
Naceur Djebali
Laboratory of Molecular Physiology of Plants, Centre of Biotechnology of Borj Cedria, B.P. 901, Hammam-Lif 2050, Tunisia
Plant Pathology Journal
Year: 2012 | Volume: 11 | Issue: 3 | Page No.: 77-86







ABSTRACT
This study aimed to determine the effect of seed hydropriming on germination and seedling growth of Triticum durum (durum wheat: cultivar Karim) and Hordeum vulgare (barley: cultivar Souihli), along with its effect on T. durum resistance to Fusarium culmorum seed infection. Seeds of Karim and Souihli were hydroprimed in distilled water at various time intervals till 9.5 h at 25 and 35°C in the dark. The results showed that the pace of water uptake and seedling growth depends on the soaking time and temperature; however the germination percentage depends only on the soaking time. Karim and Souihli seeds absorbed water very fast for up to 30 min with a higher rate at 35°C in comparison to 25°C. Thereafter, a little change was observed in the speed of water uptake for up to 9.5, with a little higher rate at 35°C. The speed of emergence of radicle, coleoptile and side roots and the seedling fresh weight were enhanced in comparison to the control at 5.5-6.5 h and at 3.5 h of seed hydration in Karim and Souihli, respectively. Hydropriming at 25°C gives better seedling fresh weight in both cereals in comparison to 35°C. The water pre-treatment did not affect the level of seed contamination by saprophytic fungi during germination. The hydroprimed Karim seeds inoculated with F. culmorum showed an increase in seedling growth and a reduction in the percentage of infection in comparison to non-hydroprimed seeds. This difference can be attributed in part to the enhanced germination rate and seedling vigour of the hydroprimed seeds.
PDF Abstract XML References Citation
Received: October 01, 2012;
Accepted: December 15, 2012;
Published: January 24, 2013
How to cite this article
Naceur Djebali, 2012. Seed Hydropriming Effect on Triticum durum and Hordeum vulgare
Germination, Seedling Growth and Resistance to Fusarium culmorum. Plant Pathology Journal, 11: 77-86.
DOI: 10.3923/ppj.2012.77.86
URL: https://scialert.net/abstract/?doi=ppj.2012.77.86
DOI: 10.3923/ppj.2012.77.86
URL: https://scialert.net/abstract/?doi=ppj.2012.77.86
INTRODUCTION
The kinetic and amount of seed germination and seedling emergence had a great influence on crop stand and production in arid and semi-arid area. Several pre-sowing seed treatments were used to improve the rate and uniformity of germination in the field to increase crop yield (Khan et al., 2008). Soaking seeds in water or in a salt solution for a specified period of time and then re-dry them just before radicle emerges, known as seed priming (Desai et al., 1997), was successfully used to increase germination and seedling emergence (Joudi and Sharifzadeh, 2006; Amooaghaie, 2011). Seed priming enhances the germination by inducing several biochemical changes such as breaking of dormancy, hydrolysis or mobilization of inhibitors and enzyme activation (Amooaghaie, 2011). The resulting improved stand can increase the drought tolerance, reduce pest damage and pathogen attack and increase crop yield in cereals and legumes (Harris et al., 1999; Khan et al., 2005; Amooaghaie, 2011). Seed priming with water (hydropriming) in maize, rice, alfalfa, pinto bean and chickpea resulted in faster seed emergence, improved establishment, better crop growth, earlier flowering and high yield (Harris et al., 1999; Ghassemi-Golezani et al., 2010). Rashid et al. (2006) noticed that presoaking of barley seeds in water followed by drying back to its original moisture level significantly increased yield of both grain and straw in normal and saline soil. Hydropriming has been considered as a simple and cost-effective strategy to alleviate the NaCl and PEG induced stresses in Brassica juncea (Shrivastava et al., 2010).
Fusarium head blight (FHB) is among the most important diseases of wheat (Goswami and Kistler, 2004). The effects of FHB go beyond yield and kernel quality reductions, as trichothecene mycotoxins produced during infection contaminate raw grain and processed wheat products (Edwards et al., 2009), placing human and livestock health at risk. FHB of wheat is caused by a complex of Fusarium species including F. graminearum, F. culmorum, F. avenaceum, F. poae and F. sporotrichioides (Parry et al., 1995). In Tunisia, F. culmorum is the most prevalent species of the FHB complex (Gargouri et al., 2001). Since cultural measures, use of partially resistant cultivars and use of fungicides provide only partial FHB control, alternative control methods are being explored as an additional tool in the integrated management of this disease. Seed priming can offer a promising method for the control of plant pathogens through the induction of plant defence systems (Worrall et al., 2012).
