
A. Pandey
Molecular Plant Pathology Laboratory, Central Institute for Subtropical Horticulture, Lucknow-227 107, U.P., India
B.K. Pandey
Molecular Plant Pathology Laboratory, Central Institute for Subtropical Horticulture, Lucknow-227 107, U.P., India
M. Muthukumar
Molecular Plant Pathology Laboratory, Central Institute for Subtropical Horticulture, Lucknow-227 107, U.P., India
L.P. Yadava
Molecular Plant Pathology Laboratory, Central Institute for Subtropical Horticulture, Lucknow-227 107, U.P., India
U.K. Chauhan
Center for Biotechnology, School of Environmental Biology, A.P.S. University, Rewa- 486 003, M.P., India
Plant Pathology Journal
Year: 2012 | Volume: 11 | Issue: 1 | Page No.: 18-24







ABSTRACT
Anthracnose, caused by Colletotrichum gloeosporioides, is a serious postharvest disease of mango. The histopathological studies on anatomy of naturally infected by Colletotrichum gloeosporioides and artificially inoculated leaves and healthy leaves were performed to understand the infection process of anthracnose at various intervals after inoculation. Germination and penetration processes of the pathogen within the whole leaf were observed. The first evidence of penetration into the whole leaf was observed 48 h after invasion. It also revealed that mycelia were prominent after 120 h after invasion by the fungus (C. gloeosporioides). Subcuticular infection by hyphae was present in transverse leaf sections (T.S.) of the diseased sample after 72 h. Also, both inter and intra-cellular hyphal invasion were observed after 72 h. Mesophyll cells were highly affected by fungal invasion and rapidly collapsed. Swelling of epidermal cell walls was also observed. After 96 h almost all the cells became necrotized (Nc). Necrotized mycelial mats (M) of C. gloeosporioides was observed after 120 h and all the invaded cells became necrotized (Nc) forming a spot which eventually the cells ruptured leaving a shot hole symptom. All these observations pertained to the cells of mesophyll tissue indicating that these are the regions of fungal invasion and host tissue damage resulting in the disease symptoms. Naturally infected and artificially inoculated (in vitro) presented no significant differences suggesting that the pathogen invasion and symptom development process is similar in both the conditions.
PDF Abstract XML References Citation
Received: December 21, 2011;
Accepted: March 06, 2012;
Published: June 27, 2012
How to cite this article
A. Pandey, B.K. Pandey, M. Muthukumar, L.P. Yadava and U.K. Chauhan, 2012. Histopathological Study of Infection Process of Colletotrichum gloeosporioides Penz and Sacc. on Mangifera indica L. Plant Pathology Journal, 11: 18-24.
DOI: 10.3923/ppj.2012.18.24
URL: https://scialert.net/abstract/?doi=ppj.2012.18.24
DOI: 10.3923/ppj.2012.18.24
URL: https://scialert.net/abstract/?doi=ppj.2012.18.24
INTRODUCTION
Mango (Mangifera indica L.) is affected by a number of diseases of which anthracnose is one of the most common especially in India resulting in substantial loss. Many variety of fruit crops including almond, apple, mango (Freeman et al., 1998), avocado (Sanders and Korsten, 2003), cassava (Fokunang et al., 2000a, 2003), Arabica coffee (Agwanda et al., 1997), guava (Hasna et al., 2000; Ansari et al., 2000; Amusa et al., 2005a, 2006) banana (Haque et al., 2003; Nuangmek et al., 2008; Taechowisan et al., 2009), papaya (Al Eryani-Raqeeb et al., 2009) strawberry (Martinez-Culebras et al., 2000; Xiao et al., 2004) and yam (Amusa et al., 2005b).
For validation an organism as an etiologic agent it is must to record histopathological studies of hyphal elements in tissue, as for other mycoses (Sutton, 1999). In host specificity test, C. gloeosporioides was found to be highly pathogenic on the fruits of mango. However, other fruit crops i.e., grape (Leu and Chang, 1985), avocado (Coates et al., 1993), strawberry (Xiao et al., 2004) and olive (Talhinhas et al., 2011) are also affected by this pathogen. Meanwhile, very limited information exists on the infection process of C. gloeosporioides (Makowski, 1987). Morin et al. (1996) showed that infection of C. gloeosporioides f. sp. malvae involved intracellular vesicles, large-diameter primary hyphae that constricted at transcellular penetration sites and secondary hyphae. In susceptible host development of primary hyphae and production of secondary hyphae was extensive. The colonization was stopped by a oversensitive reaction of cell contiguous to preliminary infection site in the moderately resistant hosts M. neglecta and A. theophrasti.
