
W.M. Jurick II
Food Quality Laboratory, USDA-ARS, BARC-West, MD, Beltsville, United State of America
I. Vico
Institute of Phytomedicine, Faculty of Agriculture, University of Belgrade, Serbia
B.D. Whitaker
Food Quality Laboratory, USDA-ARS, BARC-West, MD, Beltsville, United State of America
V.L. Gaskins
Food Quality Laboratory, USDA-ARS, BARC-West, MD, Beltsville, United State of America
W.J. Janisiewicz
Appalachian Fruit Research Station, USDA-ARS, WV, Kearneysville, United State of America
Plant Pathology Journal
Year: 2012 | Volume: 11 | Issue: 1 | Page No.: 38-41







ABSTRACT
Cellulose is the most abundant form of carbon on the planet. Breakdown of cellulose microfibrils in the plant cell wall is a means by which microbes gain ingress into their respective hosts. Cellulose degradation is also important for global carbon recycling and is the primary substrate for production of biofuels. In this study, we developed a cellulase assay method that rivals the commonly used dinitrosalicylic acid (DNS) assay. It was shown that the 2-cyanoacetamide method is capable of detecting D-glucose in a linear fashion, can function in various buffers at pH ranging from 4.0 to 8.0 and is as sensitive as the DNS test at detecting fungal cellulase activity using carboxymethyl cellulose as a substrate. This method will be useful to others who desire to accurately and efficiently assay microbial cellulase activity without the use of phenol and other highly toxic and corrosive chemicals.
PDF Abstract XML References Citation
Received: November 30, 2011;
Accepted: March 26, 2012;
Published: June 27, 2012
How to cite this article
W.M. Jurick II, I. Vico, B.D. Whitaker, V.L. Gaskins and W.J. Janisiewicz, 2012. Application of the 2-Cyanoacetamide Method for SpectrophotometricAssay of Cellulase Enzyme Activity. Plant Pathology Journal, 11: 38-41.
DOI: 10.3923/ppj.2012.38.41
URL: https://scialert.net/abstract/?doi=ppj.2012.38.41
DOI: 10.3923/ppj.2012.38.41
URL: https://scialert.net/abstract/?doi=ppj.2012.38.41
INTRODUCTION
Cellulose is the most abundant carbohydrate found in plants that provides structural integrity to their cell walls (Bhat and Bhat, 1997). It is composed of a linear polymer of D-glucose linked by β-1,4 bonds which can be degraded by the cellulase enzyme complex (Karnchanatat et al., 2008). The enzyme complex is composed of endoglucanases which cleave internal portions of the cellulose chain followed by cellobiohydrolases and β-glucosidases which yield soluble cellodextrins and glucose (Wood, 1992; Youssef and Berekaa, 2009). Bacterial and fungal plant pathogens utilize cellulases to breach the plant cell wall for nutrient acquisition and colonization (Khan and Husaini, 2006; Walton, 1994).
Biological turnover of cellulose is important in carbon recycling and has received much attention for biofuel applications (Kilic and Saricicek, 2010; Milala et al., 2009; Roslan et al., 2011). Cellulases are used in the textile industry for improving softness of fabrics and are also used in the paper industry (Karnchanatat et al., 2008; Khajavi et al., 2007). Endoglucanases, a specific component of the cellulase enzyme complex, are utilized in beer filtration, oil extraction, increasing the yield of fruit juices and improving the nutritional value of baked goods and animal feed (Bhat, 2000; Devanathan et al., 2007; Pinelo et al., 2007; Zakaria et al., 2010).
Spectrophotometric methods previously used to detect cellulase activity were based on the reducing sugar method described by Miller (1959) that requires dinitrosalicylic acid (DNS), phenol and sodium hydroxide, which are known carcinogens and highly corrosive. The main objective of this study was to adapt a spectrophotometric method that is accurate, rapid and non-toxic for the detection of cellulase activity using 2-cyanoacetamide. We also sought to determine the effects of different buffer systems and pH on the detection of D-glucose and compared the modified 2-cyanoacetamide method with the DNS assay using a commercial cellulase preparation from Aspergillus niger.
