
Anshu Sharma
Department of Biotechnology, University of Horticulture and Forestry, Solan-173230, Himachal Pradesh, India
Surinder Kumar
Department of Biotechnology, University of Horticulture and Forestry, Solan-173230, Himachal Pradesh, India
Minerva Ghani
Department of Biotechnology, University of Horticulture and Forestry, Solan-173230, Himachal Pradesh, India
Manisha Thakur
Department of Biotechnology, University of Horticulture and Forestry, Solan-173230, Himachal Pradesh, India
Harender Raj
Department of Mycology and Plant Pathology, University of Horticulture and Forestry, Solan-173 230, Himachal Pradesh, India
Plant Pathology Journal
Year: 2012 | Volume: 11 | Issue: 2 | Page No.: 60-67







ABSTRACT
The demand for lily flowers has increased considerably during the last few decades, the main limiting factor in large scale cultivation is its susceptibility to a number of pathogens. The present investigation was undertaken to improve this crop by developing resistant mutant lines using in vitro cell selection technique. Resistant cell lines were selected by culturing calli derived from bulbscale segments of Oriental lily ‘Casa Blanca’ susceptible to Fusarium oxysporum on MS (Murashige and Skoog) medium supplemented with growth regulators and various concentrations of culture filtrate. After one month of inoculation about 94% of the calli showed reduction in growth. Selection was made at 15% culture filtrate level. Only 6.6% calli regenerated shoots with different concentrations of Naphthalene Acetic Acid (NAA) and Benzyl adenine (BA). Resistant calli obtained after two cycles (30 day cycle-1) of selection were used for bulblet regeneration and to check the stability the regenerated bulblets were transferred to MS medium containing 50% culture filtrate. About 75% of the resistant bulblets showed healthy growth and produced new leaves. One-year-old bulbs from selected cell lines and control were maintained in glasshouse for screening against pathogen. Thirty three plants from selected cell lines showed resistance whereas all the control plants showed morbidity or died when tested with Fusarium spore suspension. No variation was observed in the morphological characters in the selected regenerates. Resistant bulbs were developed using in vitro selection technique, which may be one of the main and effective ways of lily variation breeding with resistance to both biotic and abiotic stresses.
PDF Abstract XML References Citation
Received: April 18, 2012;
Accepted: June 09, 2012;
Published: August 15, 2012
How to cite this article
Anshu Sharma, Surinder Kumar, Minerva Ghani, Manisha Thakur and Harender Raj, 2012. Development of Mutant Lines of Oriental Hybrid Lily Resistant to Fusarium
oxysporum f.sp. lilii Toxin(s). Plant Pathology Journal, 11: 60-67.
DOI: 10.3923/ppj.2012.60.67
URL: https://scialert.net/abstract/?doi=ppj.2012.60.67
DOI: 10.3923/ppj.2012.60.67
URL: https://scialert.net/abstract/?doi=ppj.2012.60.67
INTRODUCTION
Lilium is one of the most important ornamental crops all over the world, ranking the fourth among the flowers with annual sale of 159 million Euros (Anonymous, 2009; Raj, 2010). It has a wide applicability in the floral industry as a cut flower and pot plant (Xia et al., 2006). The demand for lily cut flowers increased considerably during the past few years. The global production of lily bulbs occurs in 10 countries with the largest production area of 4,280 ha and a total production of 2.21 billion lily bulbs in the Netherlands (Benschop et al., 2010). Lilies can be propagated by both sexual and asexual methods. Most commercially grown cultivars are propagated through vegetative means viz., bulblets, bulbils and adventitious bulblet formation on bulb scales (Kumar et al., 2006). Numerous studies have been conducted on in vitro regeneration of bulblets in lily using different explants (Bacchettal et al., 2003; Nhut, 2003; Khawar et al., 2005; Kumar et al., 2006; Azadi and Khosh-Khui, 2007; Xu et al., 2009; Joshi and Dhar, 2009; Saifullah et al., 2010). Though the demand for lily flowers has increased considerably during the past few decades, the main limiting factor in large scale cultivation is its susceptibility to a number of pathogens, including fungi, bacteria and viruses. Fungal diseases are the most destructive world-wide and are responsible for causing huge loss by reducing the yield and quality of flowers (Hammond et al., 2006).
