
Reshu
Sardar Vallabhbhai Patel University, Krishi Vigyan Kendra, Rampur, India
Mohd. Mahmud Khan
Department of Plant Protection, Faculty of Agricultural Sciences, Aligarh Muslim University, Aligarh, India
Plant Pathology Journal
Year: 2012 | Volume: 11 | Issue: 1 | Page No.: 1-9
DOI: 10.3923/ppj.2012.1.9







ABSTRACT
Trichoderma harzianum, T. viride, T. virens and Aspergillus niger were isolated from mustard leaf surface. Efficacy of their conidial suspension and culture filtrates was evaluated. In dual culture test, T. viride caused maximum inhibition of radial growth of A. brassicae (74%) and A. brassicicola (77%). Highest concentration (3:1) of T. viride culture filtrates caused 60 and 54% inhibition of radial growth of A. brassicae and A. brassicicola, respectively. The culture filtrates of T. viride at their highest concentration (3:1) caused 71 and 69% inhibition of conidial germination of A. brassicae and A. brassicicola, respectively. In field trial foliar spray of 1x107 conidia mL-1 of biological control agents was done on mustard plants 24 h after artificial inoculation by A. brassicae conidial suspension (1x105 conidia mL-1. Viability of BCAs was recorded up to 30 days in phylloplane of mustard at relative humidity between 80-90% and temperatures between 20-25°C. T. viride proved to be most effective as its conidial suspension caused up to 76 and 68% reduction in disease intensity on leaves and pods, respectively. Whereas, its culture filtrate caused 45 and 36% reduction in disease severity on leaves and pods, respectively. These results suggested that T. viride can be used to control Alternaria blight disease of mustard.
PDF Abstract XML References Citation
Received: February 13, 2012;
Accepted: March 21, 2012;
Published: June 27, 2012
How to cite this article
Reshu and Mohd. Mahmud Khan, 2012. Role of Different Microbial-origin Bioactive Antifungal Compounds Against Alternaria spp. Causing Leaf Blight of Mustard. Plant Pathology Journal, 11: 1-9.
DOI: 10.3923/ppj.2012.1.9
URL: https://scialert.net/abstract/?doi=ppj.2012.1.9
DOI: 10.3923/ppj.2012.1.9
URL: https://scialert.net/abstract/?doi=ppj.2012.1.9
INTRODUCTION
Alternaria blight (Alternaria brassicae (Berk.) Sacc.) of mustard group of crops is most important, devastating and widespread disease throughout the world (Kolte, 1985; Khan et al., 2007a; Khan, 2011) on Indian mustard {Brassica juncea (Czern and Coss)}. Alternaria blight causes average yield loss of 46.5% (Kolte et al., 1987). In highly susceptible cultivars of Brassica species, the yield loss may reach up to 70% (Godika et al., 2001) as well as the quality of seed also adversely affects like seed size, oil content and discoloration of seed (Kaushik et al., 1984). The control of Alternaria leaf blight is exclusively based on the application of chemical pesticides. Several effective pesticides have been recommended against the pathogen, but they are not considered to be long term solutions, due to concerns of expense, exposure to health risk, fungicide residue and other environmental hazards. Some alternative methods of control have been adopted in an attempt to modify this condition. Recently, researches have focused on developing, long lasting, environmentally safe and effective biocontrol methods for the plant diseases management. Due to these reasons most of the work done during recent past has been directed towards biological disease management, use of antagonistic fungi in this context appeared logical and safe (Kumar et al., 2005; Latha et al., 2009).
Trichoderma harzianum Rifai, T. Viride Pers. ex Gray are important bio control agents (BCAs) of plant pathogens. These BCAs were used for the control of soil borne, foliar and post harvest diseases in various crops in the field, in commercial green house and storage depots (Papavizas, 1985; Elad, 1994; Adejumo et al., 1999; Sobowale et al., 2005; Abeysinghe, 2009; Jegathambigai et al., 2010). BCAs differ fundamentally from chemical fungicides in that they must grow and proliferate to be effective (Nelson et al., 1994). Trichoderma work against fungal phytopathogens either indirectly by competing for nutrients and space, modifying environmental conditions, promoting plant growth and plant defence mechanism and antibiosis, or directly through mechanisms such as mycoparasitism (Shakeri and Foster, 2007; Reino et al., 2008). Trichoderma spp. suppresses disease by antagonizing the pathogen (Shah and Nasreen, 2011). Some BCAs such as the plant growth promoting organisms (PGPOs), control a disease by dual action, not only suppressing a pathogen but also promoting plant growth and the defence mechanism of plants (Khan and Anwar, 2007; El-Sherif and Ismail, 2009; Mohiddin et al., 2010). Aspergillus niger is a versatile phosphate solubilizer that is abundant in various soil types (Gaur, 1990). A. niger fungus has a fair capacity to suppress plant pathogens (Fujimoto et al., 1993; Khan et al., 2006) and it increases the yield of the plant it colonizes. The effectiveness of the fungus, however, varies with the isolate (Sen et al., 1993; Khan et al., 2007b).