The objective of this research was to study the effect of different soaking times and two temperatures of seed treatment, on the speed of water uptake, germination and seedling growth of two cereal species durum wheat and barley. In addition, the hydropriming effect on durum wheat resistance to Fusarium culmorum seed infection was also worked out.
MATERIALS AND METHODS
Plant and fungal material: Seeds of the cultivars Karim (durum wheat: Triticum durum L.) and Souihli (Barley: Hordeum vulgare L.) were used free from chemical treatments and were stored at room temperature. The initial seed moisture level was about 10% for both cereal species and the mass of thousand grains was 39.3 and 44.2 g for Karim and Souihli, respectively.
The strain FC3 of Fusarium culmorum used in this study is a Tunisian durum wheat isolate kindly provided by Dr. Samia Gragouri (INRAT, Tunisia). This isolate was maintained on PDA medium at 25°C in the dark. To produce macroconidia for the inoculation tests, the fungal isolate was cultured on Joff’s medium (Dhingra and Sinclair, 1985) at 25°C in the dark for three weeks. This study was conducted in the Laboratory of Molecular Physiology of Plants between October 2010 and March 2012.
Water uptake, hydropriming and germination procedures: The water uptake was measured by putting known weights of undamaged and size uniform seeds in distilled water (pH 7) for 9.5 h at two temperatures 25°C and 35°C in the dark. After different intervals of time, the weight of the water absorbed by the seeds was ascertained by removing the seeds from the water and rapidly drying the surfaces and then weighing. The kinetic of water uptake was monitored every 30 min for 4.5 h and then after every 60 min till 9.5 h. The speed of water uptake (SWU) was calculated as the difference between the fresh weight of soaked seeds (SFW) and their initial dry weight (SDW) divided by the time of soaking (TS) according to the equation:
and expressed as mg min-1. After soaking, the seeds were removed from water and were re-dried to original weight with forced air under shade at 25°C. For germination, thirty five seeds were put with their grooves facing downwards on two layers of sterile filter paper imbibed with sterile distilled water (6 mL; pH 7) in square Petri dishes (12x12 cm) at 25°C in the dark (Essemine et al., 2007).
Fungal inoculation: Karim seeds were put for 3 h in macroconidia suspension of F. culmorum (2x106 macroconidia mL-1) supplemented with 0.01% of Tween 20 (Sigma). The inoculated seeds were placed in square petri dishes on two layers of imbibed filter paper to germinate as previously described.
Measured parameters and statistical analyses: The germination of seeds was monitored each 24 h until 72 h by calculating the Mean Radicle Emergence Time (MRET), the mean side root emergence time (MSRET) and the mean coleoptile emergence time (MCET) according to the equations:
where, n1, n2 and n3 are the numbers of seeds with emerged radicle, side roots and coleoptile, respectively on day D and D is the number of days counted from the beginning of germination test. In addition, the length of shoot (coleoptile±first leaf) and root were measured along with the fresh weight of ten seedlings at 72 h. Final seedling length (SL = shoot+root lengths in cm) and percentage of germination (FG%) were used for Vigour Index (VI) estimation according to Abdul-Baki and Anderson (1973):
VI = FG%xSL |
The percentage of seed contamination by saprophytic fungal species (mainly of the genus Alternaria) was determined. In the assay of resistance to F. culmorum we also determined the percentage of seed infection by this pathogen.
The analysis of variance (ANOVA) and the comparison of means (Duncan multiple range test) of the measured parameters were performed using Statistica software version 5.1 (StatSoft, France). Correlations between the measured parameters were estimated using the same software by computing Pearson’s correlation coefficient (R). The level of significance was set to 5%.
RESULTS
Effect of the soaking time and temperature on the speed of water uptake: The variation in rate of water uptake was significantly influenced by the Cereal Species (CS), soaking temperature (ST°) and soaking time (St) and their interactions, except for CSxSt interaction (Table 1). For the two cereal species, the speed of water uptake increases considerably until 30 min of imbibition in water and then decline to reach a stable stage at 2.5 h (Fig. 1). At 30 min of imbibition, the speed of water uptake was higher in the Souihli seeds in comparison to Karim seeds and it was superior at 35°C in comparison to 25°C (Fig. 1).