The Scanning Electron Microscopy (SEM) was found to be an excellent tool for presenting details of plant structure (Troughton and Donaldson, 1972). With this basic information, the present study was initiated to comprehend the infection process of the pathogen, the anatomical changes that took place after invasion, pathogenesis and finally how the disease developed.
MATERIALS AND METHODS
Plant material: To observe the anatomical changes in leaf during host pathogen interaction tissue samples of leaf were collected from naturally and artificially infected plants for expression study analysis. Leaf samples from healthy mango plant (height 100-150 cm) grown under green house conditions were also taken as control in the experiment.
Preparation of spore suspension of C. gloeosporioides: C. gloeosporioides was grown on potato dextrose agar (PDA) medium and incubated at 25-30°C for seven days. A 5 mm disc was used for making spore suspension in different concentrations (2.6x105, 0.16x105, 0.032x105, 0.0025x105 and 0.00015x105 mL-1) by serial dilution with sterile distilled water.
In vitro inoculation and pathogenicity assay: Spore suspension of fungal growth were inserted into artificially made wounds by pinching using sterile needle and then leaves were surface-sterilized with 70% ethanol. Control was also maintained. Samples were observed at 48, 72, 96 and 120 h after inoculation. Control plant was similarly sampled with distilled water. The leaves were covered with polythene bags and incubated at 28±2°C to ensure high humidity for establishing favorable conditions for conidial germination and infection. After 5 days of incubation, pathogen was reisolated and its identity was confirmed. Wound inoculation method was used to test the pathogenicity on Dashehari mango leaves as suggested by Bhuvaneswari and Rao (2001).
Fixation of tissues: Tissue blocks from healthy or infected mango leaves were washed in distilled water and dried between folds of sterilized paper towels and processed. The tissues were cut into small portions (5-10 mm long), dipped in (FAA) formaldehyde: acetic acid: absolute alcohol (5:5:90 mL) (Sass, 1940). The samples were left in the above solution till they were processed further.
Dehydration, infiltration and embedding: The dehydration of specimens were processed by using chemicals i.e., alcohol 30, 50, 70, 80 and 95%, absolute alcohol, alcohol 75% + xylene 25%, alcohol 50% + xylene 50%, alcohol 25%+xylene 75% and pure xylene consequently each for 30 minutes with the help of automatic tissue processor (York, New Delhi). There after to prepare blocks, tissues were embedded in melted paraffin wax at 54 to 56°C for 4-8 h in order to completely replace the xylene with paraffin wax (Berlyn et al., 1976).
Staining and mounting: A thin section of 10 micron thickness was cut with the help of a microtome (Microm, Germany). At least 20 slides were prepared for each sample. The sections were stained in 0.1% aqueous toluidine blue and were mounted in DPX-mount after bringing them to xylene through alcohol-xylene series as described by Jensen (1962). The healthy leaf was also examined in the study as control along with diseased samples which were assessed for anatomical changes as per the technique described by Pandey et al. (1987).
Light microscopy observation: Light microscopic observation was made at 48, 72, 96 and 120 h after inoculation on 5, 10, 40 and 100X magnification, respectively following the method of Begum et al. (2007).
RESULTS
The histopathological studies were carried out to investigate the behavior and interaction of C. gloeosporioides on mango leaves under natural infection and in vitro conditions (Fig. 1). Typical features of shape and size of the tissues was shown in transverse section (T.S.) of dorsiventral healthy leaf (Fig. 2). The collaterally closed vascular bundles were found scattered and surrounded by Parenchymatic cells (P) which delimited by fibres. Parenchymatous cells spread above and below the vascular bundles. Transverse sections (T.S.) of the tissues showed no material inside vascular bundles (Vb), [xylem (Xy) and phloem (Ph)]. The intercellular spaces and the interfaces of contact between neighbouring cells were completely free.
