44 treatment had a positive effect on the shoot as well as on fresh and dry weights. The S44 shoot length, fresh weight and dry weight were 63 mm, 808.3 mg and 142.5 mg, respectively. For S44, the shoot length, fresh weight and dry weight improved by 17.8, 23.3 and 31.7%, respectively, compared with CK; 22.9, 24.1 and 19.1% compared with S1 and 25.4, 24.7 and 22.7% compared with S2. The control effect of S44 treatment on cotton Rhizoctoniosis was 72.9%. The incidence and disease index of mutant strains with S1 and S2 treatment had no significant difference compared with CK.
DISCUSSION
Strains of B. subtilis have been reported as powerful biological control agents of plant diseases because of their capability to produce a wide variety of antimicrobial compounds, including lipopeptide antibiotics (Shoda, 2000; Stein, 2005; Ongena and Jacques, 2008). The majority of studies have focused primarily on the biocontrol potential of this bacterium to reduce diseases. However, the mechanism of action of these strains, especially the effect of lipopeptides on the target pathogens, has not been extensively investigated (Janisiewicz and Korsten, 2002). This study reports the biological potential of a B. subtilis strain to control cotton Rhizoctoniosis based on the production of iturin A.
To clarify the role of bacterial antibiotics in the control of plant diseases of a given biocontrol agent, molecular analysis by mutagenesis has been successfully used (Silo-Suh et al., 1994; Heungens and Parke, 2001). Inactivation of antibiotic production by mutagenesis disrupts biosynthetic genes, generally resulting in a reduction or loss of antagonistic capability to control the pathogen (Raaijmakers et al., 2002). Based on these premises and on the antifungal activity previously demonstrated by B. subtilis S44, focusing on the disruption of iturin A production. A site-directed mutagenesis strategy to inactivate the ituD gene of strain S44 was employed. The use of the electroporation protocol developed by Cao et al. (2011) to transform the B. subtilis component to construct S44 mutants that are deficient for iturin A production. Furthermore, antifungal activity, HPLC analysis, as well as pot and plot experiments provided clear evidence that this procedure is suitable for successful gene manipulation and molecular genetic analysis of this bacterium.
The potential role of the ituD gene was examined in growth of B. subtilis. The results indicated that the growth rate of the recombinant is severely depressed compared with wild type (Fig. 3a). This suggests that the ituD gene plays an important role in growth. The strains of S1 and S2 had the same growth rate in different concentration of tetracycline in the culture medium (Fig. 3b, c). The results show that tetracycline had no effect on the growth rate the mutant strains.
HPLC analysis showed iturin A still exist in the mutant strain, only 3.41 μg mL-1, may be due to mechanical residual, or lpa-14 gene regulation (Hsieh et al., 2008). Although a small amount of iturin A was detected in the mutant strain in this study, this strain did not show any antifungal activity (Fig. 2, 4). This fact highlights the importance of iturin A in the antagonistic activity of B. subtilis S44 against pathogens. Moreover, the results obtained from the pot and plot experiments using treated seeds confirmed the expected role of iturin A in the disease suppression capability of S44 wild type strain by demonstrating the biocontrol efficacy of wild type treatments compared with the inefficacy of the treatments with S1 or S2 (Table 1, 2). Taken together, the findings enable us to conclude that iturin A is a major factor involved in the biological control capability of the B. subtilis strain S44 against cotton Rhizoctoniosis elicited by R. solani.
In the pot experiment, the percentage of diseased plants treated with S1 and S2 (100 and 95.8%) was significantly higher than that of plants with S44 treatment using R. solani (23.2%). S44 treatment compared with the S1 and S2 treatments shoot length were no significant differences with R. solani. But treated with S44 has significant difference with S1 and S2 without R. solani (Table 1). Possibly owing to the higher concentration of bacteria, plant growth was suppressed. In contrast to the outcome of the pot experiment, the shoot length of S44 treatment was significantly higher than that of S1 and S2 in the plot experiment (Table 2). The effect was likely attributed to rain erosion and irrigation which reduces the amount of bacteria to promote plant growth. From these experiments, concluded that the major mechanism of suppression by S44 is production of the antibiotics iturin A.
The use of B. subtilis strains to control cotton seeding rot disease is a good alternative to the restricted use of chemical fungicides. In this context, this study provides experimental evidence of the strong antifungal effect and biological control capability of the B. subtilis strain S44 in the reduction of cotton seeding rot elicited by R. solani which is mostly based on iturin A antibiotic production. The results support the possibility of their use as a biological control agent of soil-borne diseases as well as the development of commercial formulations. However, lipopeptide production is not the only factor to consider in the development of this biological control product. Additional research on this strain is needed before it can be incorporated into the biological control program for cotton Rhizoctoniosis.
ACKNOWLEDGMENTS
This study was supported by the National Natural Science Foundation of China (30800733) and Xinjiang Corps Funds (No. 2013 CC002).
REFERENCES
- Arrebola, E., R. Jacobs and L. Korsten, 2010. Iturin A is the principal inhibitor in the biocontrol activity of Bacillus amyloliquefaciens PPCB004 against postharvest fungal pathogens. J. Applied Microbiol., 108: 386-395.
CrossRef - Asaka, O. and M. Shoda, 1996. Biocontrol of Rhizoctonia solani damping-off of tomato with Bacillus subtilis RB14. Applied Environ. Microbiol., 62: 4081-4085.