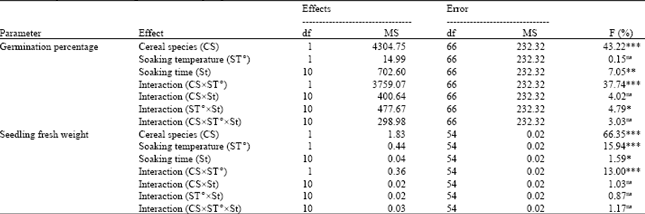
Effect of the soaking time and temperature on the germination percentage: The analysis of variance showed a significant effect of the Cereal Species (CS), the Soaking time (St) and the interactions of CSxST° and ST° x St on the percentage of seeds germination (Table 2). The soaking temperature had no effect on the percentage of seed germination of Karim and Souihli seeds (Table 2). The soaking time had no effect on the level of seed contamination by saprophytic fungi mainly of the genus Alternaria during germination (data not shown).
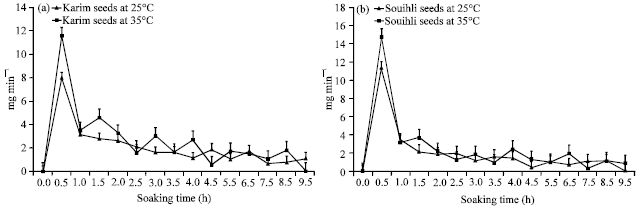 | |
| Fig. 1(a-b): | Speed of water uptake of (a) Karim and (b) Souihli seeds at different soaking times and under two temperature regimes |
| Table 1: | Analysis of variance of the speed of water uptake of Karim and Souihli seeds |
 | |
| F: Index of Fisher-Snedecor, ns: Not significant (p>0.05), *Significant (0.05>p>0.01), **Highly significant (0.01>p>0.001), ***Very highly significant (p<0.001) | |
| Table 2: | Analysis of variance of germination of hydroprimed Karim and Souihli seeds |
 | |
| F: Index of Fisher-Snedecor in percent, ns: Not significant (p>0.05), *Significant (0.05>p>0.01), **Highly significant (0.01>p>0.001), ***Very highly significant (p<0.001) | |
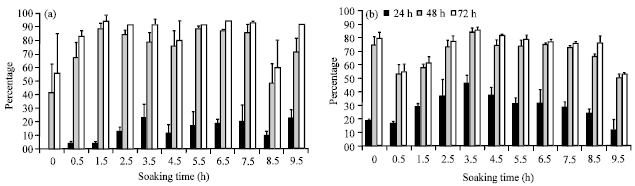 | |
| Fig. 2(a-b): | Percentage of germination of hydroprimed (a) Karim and (b) Souihli seeds at different soaking times |
An early emergence of radicle, coleoptile and side roots of the hydroprimed seeds in comparison to the control was observed in both species (Fig. 2). For Karim seeds, all soaking times gave a superior percentage of germination in the hydroprimed kernels in comparison to the control at 24 h (Fig. 2), whereas for the Souihli kernels an increase in this parameter was observed for the soaking times ranging from 1.5 h to 8.5 h, with an optimal soaking time at 3.5 h (Fig. 2). A slight increase in the percentage of seed germination was noticed for the hydroprimed kernels of durum wheat and barley at 72 h showing a sustained development for the water pre-treated kernels. In addition, the homogeneity of seed germination for the water pre-treated kernels was better in comparison to the non treated kernels. In fact, the Fig. 1 show large standard errors for the germination percentage of the control seeds in comparison to the water pre-treated seeds at the optimal soaking time for Karim (Fig. 2).
Effect of the soaking time on the mean emergence time of radicle, side roots and coleoptile: The mean emergence time of radicle (MRET), side roots (MSRET) and coleoptile (MCET) varied significantly with the soaking time but not with the soaking temperature (data not shown), thus only MRET, RSRET and MCET at 25°C are presented in Fig. 3. In control non-hydroprimed seeds MRET, MSRET were significantly higher in Karim in comparison to Souihli, however the opposite was noticed for MCET (Fig. 3). For Karim hydroprimed seeds MRET, MSRET and MCET declined substantially from 0 to 6.5 h of imbibition and slightly increase then after. For Souihli hydroprimed seeds the previous parameters declined from 0 to 3.5 h of imbibition and increased then after (Fig. 3).