Among fungal diseases, Fusarium wilt is caused by Fusarium oxysporum, which is also one of the most destructive pathogens in a number of crops (Thakur et al., 2002; Lim et al., 2005; Nasir and Riazuddin, 2008; Amini, 2009; Sallam and Abdel-Monaim, 2012). This pathogen causes wilting and bulb rot in Lilium under field and storage conditions resulting up to 50-100% loss (Anonymous, 2006). Prados-Ligero et al. (2008) reported for the first time on the occurrence of F. oxysporum f.sp. lilii and Fusarium proliferatum in Spain resulting in 80-90% of Lilium plants/bulbs malformation. The fungal diseases in Lilium are mostly managed by using a variety of fungicides (Jana and Roy Choudhary, 1989; Sunita et al., 2002), however, these chemicals are expensive and not eco-friendly. Moreover, chemical control of fungal diseases is difficult due the absence of highly effective fungicides (Loffler and Mouris, 1992). To overcome this problem, attempts have been made to induce resistance through conventional breeding approaches (Straathof and Loffler, 1994; Straathof and van Tuyl, 1994; Van Tuyl and van Holsteijn, 1996), which are time consuming and expensive (Garibaldi and Gullino, 1987; Liu et al., 2005). It is therefore, imperative that novel methods such as mutagenesis, in vitro cell selection and genetic transformation with desirable genes be applied for inducing variability in the existing germplasm. Ding et al. (2011) obtained anti-Fusarium clones of lily Acapulco and Casa Blanca by adding various concentrations of toxin in MS medium. A high level of Fusarium resistance in some lily cultivars, especially Asiatic hybrid lilies and none in oriental hybrids has already been reported (Loffler et al., 1996). The present study was performed to develop mutant lines through in vitro selection of oriental hybrid lily cv. ‘Casa Blanca’ resistant to Fusarium oxysporum f.sp. lilii, the causal agent of fusarium wilt.
MATERIALS AND METHODS
Explant source and preparation: Vernalized bulbs of oriental lily hybrid `Casa Blanca’ were procured from the Department of Floriculture and Landscaping, University of Horticulture and Forestry, Solan, India during the year 2008. Scales were separated and washed under running tap water for 30 min. The basal and apical portions of the scale were trimmed and the middle portion of the scale (3-4 mm) was used as explant. These were washed in 1% Tween-20 (v/v) solution followed by 3-4 washing with distilled water. The explants were surface-sterilized by dipping in 0.1% mercuric chloride solution for 4-5 min followed by thorough washing with sterile distilled water.
Culture medium and culture conditions: One hundred sterilized explants were cultured with the abaxial side down on solidified MS (Murashige and Skoog, 1962) medium supplemented with 1.5 mg L-1 benzyladenine (BA), 2 mg L-1 Naphthalene Acetic acid (NAA) and 5% (w/v) sucrose for callus induction. The pH of the medium was adjusted to 5.8 prior to adding 0.8% (w/v) Difco bacto agar. The medium was dispensed in 30 mL aliquots into 100 mL Erlenmeyer flasks (Borosil, Mumbai, India), which were plugged with non-absorbent cotton plugs. Medium was autoclaved at 1.1 kg cm-2 for 15 min at 121°C. The cultures were maintained in dark at a temperature of 24±2°C for the initiation of callus. The cultures were transferred at four weeks interval to fresh medium with the same composition.
Maintenance and multiplication of the pathogen culture: The pure cultures of Fusarium oxysporum f.sp. lilii (local isolates) were obtained from the Department of Mycology and Plant Pathology, University of Horticulture and Forestry, Solan. The isolates were multiplied at 25°C on PDA (200 g L-1 potato, 20 g L-1 dextrose, 20 g L-1 agar) medium. The pH of the PDA medium was 6.0-6.5. After inoculation, the cultures were incubated at 25°C in dark for four days until a uniform fluffy mycelial growth was obtained. The stock cultures were maintained at 4°C on the same medium.
Preparation of culture filtrate: The culture filtrate (CF) was prepared by inoculating 2 mm2 bit of the fungal mycelium in 100 mL liquid Czapek Dox medium (Czapek, 1922) containing 30 g L-1 sucrose, 2 g L-1 sodium nitrate, 1 g L-1 dipotassium hydrogen phosphate, 0.5 g L-1 magnesium sulphate, 0.5 g L-1 potassium chloride, 0.01 g L-1 ferrous sulphate, pH 6.6-7.2. The culture filtrate was then incubated in BOD incubator shaker at 25°C in the dark. After 21 d, the fungal cultures were filtered through Whatman filter paper No. 1. The filtrate was centrifuged at 20,000 x g for 30 min at 25°C while maintaining the pH at 5.6 with 1 N HCl and/or 1 N NaOH. The supernatant was then filtered through Whatman filter paper No. 42 and finally through sintered glass filter (G-5 grade, 0.25 μM pore size) to produce the CF, which was transparent in appearance.