Transverse sections of tissues naturally infected by C. gloeosporioides indicated cell plasmolysis, disorganization and dark brown discoluration of parenchymatous cells (P) surrounding vascular bundles (Fig. 2). Parenchymatic cells (P) were filled with brown to dark mycelia. In advanced stages, the pathogen spread to adjoining invaded tissues was observed and fibers causing complete breakdown. Mycelia were also detected in and within the metaxylem (Xy) elements. Hyphae extensively ramified in the invaded intercellular spaces, surrounded cells and xylem units. Massive dissolution of parenchymatous cells (P) and frequently xylem or fibrous tissues were also observed.
 | |
| Fig. 1(a-e): | Infection process on leaves, (a) Inoculation on the first day, (b) Initiation of symptoms on leaf after 48 h of inoculation, (c) Small spots symptom after 72 h of inoculation, (d) Enlarged spot symptom on leaf after 96 h of inoculation and (e) Anthracnose symptoms after 120 h of inoculation (Arrows represent the position/location of the symptoms) |
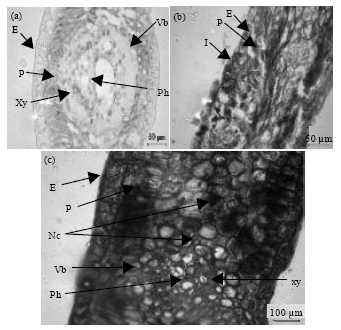 | |
| Fig. 2(a-c): | Transverse section (T.S.) of healthy and naturally infected mango leaves by C. gloeosporioides; (a) Regular and typical structure of shape and size of the epidermis (E), parenchyma (P), vascular bundle (Vb), xylem (Xy) and phloem (Ph) in healthy leaf (b) Initiation of anthracnose symptom in epidermis (E), browning and darkening due to necrosis in naturally infected mango leaf (c) Typical symptoms of anthracnose such as dark tissues and necrotized cells (Nc) and changes in shape and size of epidermis (E), parenchyma (P), vascular bundle (Vb), xylem (Xy) and phloem (Ph) in the naturally infected mango leaf |
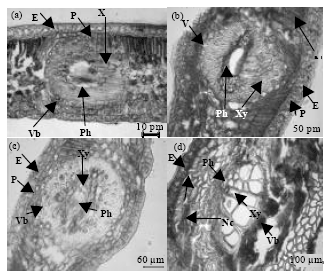 | |
| Fig. 3(a-d): | Transverse section (T.S.) of mango leaves of artificially inoculated control (H2O) and spore suspension of C. gloeosporioides observed after various time intervals after inoculation, (a) Typical T.S. of healthy leaf and the shape and size of the epidermis (E), parenchyma (P), vascular bundle (Vb), xylem (Xy) and phloem (Ph) after 48 h of inoculation, (b) Initiation of anthracnose symptoms and some cells are necrotize (Nc) after 48 h of inoculation, (c) T.S. of healthy leaf after 72 h of inoculations showing that all the structures and tissues are intact, regular and the cells have grown in size and volume, (d) Subcuticular infection and infection of inter and intra-cellular spaces observed after 72 h of inoculation and changes in the epidermis (E) and outer cells of Vb during necrosis observed in infected leaf |
No visible changes in the histological structure were noticed in the other tissues or vascular bundle elements. These results were positively correlated and could interpret with the degree of virulence of each pathogenic fungus.
The in vitro inoculated tissues with spore suspension of C. gloeosporioides exerted the same deleterious effects (Fig. 3, 4). Similar results were also recorded in the tissues infected with spore suspension of C. gloeosporioides at 96 and 120 h after inoculation. Mycelia were prominent after 120 h after invasion by the fungus, C. gloeosporioides. But the first evidence of penetration into the whole leaf was observed 48 h after invasion. Subcuticular infection by hyphae was present in transverse leaf sections (T.S.) of the diseased sample after 72 h. Also, both inter and intra-cellular hyphal invasion were observed after 72 h. Mesophyll cells were highly affected by fungal invasion and rapidly collapsed. Swelling of Epidermal cell (E) walls was also observed. After 96 h almost all the cells became necrotized (Nc). Necrotized mycelial mats (M) of C. gloeosporioides was observed after 120 h and all the invaded cells became necrotized (Nc) forming a spot which eventually the cells ruptured leaving a shot hole symptom (Fig. 4). All these observations pertained to the cells of mesophyll tissue (parenchyma cells) indicating that these are the regions of fungal invasion and host tissue damage resulting in the disease symptoms.