Direct Link - Bonmatin, J.M., O. Laprevote and F. Peypoux, 2003. Diversity among microbial cyclic lipopeptides: Iturins and surfactins. Activity-structure relationships to design new bioactive agents. Comb. Chem. High Throughput Screen., 6: 541-556.
CrossRef - Cao, G., X. Zhang, L. Zhong and Z. Lu, 2011. A modified electro-transformation method for Bacillus subtilis and its application in the production of antimicrobial lipopeptides. Biotechnol. Lett., 33: 1047-1051.
CrossRef - Crane, J.M., D.M. Gibson, R.H. Vaughan and G.C. Bergstrom, 2013. Iturin levels on wheat spikes linked to biological control of Fusarium head blight by Bacillus amyloliquefaciens. Phytopathology, 103: 146-155.
CrossRef - Heungens, K. and J.L. Parke, 2001. Postinfection biological control of oomycete pathogens of pea by Burkholderia cepacia AMMDR1. Phytopathology, 91: 383-391.
CrossRef - Hiraoka, H., T. Ano and M. Shoda, 1992. Characterization of Bacillus subtilis RB14, coproducer of peptide antibiotic iturin A and surfactin. J. Gen. Applied Microbiol., 38: 635-640.
CrossRef - Hsieh, F.C., T.C. Lin, M. Meng and S.S. Kao, 2008. Comparing methods for identifying Bacillus strains capable of producing the antifungal lipopeptide iturin A. Curr. Microbiol., 56: 1-5.
CrossRefDirect Link - Janisiewicz, J.W. and L. Korsten, 2002. Biological control of postharvest diseases of fruits. Annu. Rev. Phytopathol., 40: 411-441.
CrossRefDirect Link - Kim, P.I., J. Ryu, Y.H. Kim and Y.T. Chi, 2010. Production of biosurfactant lipopeptides Iturin A, fengycin and surfactin A from Bacillus subtilis CMB32 for control of Colletotrichum gloeosporioides. J. Microbiol. Biotechnol., 20: 138-145.
PubMedDirect Link - Luo, Y.N., J. Du, J.H. Li, Q.J. Xu and S.F. Zhao, 2010. Screening and identification of antagonistic Bacteria control Rhizoctonia solani-onduced damping-off of cotton. J. Shihezi Univ., 28: 555-560 (In Chines).
Direct Link - Mizumoto, S., M. Hirai and M. Shoda, 2007. Enhanced iturin A production by Bacillus subtilis and its effect on suppression of the plant pathogen Rhizoctonia solani. Applied Microbiol. Biotechnol., 75: 1267-1274.
CrossRef - Ongena, M. and P. Jacques, 2008. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol., 16: 115-125.
CrossRefDirect Link - Pathak, K.V. and H. Keharia, 2013. Characterization of fungal antagonistic bacilli isolated from aerial roots of banyan (Ficus benghalensis) using intact-cell MALDI-TOF mass spectrometry (ICMS). J. Applied Microbiol., 114: 1300-1310.
CrossRefDirect Link - Raaijmakers, J.M., M. Vlami and J.T. de Souza, 2002. Antibiotic production by bacterial biocontrol agents. Antonie Leeuwenhoek, 81: 537-547.
CrossRefDirect Link - Romero, D., A. de Vicente, R.H. Rakotoaly, S.E. Dufour and J.W. Veening et al., 2007. The iturin and fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Mol. Plant-Microbe Interact., 20: 430-440.
CrossRefDirect Link - Silo-Suh, L.A., B.J. Lethbridge, S.J. Raffel, H. He, J. Clardy and J. Handelsman, 1994. Biological activities of two fungistatic antibiotics produced by Bacillus cereus UW85. Applied Environ. Microbiol., 60: 2023-2030.
Direct Link - Stein, T., 2005. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol., 56: 845-857.
CrossRef - Tsuge, K., T. Akiyama and M. Shoda, 2001. Cloning, sequencing and characterization of the iturin A operon. J. Bacteriol., 183: 6265-6273.
CrossRef - Yao, D., Z. Ji, C. Wang, G. Qi, L. Zhang, X. Ma and S. Chen, 2012. Co-producing iturin A and poly-γ-glutamic acid from rapeseed meal under solid state fermentation by the newly isolated Bacillus subtilis strain 3-10. World J. Microbiol. Biotechnol., 28: 985-991.
CrossRefDirect Link - Ye, Y.F., Q.Q. Li, G. Fu, G.Q. Yuan, J.H. Miao and W. Lin, 2012. Identification of antifungal substance (Iturin A2) produced by Bacillus subtilis B47 and its effect on southern corn leaf blight. J. Integr. Agric., 11: 90-99.
Direct Link - Yoshida, S., S. Hiradate, T. Tsukamoto, K. Hatakeda and A. Shirata, 2001. Antimicrobial activity of culture filtrate of Bacillus amyloliquefaciens RC-2 isolated from mulberry leaves. Phytopathology, 91: 181-187.
CrossRefDirect Link - Zhang, Z., S. Schwartz, L. Wagner and W. Miller, 2000. A greedy algorithm for aligning DNA sequences. J. Comput. Biol., 7: 203-214.
CrossRefPubMedDirect Link