Testing of toxicity of the CF: The callus was divided into small pieces and macerated with 0.5% macerozyme (Sigma-Aldrich, Bangalore, India) for 30 min in the dark. The cell clumps were separated through a nylon sieve (64 μM, pore size). After washing 4 times with Czapek Dox medium, the macerated cells were again separated through nylon sieve (64 μM, pore size) and incubated in Czapek Dox medium and CF (undiluted). After 48 h, cells were stained with 0.1% erythrocin B (which stains only dead cells) and observed under a light microscope.
Cell selection: Fifty pieces of callus (10 mg callus-1) in five petri plates (each plate having 10 pieces) for each culture filtrate treatment were inoculated on MS medium containing 0, 2.5, 5.0, 7.5, 10.0, 12.5, 15.0, 16 and 17% (v/v) CF and the optimum concentrations of growth regulators used for callus induction (2 mg L-1 NAA+1.5 mg L-1 BA). In the control cultures, CF was replaced by an equal volume of Czapek Dox medium. Cultures were incubated at 25°C under 16/8 h photoperiod with light intensity of 50-60 μmol m-2 sec-1. Light green calli obtained after incubation on selection medium containing 15% CF, the highest concentration of CF at which only few calli survived were subcultured onto the same selection medium for two more cycles (30 day cycle-1) to produce resistant calli. These calli were later multiplied on callus multiplication medium (2 mg L-1 NAA+1.5 mg L-1 BA) with 15% CF.
Bulblet regeneration and analysis of resistibility: The selected calli were inoculated on MS basal medium containing 1, 1.5 or 2 mg L-1 NAA and 0.5, 1.5 or 2 mg L-1 BA to regenerate bulblets. The regenerated bulblets were multiplied by separating the individual bulblets on the same medium. The bulblets were inoculated on the culture medium in the absence of pathogen culture filtrate for 3 subcultures to get rid of physiological adapted resistance. Then 50 bulblets regenerated from selected calli, were cultured on MS medium with 50% pathogen culture filtrate to test the stability of resistance. There were three replicates for each treatment. Bulblets derived from cultures without culture filtrate were used as control.
Bulblet storage: The individual bulblet was transferred on MS medium without growth regulators. When the leaves dried, bulblets obtained from each of the control and selected lines were taken out of the culture vessels, washed thoroughly and dried at room temperature. The bulblets were treated with 0.1% Bavistin (Indofil, Mumbai, India) and stored at 2°C in coco peat.
Disease resistance identification under in vivo conditions: Bulbs (one year-old) from control and selected lines were transferred to the beds in glass house. The beds were prepared by mixing sterile sand and soil in 1:1 ratio. Thirty bulbs of control and 60 bulbs of selected lines were maintained for screening against pathogen. After the bulbs sprouted, these were subjected to infection under in vivo conditions by spraying freshly prepared spore suspension (prepared by dissolving 1 mm2 bit of Fusarium mycelium in distilled water in such a way to keep the spore concentration at 40 spores m L-1). The disease resistance of the control and selected lines resistant to spore suspension was assessed by the living state and survival rate after 30 days of inoculation with Fusarium oxysporum spores suspension. Disease severity was recorded according to scale with group categories 1 through 6 (Straathof et al., 1993):
| • | Group 1 = Healthy |
| • | Group 2 = Slightly rotten |
| • | Group 3 = Moderately rotten |
| • | Group 4 = Heavily rotten |
| • | Group 5 = Very heavily rotten |
| • | Group 6 = Completely decayed |
Statistical analysis: The experiment was repeated three times. The data were analysed statistically using completely randomized design (Gomez and Gomez, 1984). The statistical analysis based on mean value per treatment was made using the technique of analysis of variance. The comparative LSD test (p≤0.05) was used to determine difference between treatment means.