Effect of the soaking time and temperature on seedling growth and vigour index: The effect of hydropriming on the seedling fresh weight was dependent on soaking time and temperature during this process (Table 2, Fig. 4). For the Karim cultivar, we demonstrated that plantlets derived from soaked seeds between 5.5 and 6.5 h had a superior fresh weight in comparison to the other water soaking treatment and to the non treated control (Fig. 4). For the barley cultivar Souihli, an increase in the seedling fresh weight was observed at the soaking time 3.5 h (Fig. 4). At the optimal soaking times for the durum wheat and barley cultivars a superior seedling fresh weight was obtained at 25°C during the water imbibition process in comparison to 35°C (Fig. 4).
The Fig. 5 shows the measure of the radicle, coleoptile and seedling lengths at 72 h at different soaking times for the two cereal species. The results revealed a significant increase in the length of these organs in the hydroprimed seeds in comparison to the control (Fig. 5). Indeed, the water pre-treated Karim seeds (at 6.5 h) showed an increase about 1.8 and 2.4 folds in the length of radicle and coleoptile, respectively in comparison to the control. In addition, the hydroprimed seeds of the Souihli cultivar (at 3.5 h) recorded an increase about 1.7 and 3.9 folds in the radicle and coleoptile lengths respectively in comparison non treated seeds (Fig. 5).
The vigour index of Karim and Souihli seedlings varied function of the soaking time and reached a maximum value at 5.5-6.5 for Karim and at 3.5 h for Souihli (Fig. 6).
Correlations between the parameters of seed water content, seed germination and seedling growth: The correlations between the parameters of seed water content, germination and seedling growth are presented in Table 3. The seed water content was negatively correlated to the mean emergence time of radicle, coleoptile and side roots and positively correlated to the coleoptile length in the durum wheat cultivar Karim. However, this parameter was only negatively correlated to the mean emergence time of coleoptile in the barley cultivar Souihli.
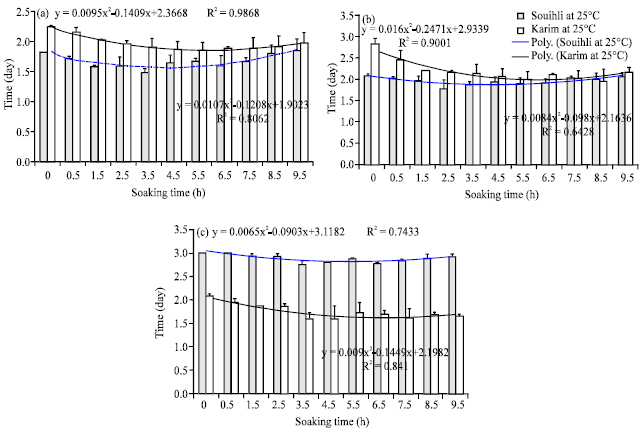 | |
| Fig. 3(a-c): | (a) Means of radicle, (b) Side roots and (c) Coleoptile emergence times of hydroprimed Karim and Souihli seeds at different soaking times |
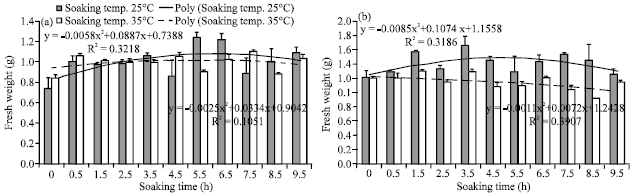 | |
| Fig. 4(a-b): | Fresh weight of (a) Karim and (b) Souihli seedlings derived from hydroprimed seeds at different soaking times and two temperatures |
The mean emergence time of radicle, coleoptile and side roots were positively correlated to each other and negatively correlated to the parameters of seedling growth such as seedling length, fresh weight and vigour index in the two cereal species. The parameter of final germination percentage was positively correlated to the seedling vigour in Karim and Souihli cultivars (Table 3).
Seed hydropriming effect on durum wheat resistance to F. culmorum seed infection: The results of the effect of F. culmorum inoculation on germination and seedling growth of non- and hydroprimed Karim seeds are shown in Table 4. The inoculated hydroprimed seeds showed an increase in the percentage of germination at 24 and 48 h in comparison to the inoculated non-hydroprimed seeds. The mean emergence time of radicle, side roots and coleoptile were significantly lower in the inoculated hydroprimed seeds in comparison to the inoculated control. In addition, seedling length, fresh weight and vigour were significantly higher in water pre-treated Karim seeds. A decrease in the percentage of seed infection with F. culmorum in hydroprimed seeds was observed in comparison to non hydroprimed seeds (Table 4).