MATERIALS AND METHODS
Detection of D-glucose using 2-cyanoacetamide was carried out using the method previously described for D-galacturonic acid by Gross (1982). D-glucose standards were dissolved in 50 mM sodium acetate buffer pH 5.0 at 5, 25, 50, 100, 200, 400 and 800 mg mL-1. Each concentration was measured in triplicate along with a corresponding blank. The blank value was subtracted to adjust for background. One hundred microliters of a glucose solution was added to 1 mL of 100 mM borate pH 9.0 and 200 μL of 1% 2-cyanoacetamide. The solution was mixed by vortexing and then boiled for 10 min. Samples were cooled to room temperature and absorbance at 276 nm was determined using a Shimadzu spectrophotometer model UV-2501PC (Shimadzu Scientific, North America). To test the effect of pH and buffer composition on the detection of D-glucose using 2-cyanoacetamide, standard solutions of D-glucose were dissolved in 50 mM MES (2-(N-morpholino) ethanesulfonic acid), tris (2-Amino-2-hydroxymethyl-propane-1,3-diol) or succinate. Each buffer solution was adjusted to its pKa and therefore succinate was adjusted to pH 4.0, MES at pH 6.0 and Tris at pH 8.0. The standards were prepared in triplicate and the readings were adjusted with a corresponding blank.
Detection of D-glucose using the DNS (dinitrosalicylic acid) method was carried out as previously described by Miller (1959). D-glucose standards were prepared in 50 mM sodium acetate buffer pH 5.0 at 5, 25, 50, 100, 200, 400 and 800 mg mL-1 and the reaction was carried out with a reagent composed of: 141.6 mL water, 1.06 g of DNS and 1.89 g sodium hydroxide were mixed and then 30.6 g sodium potassium tartrate, 0.76 mL phenol solution and 0.83 g of sodium metabisulfite were added. One hundred microliters of the sample was mixed with the DNS reagent and was boiled for 5 min. Samples were cooled to room temperature and absorbance at 540 nm was determined using a Shimadzu spectrophotometer model UV-2501PC (Shimadzu Scientific, North America).
Determination of cellulase enzyme activity using CMC as a substrate was carried out using both the 2-cyanoacetamide and the DNS methods for comparison. A commercial source of cellulase enzyme was prepared from A. niger (Sigma Aldrich, St. Louis, MO).
The cellulase preparation was diluted 1:500 in water and 500 μL of this solution was added to 500 μL of 1% carboxymethyl cellulose dissolved in 50 mM sodium acetate buffer pH 5.0. Samples were incubated in a water bath at 37°C for 0, 20, 40, 60 and 120 min. There were three samples and a blank for each time point. Samples were then treated according to the protocols for both methods outlined above concerning the termination of reactions and product measurement.
RESULTS AND DISCUSSION
It was shown that the 2-cyanoacetamide method is as linear as the DNS method with respect to the detection of D-glucose in solution (Fig. 1a, b). This is expected since the 2-cyanoacetamide method has previously been shown to detect other reducing sugars (i.e., galacturonic acid) which form UV-absorbing products that fluoresce (Gross, 1982). The 2-cyanoacetamide method does not use phenol, dinitrosalicylic acid or NaOH and is equally sensitive as the DNS method at detecting D-glucose (Miller, 1959). Therefore, the modified 2-cyanoacetamide procedure provides an advantage over the DNS method in that it does not use potential carcinogens and corrosive chemicals and does not require a fume hood to conduct the assay.
To determine the effect of pH and buffer composition on the detection of D-glucose, we constructed standard curves in three different buffers (MES, succinate and TRIS) and adjusted them to pH’s ranging from 4 to 8. These specific buffers were chosen because they are commonly used to assay enzyme activity including cellulase (Karnchanatat et al., 2008). All buffer systems tested yielded similar results as indicated by R2 values of 0.95 that were obtained from linear regressions from each data set (Fig. 2a-c).