RESULTS
Induction of callus and confirmation of toxicity of culture filtrate: 60% of the bulb scale segments cultured on MS medium supplemented with 1.5 mg-1 BA, 2 mg L-1 NAA and 5% sucrose gave best callus induction and growth (Fig. 1). Repeated subculturing was done on the same medium to obtain friable calli required for the selection of resistant lines. Single macerated cells incubated in CF and Czapek Dox medium containing 0.1% erythrocin B showed that all the cells incubated in CF did stain after 48 h however, no staining was observed in the cells incubated in liquid Czapek Dox medium, thus proving the toxicity of CF to the lily cells.
Effect of culture filtrate on the development of resistant callus: On the culture medium with no culture filtrate, higher callus growth rate was obtained. The rate of callus growth decreased significantly (p≤0.05) as the concentration of CF increased (Fig. 2).
 | |
| Fig. 1: | Callus induction from bulbscale segments cultured on MS medium supplemented with 1.5 mg L-1 BA, 2 mg L-1 NAA and 5% sucrose after 60 days of inoculation |
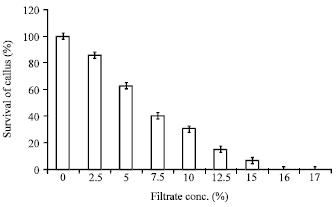 | |
| Fig. 2: | Effect of different concentrations of pathogen culture filtrate on growth and survival of callus |
| Table 1: | Effect of growth regulators on calli regenerating bulblets and number of bulblets per calli |
 | |
| Numbers within parentheses are arc sine transformed values, Experiment was repeated three times, 25 pieces of callus were maintained for each treatment NAA: Naphthalene acetic acid, BA: Benzyladenine | |
| Table 2: | Analysis of resistibility of the selected bulblets to 50% pathogen culture filtrate |
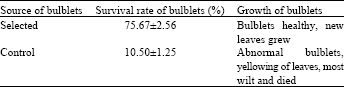 | |
| Bulblets were obtained from selected calli, Selection was made at 15% concentration of pathogen culture filtrate±standard error | |
 | |
| Fig. 3: | Browning of calli at higher concentration (16%) of pathogen culture filtrate |
The frequency of survival of calli at 0-15% of the culture filtrate stress was 100, 85.6, 62.7, 40, 30.5, 15 and 6.6%, respectively while that of 16% and above was nil. The cell selection was therefore carried out at 15% CF level to exert maximum selection pressure that allowed the recovery of resistant lines. The average number of 3.3 surviving calli were again cultured at 15% CF for two more cycles (30 day cycle-1) before the bulblets were regenerated. It was observed that higher the concentration of crude pathogen culture filtrate was, the more the browning response (Fig. 3). It showed that the CF had a strong inhibition of the callus growth and it could be used to screen mutant lines resistant to fusarium wilt.
Differentiation of selected callus cultures: The calli selected at 15% of CF were transferred to multiplication medium (1.5 mg L-1 BA and 2 mg L-1 NAA). After 30 days, 100 pieces of calli (25 pieces treatment-1; 2 mm2) were cultured on bulblet regeneration medium (Fig. 4). The maximum calli (75%) regenerating bulblets was observed in medium containing 2 mg L-1 NAA and 1.5 mg L-1 BA (Table 1).
 | |
| Fig. 4: | Bulblet regeneration from resistant calli obtained after selection against 15% of culture filtrate |
The percentage calli producing bulblets on each medium did not differ with each other. Although the bulblets were produced on all media, the maximum average number of bulblets (6.0) was produced on medium containing 2 mg L-1 NAA and 1.5 mg L-1 BA, which was statistically on a par with media containing 1.5 mg L-1 NAA and 1.5 mg L-1 BA.
Resistibility of in vitro regenerated bulblets: The results of resistibility of the control and selected bulblets are shown in Table 2. The control bulblets did not grow normally, yellowing of leaves and even wilt and died. The survival rate was 10% only. However, selected bulblets showed good growth and grew new leaves with survival rate of 75% when tested against 50% pathogen culture filtrate. The results indicated that the culture filtrate selected bulblets showed a stable resistance to high level of culture filtrate stress.
Disease resistance under in vivo conditions: The Fusarium wilt resistance of the selected plants was greatly improved as compared to control plants under the same disease stress conditions. There were 33 plants regenerated (Fig. 5) from the resistant bulbs belonged to group 1 and the rest fall in the groups 3, 4, 5 and 6. In control, all the plants showed morbidity and fall in the groups 4, 5 and 6 (Table 3).
 | |
| Fig. 5: | Resistant plants raised from one year-old resistant bulbs in beds in the glasshouse |
| Table 3: | Disease resistance identification of the regenerated plants under glasshouse |
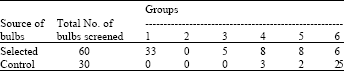 | |
The results indicated that in vitro screening of Lilium resistant mutants for Fusarium wilt is possible and is an important compliment to classical breeding methods.