DISCUSSION
Cells, of host plant affected by anthracnose, exhibited unnatural tissues that turn brown to dark (Rivera-Vargas et al., 2006) and filled with pentosan like substances (Klotz and Fawcett, 1932). On the contrary, the fungus T. paradoxa/C. gloeosporioides sporulated in masses in parenchymatic tissues and metaxylem vessels. Besides, sections exerted mycelium in fibrous sclerenchyma cells surrounding the vascular elements. Spores, gum like substances and tylosis that formed in the infected tissues might bring about disfunction of the xylem elements and might interfere seriously with the transpiration stream and produce a permanent wilting of the pinnae several feet beyond the region invaded by the fungus (Lopez-Llorca and Orts, 1994).
C. gloeosporioides might produce extracellular cutinase to weaken the host barrier and thus assist infection peg to penetrate through the cuticle (Kolattukudy, 1985). The penetration procedure of C. gloeosporioides on mango leaf is similar with that of C. gloeosporioides on avocado (Binyamini and Schiffmann-Nadel, 1972), Robinson tangerine (Brown, 1977), papaya (Chau and Alvarez, 1983), grape (Leu and Chang, 1985), C. capsici on red pepper (Adikaram et al., 1983) and C. gloeosporioides f. sp. manihotis in cassava (Fokunang et al., 2000b).
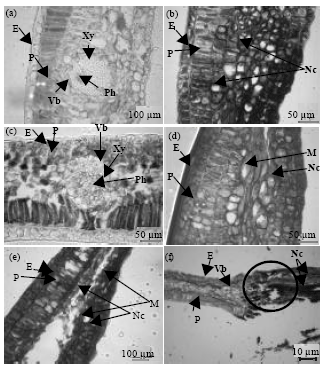 | |
| Fig. 4(a-f): | Transverse section (T.S.) of mango leaves in vitro inoculated control and spore suspension of C. gloeosporioides observed after various time intervals after inoculation, (a) Healthy leaf after 96 h of inoculation, (b) Infected leaf after 96 h and some cells of epidermis (E) and parenchyma cells (P) are found necrotized (Nc), (c) Healthy leaf after 120 h of inoculation, (d) Infected leaf showing that most of all cells of epidermis (E) and parenchyma (P) are necrotized (Nc) and infected with some mycelial part (M) of C. gloeosporioides after 120 h of inoculation, (e) Approximately all the cells of epidermis (E) and parenchyma (P) are found necrotized (Nc) and some M are also observed, (f) Shot hole symptom and rupturing of the tissues marked by ring |
This mode of infection was in accordance with other anthracnose (Manandhar et al., 1985).
Epidemiology and histopathology studies showed the presence of the pathogens on vegetative organs year-round, particularly on olive leaves and branches and on weeds. These represent inoculums reservoirs where secondary conidiation occurs and conidia are then dispersed by spring rains reaching flowers and young fruits or by autumn rains reaching pre-mature fruits. Unripe fruits were colonized without showing symptoms up to penetration of the cuticle, but further colonization and symptom production was completed only as fruits matured (Talhinhas et al., 2011).
In all interactions studied prepenetration processes were generally similar, leading to the formation of predominantly sessile melanized appressoria. Penetration occurred only from melanized appressoria, and in both compatible and incompatible interactions (Trevorrow et al., 1988). C. gloeosporioides sp. penetrated plant cuticles directly and produced infection structures within epidermal cells. The subcuticular hyphae seen in C. gloeosporioides infecting other plants were not observed in this study (Brown, 1977; Chau and Alvarez, 1983).
Similarly, observed formation of pinpoint lesions (0.5 to 1 mm diam.) on stems or explants of A. virginica within 48 h after inoculation with C. gloeosporioides. Infection resulted in the formation of stem lesions, 2 to 3 cm long, which encircled the stem within 6 to 8 days after inoculation. Intracellular mycelium grew within the cortex, cambium, xylem and pith ray tissues. Death of A. virginica seedlings was caused by collapse of infected stem tissues. Coalescence of lesions enhanced girdling of stems and hastened death. The fungus sporulated abundantly on lesion surfaces (TeBeest et al., 1978). Necrotic lesions produced orange to salmon conidial masses on acervuli 7 days after inoculation (Fokunang et al., 2000c; Rivera-Vargas et al., 2006).
Microscopic examination of sections taken from infected tissues indicates the ability of pathogenic fungi to invade the different tissues causing changes in cells structure and caused plasmolysis and discoloration of parenchymatous cells. Hyphae were seen to grow intra and intercellular very often. Spores were found to form (or transferred) within parenchyma cells and xylem vessels. Both intracellular hemibiotrophic and subcuticular intramural Colletotrichum species avoid triggering resistance responses during the symptomless phase.