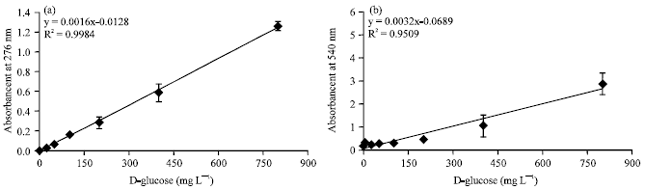 | |
| Fig. 1(a-b): | Relationship between D-glucose concentration (5-800 mg L-1) in 50 mM sodium acetate buffer pH 5.0 and absorbance using (a) 2-cyanoacetamide and (b) DNS methods, Each point on the line represents absorbance of samples assayed in triplicate which were adjusted for background. The error bars indicate the standard deviation |
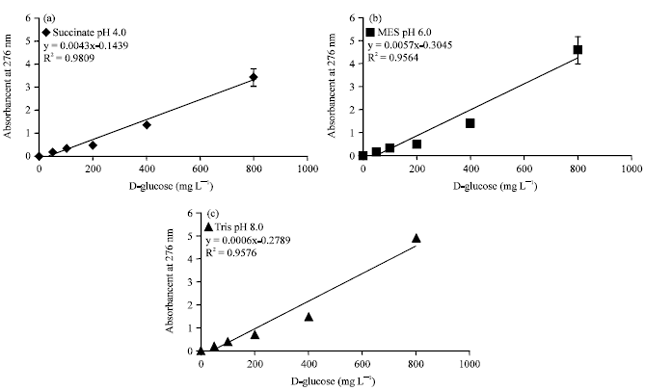 | |
| Fig. 2(a-c): | Relationship between D-glucose concentration (5-800 mg L-1) in different buffers adjusted from pH 4.0 to 8.0 and absorbance at 276 nm using the 2-cyanoacetamide method, (a) D-glucose in 50 mM succinate pH 4.0, (b) D-glucose in 50 mM MES pH 6.0 and (c) D-glucose in 50 mM succinate pH 8.0, Each point on the line represents absorbance of samples assayed in triplicate that were adjusted for background, The error bars indicate the standard deviation |
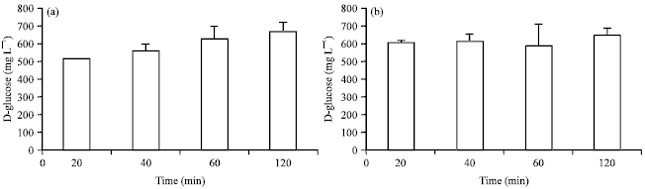 | |
| Fig. 3(a-b): | Comparison of cellulase activity Aspergillus niger cellulase using carboxymethyl cellulose as a substrate at different time points. Detection of reducing sugars using (a) The 2-cyanoacetamide method and (b) The DNS method, Bars represent the mean values assayed in triplicate that were adjusted for background, The error bars indicate the standard deviation |
Both methods (2-cyanoacetamide and DNS) were capable of detecting cellulase activity of a commercial source of enzyme obtained from Aspergillus niger using carboxymethyl cellulose (CMC) as a substrate (Fig. 3a, b). CMC is widely used as a substrate to assay cellulase activity (Arunachalam et al., 2010; Hurst et al., 1977; Parry et al., 2002; Sharrock, 1988). Both methods were capable of detecting D-glucose liberated from CMC over time. The 2-cyanoacetamide assay detected a linear increase in D-glucose concentration with increasing incubation time whereas D-glucose detection using the DNS assay was variable with prolonged incubation.
The use of the modified 2-cyanoacetamide method to assay cellulase activity, presented in this technical note, has several advantages over the DNS method. The 2-cyanoacetamide method can accurately detect D-glucose in a linear fashion in various buffer systems over a wide pH range. The modified 2-cyanoacetamide method can also accurately assay cellulase activity using CMC as a substrate with very low sample variability. Moreover, this assay can be conducted without the use of a fume hood because the 2-cyanoacetamide method does not involve phenol, sodium hydroxide or dinitrosalicylic acid all of which also require special handling for disposal. A final advantage of the 2-cyanoacetamide method is that one can easily detect other enzyme activities (i.e., polygalacturonase) by simply changing the substrate in the assay buffer (Gross, 1982).