The present study aimed at exploring the possibility of using conidial suspension and culture filtrates of Trichoderma spp. and A. niger in the control of A. brassicae and A. brassicicola causing leaf blight of mustard.
MATERIALS AND METHODS
Isolation of pathogenic fungi: Alternaria brassicae and A. brassicicola were isolated from diseased leaves of mustard and multiplied on Potato Dextrose Agar (PDA) medium in Petri plates. The pathogen inoculated plate(s) were incubated at 25±2°C.
Biological control agents and their culture: Trichoderma harzianum, T. viride, T. virens and A. niger were isolated from mustard leaves. Single spore method was adopted to get pure culture. Pure cultures were multiplied in Petri plates and these plates were incubated at 25±2°C in BOD incubator. Mancozeb was purchased from the local market and concentration was prepared as per requirement. Standard cultures of T. harzianum, T. viride, T. virens and A. niger were procured from Indian Agricultural Research Institute, New Delhi.
In vitro dual culture test: A dual culture test was conducted in vitro to assay the ability of biocontrol agents viz., T. harzianum, T. viride, T. virens and A. niger to inhibit hyphal growth of A. brassicae and A. brassicicola as well as to observe interactions between colonies. Assays were performed in petri plates containing PDA. The mycelial disc of 5 mm diameter was cut from the margin of 7 days old culture of pathogen as well as BCAs. Each disc of pathogen and BCAs was placed on PDA at 5 cm apart from each other. Petri plates inoculated with both the organisms were incubated in BOD incubator at 25± 2°C. A disc of filter paper of 5 mm was placed 5 cm apart from pathogenic fungus after dipped in 0.2% mancozeb solution prepared in double distilled water. Controls were maintained as single organism inoculated in the centre of one individual. Observations were taken on growth of both the organisms. Mycelium was picked from the zone of interaction from plates of dual cultures and then microscopically investigated and the changes in the mycelium of A. brassicae and A. brassicicola were recorded.
Collection of culture filtrates of biological control agents: Culture filtrates of fungal bioagents were obtained by growing biocontrol fungi separately on potato dextrose broth (PDB). Conical flasks of one litre capacity containing 250 mL PDB were inoculated with 5 mm mycelial plugs of 7 days old culture grown on PDA. Culture flasks were placed on an orbital shaker (150 rotations min-1) during the first week. After inoculation, culture flasks were checked visually and flasks showing only pure growth of the respective isolate after 35 days of incubation were taken to collect culture filtrate. The liquid medium was filtered through oven dried Whatman No. 42 filter paper to separate fungal mycelium. Filtrate was collected and was considered as 100% concentration (Vishwanath and Kolte, 1997). The culture filtrate thus obtained was centrifuged at 5000 rpm for 15 min to make cell free and was sterilized by filtration using 0.02 μm millipore filters.
Effect of culture filtrates on biomass of pathogenic fungus: The in vivo efficacy of different concentrations of culture filtrates obtained from BCAs was evaluated on biomass production of pathogenic fungi, A. brassicae and A. brassicicola. It was grown separately on Potato Dextrose Broth (PDB). Conical flask of one litre capacity containing 100 mL of culture filtrate and medium (C:M) in 1:3, 2:2, 3:1 were inoculated with 5 mm mycelial plugs of 10 days old culture grown on PDA. Culture flasks were placed on an orbital shaker (150 rotations min-1) during the first week and observed visually. After eight days of inoculation, fungal growth was harvested by filtration through oven dried Whatman No. 42 filter paper. The fungal mat was washed thoroughly with distilled water and then oven dried at 60°C for 72 h. It was subsequently cooled in desiccators having anhydrous calcium chloride and accurately weighed (Singh et al., 2003). The conical flasks containing 100 mL PDB inoculated with A. brassicae and A. brassicicola as control (check) and maintained by mixing distilled water in place of culture filtrate.