ACKNOWLEDGMENT
Authors are thankful to Director of Central Institute for Subtropical Horticulture, Lucknow for providing necessary research facilities and research grant in the form of ICAR-SRF.
REFERENCES
- Adikaram, N.K.B., A.E. Brown and T.R. Swinburne, 1983. Observations on infection of Capsicum annuum fruit by Glomerella cingulata and Colletotrichum capsici. Trans. Br. Mycol. Soc., 80: 395-401.
Direct Link - Agwanda, C.O., P. Lashermes, P. Trouslot, M.C. Combes and A. Charrier, 1997. Identification of RAPD markers for resistance to coffee berry disease Colletotrichum kahawae in arabica coffee. Euphytica, 97: 241-248.
Direct Link - Amusa, N.A., O.A. Ashaye, M.O. Oladapo and M.O. Oni, 2005. Guava fruit anthracnose and the effects on its nutritional and market values in Ibadan, Nigeria. World J. Agric. Sci., 1: 169-172.
Direct Link - Amusa, N.A., A.A. Adegbite and M.O. Oladapo, 2005. Investigations into the role of weeds, soil and plant debris in the epidemiology of foliar fungal diseases of yam in Western Nigeria. Int. J. Bot., 1: 111-115.
CrossRefDirect Link - Amusa, N.A., O.A. Ashaye, J. Amadi and O. Oladapo, 2006. Guava fruit anthracnose and the effects on its nutritional and market values in Ibadan, Nigeria. J. Applied Sci., 6: 539-542.
CrossRefDirect Link - Ansari, T.H., T. Yoshida and M.B Meah, 2000. An integrated approach to control anthracnose of guava (Psidium guajava). Pak. J. Biol. Sci., 3: 791-794.
CrossRefDirect Link - Begum, M.M., M. Sariah, M.A. Zainal Abidin, A.B. Puteh and M.A. Rahman, 2007. Histopathological studies on soybean seeds infected by Fusarium oxysporum f. sp. glycines and screening of potential biocontrol agents. Res. J. Microbiol., 2: 900-909.
CrossRefDirect Link - Binyamini, N. and M. Schiffmann-Nadel, 1972. Latent infection in avocado fruit due to Colletotrichum gloeosporioides. Phytopathology, 62: 592-594.
Direct Link - Chau, K.F. and A.M. Alvarez, 1983. A histological study of anthracnose on Carica papaya. Phytopathology, 73: 1113-1116.
Direct Link - Coates, L.M., L.F. Muirhead, J.A.G. Irwin and D.H. Gowanlock, 1993. Initial infection processes by Colletotrichum gloeosporioides on avocado fruit. Mycol. Res., 97: 1363-1370.
Direct Link - Fokunang, C.N., T. Ikotun, A.G.O. Dixon, C.N. Akem, E.A. Tembe and E.N. Nukenine, 2000. Efficacy of antimicrobial plant crude extracts on the growth of Colletotrichum gloeosporioides f. sp. manihotis. Pak. J. Biol. Sci., 3: 928-932.
CrossRefDirect Link - Fokunang, C.N., T. Ikotun, C.N. Akem, A.G.O. Dixon, E.A. Tembe and P. Koona, 2000. Investigation of inoculum threshold and latent infection in Colletotrichum gloeosporiodes f.sp. manihotis, in Cassava cultivars. Pak. J. Biol. Sci., 3: 713-716.
CrossRefDirect Link - Fokunang, C.N., A.G.O. Dixon, C.N. Akem and T. Ikotun, 2000. Cultural, morphological and pathogenic variability in Colletotrichum gloeosporioides f.sp. Manihotis isolates from Cassava (Manihot esculenta) in Nigeria. Pak. J. Biol. Sci., 3: 542-546.
CrossRefDirect Link - Fokunang, C.N., A.G.O. Dixon and T. Ikotun, 2003. Synergistic relationship of bacterial blight and anthracnose disease pathogen in cassava multiple infection. J. Biol. Sci., 3: 596-606.
CrossRefDirect Link - Freeman, S., T. Katan and E. Shabi, 1998. Characterization of Colletotrichum species responsible for anthracnose diseases of various fruits. Plant Dis., 82: 596-605.
CrossRefDirect Link - Haque, M.A., K.M. Khalequzzaman, M.S. Islam and M.M. Hossain, 2003. Survey the prevalence of market diseases of banana. Plant Pathol. J., 2: 169-173.