DISCUSSION
With the progress in biotechnology, new methods in plant pathology have been developed, which elucidated the relationship between in vitro and in vivo mechanism of resistance and tried to develop practical applications to obtain resistant breeding lines.
Sustainable farming system strives to minimize the use of synthetic pesticides and to optimize the use of alternative management strategies to control soil borne pathogens. The use of crude pathogen culture filtrate or crude toxin to induce disease resistant plant by in vitro culture has proven an effective way of plant mutation selection or breeding in plants (Zhang et al., 2012). Somaclonal breeding programmes have proved extremely successful in developing disease-resistant cultivars in a number of crop species (Kintzios et al., 1996; Thakur et al., 2002; Svabova and Lebeda, 2005). It has either been attempted at the plant level where a large population of plants, raised through in vitro callus cultures, have been screened for resistance in the field directly or via a more targeted approach of regeneration of disease resistant plants through resistant callus cultures selected against fungal toxins (Saxena et al., 2008). The results obtained in the present study from in vitro experiments revealed that the bulb scale segments cultured on MS medium supplemented with 5% sucrose, 2 mg L-1 NAA and 1.5 mg L-1 BA, placed in dark for two months gave best callus growth. Repeated sub culturing was done to obtain friable callus required for the selection of resistant lines.
It has been reported that the culture filtrate showed the presence of extracellular non-specific phytotoxins that determine pathogenicity (Pennisi et al., 1988; Sesto et al., 1990). A correlation is must between the in vitro and in vivo responses of the host to the infection caused by the pathogen toxin used for selection of disease-resistant genotypes. In the present study, the toxicity of the culture filtrate was confirmed by macerating single cell in CF and Czapek Dox medium with 0.1% erythrocin B. The cells did stain after 48 h, proving the toxicity of CF to the lily cells. It was further supported by the fact that the pathogen culture filtrate of Fusarium oxysporum has clearly negative effect on the survival and growth of the callus. The inhibitory effects become intensified with increase in the concentration of pathogen culture filtrate. The frequency of survival of calli at 0-15% of culture filtrate was 100, 85.6, 62.7, 40, 30.5%, 15 and 6.6%, respectively while the 16% and above was nil. Cell selection was therefore carried out at 15% CF to affect the maximum selection pressure that would allow the recovery of resistant lines. These results suggested that the CF contains toxic metabolite(s) produced by Fusarium oxysporum and may be used as an effective screening agent in a cell selection program. F. oxysporum f.sp. lilii produces toxic metabolites, which is the most important mechanism of this pathogen for killing the host tissue (Loffler and Mouris, 1992). Similarly, a number of previous studies have also found that toxins present in the CF were able to inhibit cell growth and that the cells of the host species were sensitive to the toxins than those of the non-host plants (Hartman et al., 1984; Selvapandiyan et al., 1988).
Tissue culture involves the selection of callus culture in vitro, taking the advantages of the vast somaclonal variations (Scowcroft and Larkin, 1982). These variations may be the result of calli developed from a variety of diploid and polyploids resulting in considerable variations (Hansen et al., 2007) responsible for the somaclonal variations regenerated from calli. The development of resistance in vitro in callus cultures against the phytotoxins has been correlated with the secretion of extracellular proteins by the host in culture, which may be antifungal hydrolases or pathogenesis-related proteins (Jayasankar and Litz, 1998). The callus tissue offers opportunities for a variety of selection, including tolerance to phytotoxins and may allow recovery of desired variants, related to recessive or dominant genetic changes.