Effect of culture filtrates of BCAs on conidial germination of pathogenic fungi: Conidial suspension were prepared from 10 days old culture of A. brassicae and A. brassicicola grown on plates containing PDA by adding distilled water in Petri plates and scratching with microslides. Spore suspension was passed through four layers of cheese cloth to remove mycelial fragments and centrifuged at 3000 rpm for 5 min to remove unwanted material. The conidial concentration was maintained at 1x105 conidia mL-1 (Chand and Singh, 2004; Patni and Kolte, 2006). Conidial suspension of A. brassicae and A. brassicicola containing at least 15-20 conidia per microscopic field (10x) was prepared. One drop (0.1 mL) of conidial suspension was put over acid cleaned glass cavity slide and allowed to air dry. A drop of desired concentration of culture filtrate (i.e., 1:3, 2:2 and 3:1) of bioagent was subsequently put on it. The slides were kept in moist chamber prepared by putting two folds of Whatman filter paper wet with double distilled water, in both sides of plates (Kumar et al., 2004). These plates were incubated at 25±2°C for 24 h in BOD incubator. The double distilled water was added in place of culture filtrate as control treatment. Observations on percent conidial germination and inhibition were recorded after 24 h of inoculation. To comprehend the sensitivity of A. brassicae and A. brassicicola conidia to culture filtrates of bioagents based on conidia germination test, observations on conidia germination were recorded by examining three microscopic fields (10x) on each of the 3 slides of a treatment. The data was produced as:
Effect of culture filtrates on radial growth of pathogenic fungus: The effect of culture filtrates on radial growth of A. brassicae and A. brassicicola were evaluated and culture filtrates of antagonists were mixed in PDA. PDA was prepared with additional 5 g L-1 agar in order for the medium to solidify after mixing the filtrate and medium at the ratio 1:3, 2:2 and 3:1 (culture filtrate: PDA v/v) separately prior to pouring and then 20 mL PDA was poured in each Petri dish and were inoculated by 5 mm fungal mats of 7 days old culture of A. brassicae and A. brassicicola (Singh and Singh, 2003) and were incubated for 7 days. Control was maintained with distilled water in place of culture filtrate.
In vivo evaluation of culture filtrate and conidial suspension on disease development: The effect of culture filtrates and conidial suspension of biological control agents was evaluated on A. brassicae and a field trial was laid out in Randomized Block Design (RBD). The plot size of 4x3 m-2 was used with three replications. Brassica juncea cv. Varuna was used as susceptible host. Each plot was covered by 4 m height of transparent polythene sheet supported by wooden sticks in a box shape to prevent inter plot effect of inoculum. The pathogen, A. brassicae was inoculated on control plants (check). Seeds were sown at row to row spacing of 30 cm and plant to plant spacing of 10 cm on last week of October. For artificial inoculation of plants conidial suspension of pathogenic fungi with conidia load of 1x105 conidia mL-1 of distilled water was prepared and sprayed on leaves with an atomizer. After 24 h, leaves were sprayed with conidial suspension (1x107 conidia mL-1) and undiluted culture filtrates of different BCAs as well as 0.2% mancozeb separately by an atomizer, independently. Surfactant Triton x-100 was added to conidial suspension and culture filtrates of pathogens and BCAs @ 0.1 (v/v). Observations were taken on each plot in which 1, 2 and 3 sprays were applied. First spray was done after 24 h of inoculation, second spray after 30 days and third spray after 60 days of first spray. Data on disease severity (on leaves after 95 DAS and on pods after 130 DAS) and number of spots (on leaves and pods) were taken. Disease severity was recorded following the scale of Conn et al. (1990). Percent Disease Intensity (PDI) was calculated using formula:
Statistical analysis: The experiment was conducted during two consecutive years. Each treatment was replicated thrice. The data obtained during the two years were statistically identical. So, the results are based on the experiment conducted during the second year. As the second year experiments were significantly performed with greater precision and perfection gained than the experience of the first year. All data were subjected to analysis of variance (ANOVA) using MINITAB 11.0 software. Means were then separated using the Least Significant Difference (LSD) test at p≤0.05, 0.001 and 0.001. The F-values were calculated and compared with the table value at p≤0.05, 0.01 and 0.001.