CrossRefDirect Link - Hasna, M.K., M.B. Meah and M.A. Kader, 2000. Assessment of yield loss in guava owing to fruit anthracnose. Pak. J. Biol. Sci., 3: 1234-1236.
CrossRefDirect Link - Klotz, L.J. and H.S. Fawcett, 1932. Black scorch of the date palm caused by Thielaviopsis paradoxa. J. Agric. Res., 44: 155-166.
Direct Link - Kolattukudy, P.E., 1985. Enzymatic penetration of the plant cuticle by fungal pathogens. Annu. Rev. Phytopathol., 23: 233-250.
CrossRefDirect Link - Lopez-Llorca, L.V. and S. Orts, 1994. Histopathology of infection of the palm Washingtonia filifera with pink bud rot fungus Penicillium vermoesenii. Mycol. Res., 89: 1195-1199.
Direct Link - Manandhar, J.B., I.K. Kunwar, T. Singh, G.L. Hartman and J.B. Sinclair, 1985. Penetration and infection of soybean leaf tissues in Colletotrichum truncatumand Glomerella glycines. Phytopathology, 75: 704-708.
Direct Link - Martinez-Culebras, P.V., E. Barrio, M.D. Garcia and A. Querol, 2000. Identification of Colletotrichum species responsible for anthracnose of strawberry based on the internal transcribed spacers of the ribosomal region. FEMS Microbiol. Lett., 189: 97-101.
CrossRef - Morin, L., J.A.L. Derby and E.D. Kokko, 1996. Infection process of Colletotrichum gloeosporioides f. sp. Malvae on Malvaceae weeds. Mycol. Res., 100: 165-172.
Direct Link - Nuangmek, W., E.H.C. McKenzie and S. Lumyong, 2008. Endophytic fungi from wild banana (Musa acuminata Colla) works against anthracnose disease caused by Colletotrichum musae. Res. J. Microbiol., 3: 368-374.
CrossRefDirect Link - Pandey, B.K., U.S. Singh and H.S. Chaube, 1987. Mode of infection of ascochyta blight as caused by Ascochyta rabiei. J. Phytopathol., 119: 88-93.
CrossRef - Al Eryani-Raqeeb, A., T.M.M. Mahmud, S.R.S. Omar, A.R.M. Zaki and A.R. Al Eryani, 2009. Effects of calcium and chitosan treatments on controlling anthracnose and postharvest quality of papaya (Carica papaya L.). Int. J. Agric. Res., 4: 53-68.
CrossRefDirect Link - Sanders, G.M. and L. Korsten, 2003. Comparison of cross inoculation potential of South African avocado and mango isolates of Colletotrichum gloeosporioides. Microbiol. Res., 158: 143-150.
CrossRefDirect Link - Sutton, D.A., 1999. Coelomycetous fungi in human disease. A review: Clinical entities, pathogenesis, identification and therapy. Rev. Iberoam Micol., 16: 171-179.
PubMed - Talhinhas, P., C. Mota-Capitao, S. Martins, A.P. Ramos and J. Neves-Martins et al., 2011. Epidemiology histopathology and aetiology of olive anthracnose caused by Colletotrichum acutatum and C. gloeosporioides in Portugal. Plant Pathol., 60: 483-495.
CrossRef - Taechowisan, T., N. Chuaychot, S. Chanaphat, A. Wanbanjob and P. Tantiwachwutikul, 2009. Antagonistic Effects of Streptomyces sp. SRM1 on Colletotrichum musae. Biotechnology, 8: 86-92.
CrossRefDirect Link - Trevorrow, P.R., J.A.G. Irwin and D.F. Cameron, 1988. Histopathology of compatible and incompatible interactions between Colletotrichum gloeosporioides and Stylosanthes scabra. Trans. Br. Mycol. Soc., 90: 421-429.
Direct Link - Rivera-Vargas, L.I., Y. Lugo-Noel, R.J. McGovern, T. Seijo and M.J. Davis, 2006. Occurrence and distribution of Colletotrichum spp. on mango (Mangifera indica L.) in puerto rico and Florida, USA. Plant Pathol. J., 5: 191-198.
CrossRefDirect Link - Xiao, C.L., S.J. MacKenzie and D.E. Legard, 2004. Genetic and pathogenic analyses of Colletotrichum gloeosporioides from strawberry and noncultivated hosts. Phytopathology, 94: 446-453.
PubMed