REFERENCES
- Arunachalam, R., E.G. Wesely, J. George and G. Annadurai, 2010. Novel approaches for identification of Streptomyces noboritoensis TBG-V20 with cellulase production. Curr. Res. Bacteriol., 3: 15-26.
CrossRefDirect Link - Bhat, M.K., 2000. Cellulases and related enzymes in biotechnology. Biotechnol. Adv., 18: 355-383.
CrossRefPubMedDirect Link - Bhat, M.K. and S. Bhat, 1997. Cellulose degrading enzymes and their potential industrial applications. Biotechnol. Adv., 15: 583-620.
CrossRefDirect Link - Devanathan, G., A. Shanmugam, T. Balasubramanian and S. Manivannan, 2007. Celluase production by Aspergillus niger isolated from coastal mangrove debris. Trends Applied Sci. Res., 2: 23-27.
CrossRefDirect Link - Hurst, P.L., J. Nielsen, P.A. Sullivan and M.G. Shepherd, 1977. Purification and properties of a cellulase from Aspergillus niger. Biochem. J., 165: 33-41.
PubMed - Karnchanatat, A., A. Petsom, P. Sangvanich, J. Piapukiew and A.J.S. Whally et al., 2008. A novel thermostable endoglucanase from the wood-decaying fungus Daldinia eschscholzii (Ehrenb.:Fr.) Rehm. Enzyme Microb. Technol., 42: 404-413.
CrossRefDirect Link - Khajavi, R., B. Novinrad and A. Kiumarsi, 2007. The effect of ultrasonic on the denim fabric worn out process. Pak. J. Biol. Sci., 10: 645-648.
CrossRefPubMedDirect Link - Khan, F.A.B.A. and A.A.S.A. Husaini, 2006. Enhancing α-amylase and cellulase in vivo enzyme expressions on sago pith residue using Bacilllus amyloliquefaciens UMAS 1002. Biotechnology, 5: 391-403.
CrossRefDirect Link - Kilic, U. and B.Z. Saricicek 2010. The effects of different silage additives on in vitro gas production, digestibility and energy values of sugar beet pulp silage. Asian J. Anim. Vet. Adv., 5: 566-574.
CrossRefDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Parry, N.J., D.E. Beever, E. Owen, W. Nerinckx, M. Claeyssens, J. Van Beeumen and M.K. Bhat, 2002. Biochemical characterization and mode of action of a thermostable endoglucanase purified from Thermoascus aurantiacus. Arch. Biochem. Biophys., 404: 243-253.
CrossRef - Pinelo, M., A.G. Tress, M. Pedersen, A. Arnous and A.S. Meyer, 2007. Effect of cellulases, solvent type and particle size distribution on the extraction of chlorogenic acid and other phenols from spent coffee grounds. Am. J. Food Technol., 2: 641-651.
CrossRefDirect Link - Roslan, A.M., P.L. Yee, U.K.M. Shah, S.A. Aziz and M.A. Hassan, 2011. Production of bioethanol from rice straw using cellulase by local Aspergillus sp. Int. J. Agric. Res., 6: 188-193.
CrossRefDirect Link - Youssef, G.A. and M.M. Berekaa, 2009. Improved production of endoglucanase enzyme by Aspergillus terreus: Application of plackett burman design for optimization of process parameters. Biotechnology, 8: 212-219.
CrossRefDirect Link - Zakaria, H.A.H., M.A.R. Jalal and M.A. Abu Ishmais, 2010. The influence of supplemental multi-enzyme feed additive on the performance, carcass characteristics and meat quality traits of broiler chickens. Int. J. Poult. Sci., 9: 126-133.
CrossRefDirect Link - Milala, M.A., B.B. Shehu, H. Zanna and V.O. Omosioda, 2009. Degradation of agro-waste by cellulase from Aspergillus candidus. Asian J. Biotechnol., 1: 51-56.
CrossRefDirect Link