RESULTS
In vitro dual inoculation test: In biological control, the most important principle of choosing an antagonistic organism is its capability to grow fast and occupy most of the place in the site of probable exist of the pathogen. For this reason, T. harzianum, T. viride, T. virens and A. niger were evaluated against A. brassicae and A. brassicicola using dual inoculation test. Mancozeb was used to compare the Alternaria growth inhibition with BCAs.
| Table 1: | Effect of different biological control agents on radial growth of A. brassicae and A. brassicicola in dual culture |
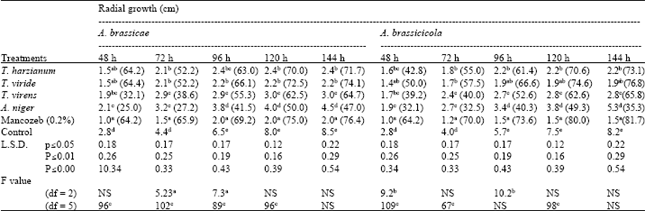 | |
| Figures in parenthesis are % control over check, NS: Non significant, Each value is a mean of three replications, In column, means followed by a common letter are not significantly different at p≤0.05 of DMRT | |
It caused maximum growth inhibition of A. brassicae of 76.4% and A. brassicicola of 81.7%. Among biocontrol agents, T. viride was recorded to be as effective as mancozeb. T. viride caused 74.1 and 76.8% inhibition of radial growth of A. brassicae and A. brassicicola, respectively (Table 1). Though, A. brassicae growth inhibition was reduced significantly equal with mancozeb, T. harzianum and T. viride. Aspergillus niger appeared to be least inhibitory to A. brassicae and A. brassicicola. The visual observation of interaction was quite interesting. The growth of A. brassicae and A. brassicicola was restricted by T. viride and overgrew on pathogenic fungus. There was distinct change in colour at the zone of contact, which became light greenish yellow in case of A. brassicae as well as A. brassicicola. No distinct zone of contact or colour change was observed in case of other biocontrol agents. The growth of A. brassicae and A. brassicicola was restricted by T. harzianum followed by over growth on Alternaria spp. However, A. niger could not restrict growth of A. brassicae and A. brassicicola, only a slight decrease in growth of pathogenic fungus was recorded. Regular growth of both the fungus was observed in dual culture Petri plates.
Effect of culture filtrates and fungicide on dry mass of pathogenic fungus: Dry mass of fungus consists of hyphae, conidia and/or any other resting structure. Estimation of dry mass of fungus is an effective tool to find out the efficacy of any control measure against fungus as due to their effect, overall growth of target fungus gets affected. Among all the treatments, maximum reduction in dry weight was caused by mancozeb. Among BCAs, T. viride caused more than 47% reduction in fungal dry weight and appeared as the most effective BCAs to inhibit biomass production of A. brassicae and A. brassicicola (Table 2).
Effect of culture filtrates and fungicide on radial growth of pathogenic fungi: Mancozeb was observed to be most inhibitory to radial growth of A. brassicae and A. brassicicola. Radial growth of pathogenic fungi was significantly (≤0.05) affected by culture filtrates of different BCAs (Table 2). Maximum reduction in radial growth of A. brassicae and A. brassicicola was recorded due to T. viride followed by T. harzianum culture filtrates. A. niger culture filtrates were least effective in inhibiting radial growth of A. brassicae and A. brassicicola.
Effect of culture filtrates on conidial germination of pathogenic fungi: Culture filtrates of different BCAs significantly inhibited spore germination of A. brassicae and A. brassicicola. Mancozeb caused maximum inhibition of conidial germination of A. brassicae and A. brassicicola (Table 3). Among BCAs, T. viride was recorded to be the inhibitoriest to conidial germination of pathogenic fungi. A. niger culture filtrate was least effective to inhibit conidial germination of pathogenic fungi. With increase in culture filtrate concentration increase in inhibitory effect on conidial germination was recorded.