| Table 3: | Correlations between the measured parameters of water uptake, germination and seedling growth of Karim (lower half of the matrix) and Souihli (upper half of the matrix) seeds at 25°C in the dark |
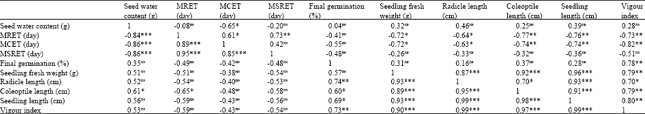 | |
| ns: Not significant (p> 0.05), *Significant (0.05>p>0.01), **Highly significant (0.01>p>0.001), ***Very highly significant (p<0.001) | |
| Table 4: | Effect of Fusarium culmorum on germination and seedling growth of non-hydroprimed (control) and hydroprimed Karim seeds |
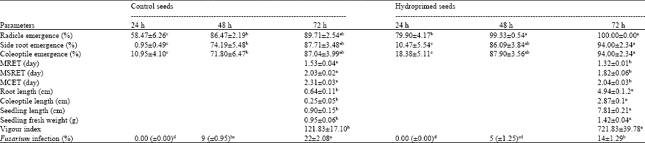 | |
| Values (Mean±SE) at the row level followed by the same letter are not significantly different at p≤0.05 | |
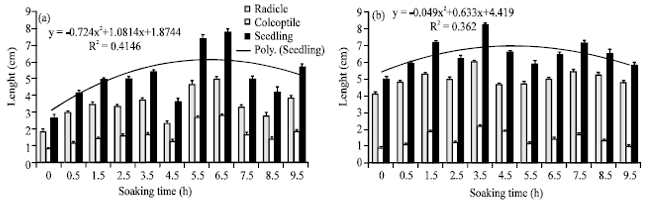 | |
| Fig. 5(a-b): | Measure of radicle, coleoptile and seedling lengths of (a) Karim and (b) Souihli plants derived from hydroprimed seeds at different soaking times |
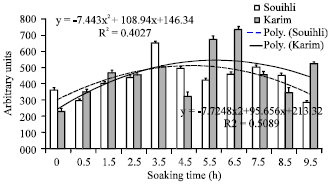 | |
| Fig. 6: | Vigour index of Karim and Souihli seedling derived from hydroprimed seeds at different soaking times |
DISCUSSION
In the field, the rate of germination and emergence of sown seeds affect the growth and yield of plants. In addition, physiological characteristics of seed influence plant resistance to biotic and abiotic stress conditions. So, increasing seed performance constitutes a promising way to enhance crop production and resistance to several diseases. Hydropriming is a simple method of pre-sowing seed treatment which does not require any particular technical equipment, except the use of distilled water as priming medium (Fujikura and Karssen, 1995). This study investigated whether seed hydropriming can improve germination and seedling growth in wheat and barley and resistance of wheat to F. culmorum seed infection.
Imbibition consists in water absorption and activation of nutritive substances contained in seeds, which is required for the initiation and completion of seed germination. This study showed that the speed of water uptake during imbibition was higher in barley in comparison to durum wheat seeds (Fig. 1). Similarly, Essemine et al. (2007) found that the seed water uptake differs between two wheat species (T. aestivum and T. durum) at various temperatures. Rahman et al. (2011) showed that the amount of water to be absorbed for seed germination depends on species and varieties. King (1984) found that water absorption by kernels of different wheat varieties is conditioned by spike and grain structure. The kinetic of water uptake by both cereal seeds showed two phases. The first phase was characterized by a rapid and short lasting water uptake during the first hour of imbibition, then after a second phase characterized by slow and long lasting water uptake was observed (Fig. 1). Accordingly, Bewley and Black (1994) found that hard wheat seeds showed two phases of water uptake, the first correspond to a rapid entry of water into the apoplast and the second correspond to a slow entry of water that transit through the cellular membrane of seed. It has been also determined that the rate of seed imbibition is temperature-dependent. The rate of imbibition increases with increasing of temperature in many crop seeds such as wheat (Essemine et al., 2007), sorghum (Kader and Jutzi, 2002), amaranth grain (Resio et al., 2006), cowpea (Kaptso et al., 2008) and maize and chickpea (Rahman et al., 2011). Murphy and Noland (1982) showed that the temperature effect on seed imbibition depends from the seed membrane properties and water viscosity. Imbibition temperature did not influence final germination percentage, but significantly influences early seedling growth in durum wheat and barley (Table 2). A better durum wheat and barley seedling growth was obtained at moderate (25°C) in comparison to high imbibition temperature (35°C). Booth and Bai (1999) found that the seedling length of most crops was favoured by imbibition temperatures between 20 and 30°C. In addition, increasing the soaking duration of seeds i.e., the seed water content did not influence the final germination percentage, but it increased the speed of emergence and length of coleoptile in durum wheat and barley (Table 3). Similarly, Mendez-Natera et al. (2008) did not found any relationship between imbibition rate and seed germination in maize, French bean and pigeon pea.