Successive screening on the higher levels of stress may easily produce physiological adaptations. In order to avoid physiological adaptations, the resistant bulblets were transferred to the culture medium in the absence of pathogen culture filtrate and subcultured two times before resistant identification on 50% pathogen culture filtrate stress. Identification of the somaclones on 50% culture filtrate stress showed that the resistance of selected lines is stable. To further confirm the stability, the regenerated bulblets were stored at 2°C for further growth and development and screened for resistance under glasshouse conditions. Out of 60 selected bulbs, 33 plants showed resistance (group 1) whereas most of the control bulbs died or showed morbidity when inoculated with Fusarium spore suspension (group 6), showing a strong correlation between in vitro selection and in vivo resistance. Similar results were reported by other workers showing the expression of resistance at the plant level in number of host-parasite interactions (Thakur et al., 2002; Kumar et al., 2008; Saxena et al., 2008). In the present investigation, an attempt was made to develop oriental hybrid lily ‘Casa Blanca’ resistant to Fusarium oxysporum f.sp. lilii through screening of callus cell lines against culture filtrate of the pathogen. Plants raised from one year-old bulbs were evaluated for resistance to fusarium wilt in glasshouse. The work is in progress to multiply bulbs and to evaluate these for the stability of character under field conditions. Further, molecular characterization of the resistant lines could be investigated to confirm the resistance at the DNA level.
CONCLUSION
In conclusion, this study was performed to develop cell lines of oriental hybrid lily resistant to fusarium wilt, which is a soil borne disease and difficult to control chemically. Resistant bulbs were developed using in vitro selection technique. The bulbs were successfully grown in the beds in glasshouse without any morphological variations. Therefore in vitro selection may become one of the main and effective ways of lily variation breeding with resistance to both biotic and abiotic stresses. In vitro selection for recovery of lily plants resistant to F. oxysporum toxins could be an alternative to genetic transformation for enhancing resistance to wilt pathogen in important cultivars of this crop.
REFERENCES
- Amini, J., 2009. Physiological Race of Fusarium oxysporum F. sp. Lycopersici in Kurdistan province of iran and reaction of some tomato cultivars to race 1 of pathogen. Plant Pathol. J., 8: 68-73.
CrossRef - Azadi, P. and M. Khosh-Khui, 2007. Micropropagation of Lilium ledebourii (Baker) Boiss as affected by plant growth regulator, sucrose concentration, harvesting season and cold treatments. Electron. J. Biotechnol., Vol. 10.
CrossRefDirect Link - Bacchettal, L., P.C. Remotti, C. Bernardini and F. Saccardof, 2003. Adventitious shoot regeneration from leaf explants and stem nodes of Lilium. Plant Cell Tissue Organ Culture, 74: 37-44.
CrossRefDirect Link - Benschop, M., R. Kamenetsky, M. Le Nard, H. Okubo and A. De Hertogh, 2010. The Global Flower Bulb Industry: Production, Utilization, Research. In: Horticultural Reviews, Volume 36, Janick, J. (Ed.). John Wiley and Sons Inc., Hoboken, NJ., USA., pp: 1-110.
CrossRefDirect Link - Ding, D., C.P. Lu, J.H. Wang, G.F. Cui, X.W. Wu and Y.P. Zhang, 2011. In vitro selection for lily clonal lines with Fusarium oxysporum f. sp. lilii toxin. J. Hunan Agric. Univ., 37: 34-38.
CrossRefDirect Link - Garibaldi, A. and M. Gullino, 1987. Fusarium wilt of carnation: Present situation, problems and perspectives (°). Acta Hortic., 216: 45-52.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Hammond, J., H.T. Hsu, Q. Huang, R. Jordan, K. Kamo and M. Pooler, 2006. Transgenic approaches to disease resistance in ornamental crops. J. Crop Improv., 17: 155-220.
CrossRef - Hansen, D.L., C. Lambertini, A. Jampeetong and H. Brix, 2007. Clone-specific differences in Phragmites australis: Effects of ploidy level and geographic origin. Aquat. Bot., 86: 269-279.
CrossRefDirect Link - Hartman, C.L., T.J. McCoy and T.R. Knous, 1984. Selection of alfalfa (Medicago sativa) cell lines and regeneration of plants resistant to the toxin(s) produced by Fusarium oxysporium f. sp. medicaginis. Plant Sci. Lett., 34: 183-194.
CrossRef - Khawar, K.M., S. Cocu., I. Parmaksiz, E.O. Sarihan and S. Ozcan, 2005. Mass proliferation of Madonna lily (Lilium candidum L.) under in vitro conditions. Pak. J. Bot., 37: 243-248.
Direct Link - Kintzios, S., A. Koliooulos, E. Karyoti, J. Drossopoulos, C.D. Holevas, A. Griggoriu and C.G. Panagopoulos, 1996. In vitro reaction of sunflower (Helianthus annuus L.) to the toxin(s) produced by Alternaria alternata, the casual agent of brown leaf spot. J. Phytopathol., 144: 465-470.