In vivo evaluation of efficacy of culture filtrates and conidial suspension of biocontrol agents on A. brassicae: Significant difference (≤0.05) between treatments and number of sprays were recorded for all the components of disease development viz., number of spots on leaves and pods and percent disease intensity on leaves and pods. Least number of spots i.e., 6.0, 2.3 and 2.0 were observed after 1 spray, 2 sprays and 3 sprays of conidial suspension of T. viride, respectively (Table 4).
| Table 2: | Effect of different concentrations (C:M) of culture filtrates of different biological control agents on radial growth and dry mass of A. brassicae and A. brassicicola |
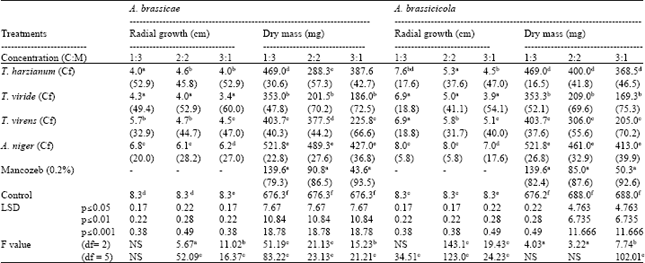 | |
| Figures in parenthesis are % control over check, NS: Non significant, Cf: Culture filtrates, each value is a mean of three replications, in column, means followed by a common letter are not significantly different at 5% level of DMRT | |
| Table 3: | Effect of different concentrations (C:M) of culture filtrates of different biological control agents on spore germination of A. brassicae and A. brassicicola |
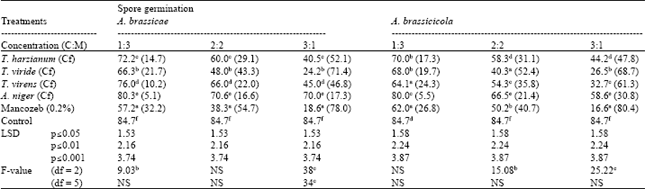 | |
| Figures in parenthesis are % control over check, NS: Non significant, Cf: Culture filtrates, Each value is a mean of three replications. In column, means followed by a common letter are not significantly different at 5% level of DMRT | |
| Table 4: | Effect of different number of sprays of biological control agents on leaf blight of mustard caused by Alternaria brassicae |
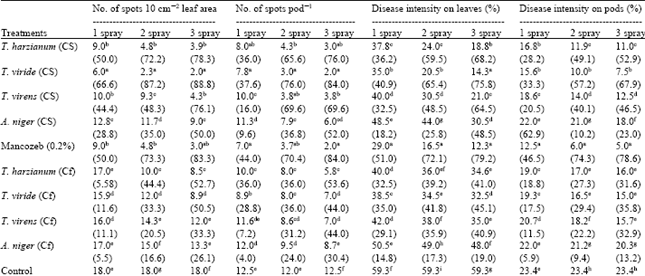 | |
 | |
| Figures in parenthesis are % control over check, NS: Not significant, Cf: Culture filtrate, Each value is a mean of three replications. In column, means followed by a common letter are not significantly different at 5% level of DMRT | |
Mancozeb caused up to 83% reduction in number of spots 10 cm-2 leaf area. Trichoderma viride and mancozeb were significantly at par or equally effective to control No. of spots 10 cm-2 leaf area, on pods and reduction of percent disease intensity on leaves after 3 sprays of each of them, separately (Table 4). In case of culture filtrates, T. harzianum culture filtrates were more effective than T. viride to reduce number of spots/unit area of leaf and number of spots/pod. Maximum reduction in disease severity was recorded after foliar spray of mancozeb. However, T. viride conidial suspension as well as culture filtrate spray was found to be the best among all the biocontrol agents to decrease disease intensity. Efficacy of conidial suspension, culture filtrates and mancozeb increased with increase in frequency of spray. Culture filtrates were less effective than conidial suspension against all the parameters of disease development as its foliar spray using similar method and number of sprays. Conidial suspension as well as culture filtrates of A. niger were least effective to decrease No. of spots/unit area.
DISCUSSION
In the present study, fastest growth was attained by Trichoderma spp. The rapid growth gives Trichoderma an important advantage in the competition for space and nutrients with plant pathogenic fungi even before deploys arsenal of mycotoxins (Barbosa et al., 2001). Accordingly in present study T. viride caused up to 76.8% inhibition of radial growth of Alternaria spp. Kumar et al. (2005) and Abd-El-Kareem (2007) observations on Alternaria alternata caused disease in bean plants are supporting to this study. Microscopic observations on hyphal interactions between antagonists and A. brassicae and A. brassicicola revealed lysis and protoplasmic disruption of hyphae of test fungus at many locations. Mycoparasitism through physical contact with coiling and pathogen cell lysis in case of Sclerotium rolfsii by T. harzianum has been reported by Upadhyay and Mukhopadhyay (1986). The disintegration of mycelium of test fungi may be due to action of enzymes produced by Trichoderma spp. (Elad et al., 1982) and production of volatile and non volatile chemical compounds (Upadhyay and Roy, 1995) and toxin (Brain and McGowan, 1945). Jackson et al. (1991) reported that Gliocladium virens, T. pseudokoningii and T. virens produced antifungal metabolites which were effective in reducing the radial growth of Rhizoctonia solani and Botrytis cinerea as well as Sclerotium cepivorum.