The evidence in the present study suggests an important role of water pre-treatment of durum wheat and barley seeds to enhance germination and seedling growth. The optimal hydropriming time for wheat and barley seeds were 5.5-6.5 and 3.5 h, respectively, which enhanced the germination percentage, decreased the speed of germination (MRET), induce an early development (RSRET and MCET), increased the seedling fresh weight and vigour at 25°C. Ahmadi et al. (2007) found that hydropriming of wheat (Triticum aestivum) seeds clearly improved speed of emergence; vigour index and seedling dry weight. Also, Joudi and Sharifzadeh (2006) demonstrated that seeds hydropriming of three barley cultivars improved germination percentage and rate, length of coleoptiles and of the longest root, dry weight of root and shoot as well as seed vigour index in control and moisture limitation and low temperature conditions. The effect of priming on improving seed performance might be attributable in part to early DNA replication (Bray et al., 1989), increased RNA and protein synthesis (Fu et al., 1988), greater ATP availability (Mazor et al., 1984), faster embryo growth (Chang et al., 2000), repair of deteriorated seed parts (Karssen et al., 1989), reduced leakage of metabolites (Ward and Powell, 1983), decreased in lipid peroxidation and increased in the antioxidant activities (Issam et al., 2012) compared with control. It was also noted from this study that hydroprimed seeds had better homogeneity in seed germination. Our results are in agreement with previous works that demonstrate that hydropriming of the seed of several crops improves uniformity of germination and emergence and enhance plant establishment in the field (Harris et al., 1999; Moradi and Younesi, 2009), which offer several benefits for cereal growers to easy manage and harvest the crop.
This study demonstrated that water pre-treatment of durum wheat and barley seeds had no effect on the percentage of seed contamination with saprophytic fungi mainly of the genus Alternaria (data not shown) during germination. So, the hydropriming treatment will not increase the risk of seed fungal contamination during storage and will not disturb the antagonistic fungal community that may control pathogens on the seed surface. Indeed, Mullenborn et al. (2008) demonstrated that disturbing the saprophytic fungal community on the wheat seeds treated with fungicides may lead to the decrease of antagonistic fungi species which allow the development of several pathogens that are not controlled with the used fungicide.
This work constitutes the first report of the effect of hydropriming on F. culmorum infection of durum wheat seeds during germination. The hydroprimed Karim seeds showed a better percentage of germination and seedling growth and a decrease in the percentage of seed infection in comparison to the non water-pretreated seeds at 3 day post inoculation with F. culmorum. The observed decrease of Fusarium infection in hydroprimed seeds can be attributed in part to enhanced germination rate and seedling vigour. Nevertheless, several works showed that hydropriming of plant seeds altered the expression of genes encoding regulated enzymes in the abscisic acid, gibberellin and ethylene biosynthetic pathways (Schwember and Bradford, 2010), which are known to regulate plant response to biotic and abiotic stresses (Anderson et al., 2004; Bari and Jones, 2009). In addition, it was shown that hydroprimed seedling plants over expressed the activity of antioxidant enzymes which protect cells against the Reactive Oxygen Species (ROS) damage associated to the pathogen infection of plant tissues (De Gara et al., 2003; Djebali et al., 2007; Djebali et al., 2011).
In conclusion this work showed that hydropriming of wheat and barley seeds enhance germination and early seedling growth which determine a good stand establishment and consequently a better yield. In addition, we showed that hydropriming of wheat seeds reduced the percentage of seed infection by F. culmorum 3 days post inoculation. The obtained results may be a basis for improving biotic stress tolerance in plants and particularly in cereals.
ACKNOWLEDGMENTS
This work was funded by the Tunisian Ministry of Higher Education and Scientific Research. The author is grateful to Dr. Aida Bouajila for providing seeds of the barley cultivar Souihli.
REFERENCES
- Abdul-Baki, A.A. and J.D. Anderson, 1973. Vigor determination in soybean seed by multiple criteria. Crop Sci., 13: 630-633.
CrossRefDirect Link - Ahmadi, A., A.S.S. Mardeh, K. Poustini and M.E. Jahromi, 2007. Influence of osmo and hydropriming on seed germination and seedling growth in wheat (Triticum aestivum L.) cultivars under different moisture and temperature conditions. Pak. J. Biol. Sci., 10: 4043-4049.