CrossRefDirect Link - Kumar, S., S. Kumar, S.P. Negi and J.K. Kanwar, 2008. In vitro selection and regeneration of chrysanthemum (Dendranthema grandiflorum Tzelev) plants resistant to culture filtrate of Septoria obesa Syd. In vitro Cell. Dev. Biol. Plant, 44: 474-479.
CrossRef - Lim, J.H., S.T. Cho, H.K. Rhee, H.R. Cho and H.Y. Joung, 2005. Level of resistant to Fusarium oxysporum f.sp. lilii (Fol 4 and 11) using endogenous antifungal substances extracted from bulbs and/or roots in Lilium genus. Acta Hort., 673: 645-652.
Direct Link - Liu, S., H. Wang, J. Zhang, B.D.L. Fitt and Z. Xu et al., 2005. In vitro mutation and selection of doubled-haploid Brassica napus lines with improved resistance to Sclerotinia sclerotiorum. Plant Cell Rep., 24: 133-144.
CrossRef - Loffler, H.J.M., H. Meijer, Th.P. Straathof and J.M. van Tuyl, 1996. Segregation of Fusarium resistance in an interspecific cross between Lilium longiflorum and Lilium dauricum. Acta Hortic., 414: 203-208.
Direct Link - Loffler, H.J.M. and J.R. Mouris, 1992. Fusaric acid: Phytotoxicity and in vitro production by Fusarium oxysporum f.sp. lilii, the causal agent of basal rot in lilies. Eur. J. Plant Pathol., 98: 107-115.
CrossRef - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nasir, I.A. and S. Riazuddin, 2008. In vitro selection for Fusarium wilt resistance in gladiolus. J. Integr. Plant Biol., 50: 601-612.
CrossRef - Nhut, D.T., 2003. The control of in vitro direct main stem formation of Lilium longiflorum derived from receptacle culture and rapid propagation by using in vitro stem node. Plant Growth Regul., 40: 179-184.
Direct Link - Prados-Ligero, A.M., M.J. BasallotE-Ureba and J.M. Maria-Vara, 2008. First report of Fusarium oxysporum f. sp. lilii and F. proliferatum affecting Lilium crops in Spain. Trop. Plant Pathol., 33: 235-236.
CrossRefDirect Link - Saifullah, K., N. Sheeba, R. Mariam, K. Naheed, N. Asma and S. Bushra, 2010. Cultivation of lilies (Lilium regale) for commercialization in Pakistan. Pak. J. Bot., 42: 1103-1113.
Direct Link - Sallam, N.M.A. and M.F. Abdel-Monaim, 2012. Influence of some agricultural practices on suppression of lentil wilt disease. Plant Pathol. J., 11: 32-37.
CrossRefDirect Link - Sesto, F., V. Grimaldi and A.M. Pennisi, 1990. Sensitivity of different citrus and non-citrus species protoplasts towards malsecco toxins. Adv. Hort. Sci., 4: 97-102.
Direct Link - Straathof, T.P. and J.M. van Tuyl, 1994. Genetic variation in resistance to Fusarium oxysporum f. sp. lilii in genus Lilium. Ann. Applied Biol., 125: 61-72.
CrossRef - Thakur, M., D. Sharma and S. Sharma, 2002. In vitro selection and regeneration of carnation (Dianthus caryophyllus L.) plants resistant to culture filtrate of Fusarium oxysporum f. sp. dianthi. Plant Cell Rep., 20: 825-828.
CrossRefDirect Link - Van Tuyl, J.M. and H.C.M. van Holsteijn, 1996. Lily breeding research in the Netherlands. Acta Hort., 414: 35-45.
Direct Link - Xu, L.F., M.F. Wang and L. Dong, 2009. Plant regeneration from in vitro cultured leaves of Lanzhou lily (Lilium davidii var. Unicolor). Sci. Hortic., 119: 458-461.
CrossRef - Zhang, L.Q., Z.H. Cheng, M.A. Khan and Y.L. Zhou, 2012. In vitro selection of resistant mutant garlic lines by using crude pathogen culture filtrate of Sclerotium cepivorum. Aust. Plant Pathol., 41: 211-217.
CrossRef - Svabova, L. and A. Lebeda, 2005. In vitro selection for improved plant resistance to toxin-producing pathogens. J. Phytopathol., 153: 52-64.
CrossRef