CrossRefPubMedDirect Link - Amooaghaie, R., 2011. The effect of hydro and osmopriming on alfalfa seed germination and antioxidant defenses under salt stress. Afr. J. Biotechnol., 10: 6269-6275.
Direct Link - Anderson, J.P., E. Badruzsaufari, P.M. Schenk, J.M. Manners and O.J. Desmond et al., 2004. Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell, 16: 3460-3479.
CrossRefDirect Link - Bari, R. and J.D.G. Jones, 2009. Role of plant hormones in plant defence responses. Plant Mol. Biol., 69: 473-488.
CrossRefDirect Link - Booth, D.T. and Y. Bai, 1999. Imbibition temperature affects on seedling vigor: In crops and shrubs. J. Range Manage., 52: 534-538.
Direct Link - Kaptso, K.G., Y.N. Njintang, A.E. Komnek, J. Hounhouigan, J. Scher and C.M.F. Mbofung, 2008. Physical properties and rehydration kinetics of two varieties of cowpea (Vigna unguiculata) and bambara groundnuts (Voandzeia subterranea) seeds. J. Food Eng., 86: 91-99.
CrossRef - Chang, S., C. Tan, E.N. Frankel and D.M. Barrett, 2000. Low-density lipoprotein antioxidant activity of phenolic compounds and polyphenol oxidase activity in selected clingstone peach cultivars. J. Agric. Food Chem., 48: 147-151.
CrossRefDirect Link - De Gara, L., M.C. de Pinto and F. Tommasi, 2003. The antioxidant systems vis-a-vis reactive oxygen species during plant-pathogen interaction. Plant Physiol. Biochem., 41: 863-870.
CrossRefDirect Link - Djebali, N., H. Mhadhbi, C. Jacquet, T. Huguet and M.E. Aouani, 2007. Involvement of hydrogen peroxide, peroxidase and superoxide dismutase in response of Medicago truncatula lines differing in susceptibility to Phoma medicaginis infection. J. Phytopathol., 155: 633-640.
CrossRef - Djebali, N., H. Mhadhbi, C. Lafitte, B. Dumas, M.T. Esquerre-Tugaye, M.E. Aouani and C. Jacquet, 2011. Hydrogen peroxide scavenging mechanisms are components of Medicago truncatula partial resistance to Aphanomyces euteiches. Eur. J. Plant Pathol., 131: 559-571.
CrossRef - Edwards, S.G., B. Barrier-Guillot, P.E. Clasen, V. Hietaniemi and H. Pettersson, 2009. Emerging issues of HT-2 and T-2 toxins in European cereal production. World Mycotoxin J., 2: 173-179.
CrossRefDirect Link - Essemine, J., S. Ammar, N. Jbir and S. Bouzid, 2007. Sensitivity of two wheat species's seeds (Triticum durum, Variety Karim and Triticum aestivum, Variety Salambo) to heat constraint during germination. Pak. J. Biol. Sci., 10: 3762-3768.
CrossRefPubMedDirect Link - Fu, J.R., S.H. Lu, R.Z. Chen, B.Z. Zhang, Z.S. Liu and D.Y. Cai, 1988. Osmoconditioning of peanut (Arachis hypogea L.) seeds with PEG to improve vigour and some biochemical activities. Seed Sci. Technol., 16: 197-212.
Direct Link - Fujikura, Y. and C.M. Karssen, 1995. Molecular studies on osmoprimed seeds of cauliflower: A partial amino acid sequnence of a vigoure-related protein and osmopriming-enhanced expression of putative aspartic protease. Seed Sci. Res., 5: 177-181.
CrossRef - Gargouri, S., M.R. Hajlaoui A. Gurmech and M. Marrakchi, 2001. Identification des especes fongiques associees a la pourriture du pied du ble et leur repartition selon les etages bioclimatiques [Identification of fungal species associated with foot rot of wheat and study their distribution by bioclimatic zones in Tunisia]. OEPP Bull., 31: 499-503.
CrossRefDirect Link - Ghassemi-Golezani, K., A. Chadordooz-Jeddi, S. Nasrollahzadeh and M. Moghaddam, 2010. Effects of hydro-priming duration on seedling vigour and grain yield of pinto bean (Phaseolus vulgaris L.) cultivars. Notulae Bot. Horti Agrobot. Cluj, 38: 109-113.
Direct Link - Goswami, R.S. and H.C. Kistler, 2004. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol., 5: 515-525.
CrossRefDirect Link - Harris, D., A. Joshi, P.A. Khan, P. Gothkar and S.P. Sodhi, 1999. On-farm seed priming in semi-arid agriculture: Development and evaluation in maze, rice and chickpea in India using participatory methods. Exp. Agric., 35: 15-29.
CrossRefDirect Link - Issam, N., M. Kawther, M. Haythem and J. Moez, 2012. Effects of CaCl2 pretreatment on antioxidant enzyme and leaf lipid content of faba bean (Vicia faba L.) seedlings under cadmium stress. Plant Growth Regul., 68: 37-47.
CrossRef - Joudi, M. and F. Sharifzadeh, 2006. Investigation of hydropriming effects on barley cultivars. Sci. Infom. Database, 11: 99-109.
Direct Link - Kader, M.A. and S.C. Jutzi, 2002. Temperature, osmotic pressure and seed treatments influence imbibition rates in sorghum seeds. J. Agron. Crop Sci., 188: 286-290.
CrossRefDirect Link - Khan, A., S.K. Khalil, A.Z. Khan, K.B. Marwat and A. Afzal, 2008. The role of seed priming in semi-arid area for mungbean phenology and yield. Pak. J. Bot., 40: 2471-2480.
Direct Link - King, R.W., 1984. Water uptake in relation to pre-harvest sprouting damage in wheat: Grain characteristics. Aust. J. Agric. Res., 35: 337-345.
CrossRef - Mazor, L., M. Perl and M. Negbi, 1984. Changes in some ATP-dependent activities in seeds during treatment with polyethyleneglycol and during the redrying process. J. Exp. Bot., 35: 1119-1127.
CrossRef - Mendez-Natera, J.R., J.F.M. Pinto and N.J.M. Mata, 2008. Relationship between imbibition rate and seed germination of corn, French bean and pigeonpea seeds. Rev. Cient. UDO Agric., 8: 61-66.
Direct Link - Moradi, A. and O. Younesi, 2009. Effects of osmo- and hydro-priming on seed parameters of grain sorghum (Sorghum bicolor L.). Aust. J. Basic Applied Sci., 3: 1696-1700.
Direct Link - Mullenborn, C., U. Steiner, M. Ludwig and E.C. Oerke, 2008. Effect of fungicides on the complex of Fusarium species and saprophytic fungi colonizing wheat kernels. Eur. J. Plant Pathol., 120: 157-166.
CrossRef - Murphy, J.B. and T.L. Noland, 1982. Temperature effects on seed imbibition and leakage mediated by viscosity and membranes. Plant Physiol., 69: 428-431.
Direct Link - Parry, D.W., P. Jenkinson and L. McLeod, 1995. Fusarium ear blight (Scab) in small grain cereals-A review. Plant Pathol., 44: 207-238.
CrossRefDirect Link - Rahman, M.M., K.U. Ahammad and M.M. Alam, 2011. Effect of soaking condition and temperature on imbibition rate of maize and chickpea seeds. Res. J. Seed Sci., 4: 117-124.
CrossRef - Rashid, A., P.A. Hallington, D. Harris and P. Khan, 2006. On-farm seed priming for barley on normal, saline, saline-sodic soils in North West Frontier Province, Pakistan. Eur. J. Agron., 24: 276-281.
CrossRef - Resio, A.C., R.J. Aguerre and C. Suarez, 2006. Hydration kinetics of amaranth grain. J. Food Eng., 72: 247-253.
CrossRef - Schwember, A.R. and K.J. Bradford, 2010. Quantitative trait loci associated with longevity of lettuce seeds under conventional and controlled deterioration storage conditions. J. Exp. Bot., 61: 4423-4436.
CrossRef - Srivastava, A.K., V.H. Lokhande, V.Y. Patade, P. Suprasanna, R. Sjahril and S.F. D'Souza, 2010. Comparative evaluation of hydro-chemo-and hormonalpriming methods for imparting salt and PEG stress tolerance in Indian mustard (Brassica juncea L.). Acta Physiol. Plant, 32: 1135-1144.
Direct Link - Ward, F.H. and A.A. Powell, 1983. Evidence for repair processes in onion seeds during storage at high seed moisture contents. J. Exp. Bot., 34: 277-282.
CrossRef - Worrall, D., G.H. Holroyd, J.P. Moore, M. Glowacz and P. Croft et al., 2012. Treating seeds with activators of plant defence generates long-lasting priming of resistance to pests and pathogens. New Phytol., 193: 770-778.
CrossRef