In present study, culture filtrates of different biological control agents were used to evaluate their efficacy in inhibiting radial growth of pathogenic fungus i.e., Alternaria and its spore germination. T. viride caused 21.7 and 19.7% inhibition of conidial germination at its lowest concentration. Dennis and Webster (1971) and Jinantara (1995) observed that culture filtrates produced by Trichoderma contained inhibitory substances against microorganisms. Among them antibiotics produced by T. harzianum were 6-n-pentyl-2H-Pyran-one. In present study, T. viride culture filtrates were more effective (up to 25% ) than T. virens culture filtrates. This finding is in the contradiction to the observation taken by Jackson et al. (1991). They reported T. virens culture filtrates were more effective than T. viride culture filtrates, though, they have tested these BCAs against S. cepivorum not Alternaria spp. It has been observed in the present study that the efficacy of culture filtrates in inhibition of radial growth and conidial germination increased with increase in their concentration. Effectiveness of T. viride culture filtrates increased up to 50% with increase in its concentration from 1:3 to 3:1 (culture filtrate: Medium). Similar observations were reported by Imtiaj and Lee (2008) on onion blotch disease caused by Alternaria porri. Pandey et al. (1993) had also reported that culture filtrates of A. niger caused more than 50% inhibition of Colletotrichum gleosporioides. Volatiles produced from the culture of A. niger were also reported to be effective against Colletotrichum gleosporioides. Brunner et al. (2005) reported increase in the ability of a biocontrol strain of T. atroviride to activate systemic resistance response in the host plant by using a glucose oxidase encoding gene obtained from another fungus (A. niger). Aspergillus niger aggregate is a contact as well as an invasive necrotroph (Mondal et al., 2000; Abarca et al., 2004) and grows and sporulate well on the mycelium of fungi such as Rhizoctonia solani (Sen et al., 1995). The production of NH3, HCN and siderophore might have contributed in the suppression of R. solani (Nair and Burke, 1988; Palakshappa et al., 1989).
Effectiveness of a biocontrol agent is related to its potential to proliferate and survive on the plants for a prolonged period after application, mainly on tissues that are usually prone to infection by the target pathogen (Elad, 1990). Therefore, in vivo studies, foliar spray of conidial suspension and culture filtrates was done to ascertain their efficacy against A. brassicae. Biocontrol agents can be applied as foliar spray Alternaria spp. are necrotrophic pathogen and they mainly produce symptoms on leaves. Biocontrol agents must normally achieve a high population in the phylloplane to control other organisms, but colonization by the BCAs may be reduced by competition with the indigenous microflora (Kubota, 1996) or other factors. Application of BCAs, operating through nutrient competition to healthy leaves, however, is generally not very effective, because the pathogen rapidly penetrates the leaf and escape competition (Fokkema, 1993).
Effectiveness of T. viride increased up to 35% with increase in its number of sprays from one spray to three sprays at 30 days interval. Three sprays of T. viride decreased disease severity by 76% that is as effective as mancozeb (0.2%) which caused 79% reduction in disease severity on leaves. Similar results were recorded by Elad et al. (1993), Jegathambigai et al. (2010) and Mir et al. (2011). They sprayed a conidial preparation of T. harzianum on cucumber plants in greenhouse in order to control fruit and stem grey mold. Up to 90% control was achieved by the biocontrol agent (0.5, 1.0 g L-1). In most experiments in commercial conditions was as effective as dicarboxamide fungicides, iprodione or vinclozolin (0.5 g L-1 each). Population of T. harzianum were on a level of 3x105-8x105 CFU leaf-1 and ten times lower on the fruit. They remained higher after the second and third sprays. Nutrient changes preceding germination in Trichoderma conidia can either enhance or decrease their biocontrol potential, depending on environmental conditions in the microhabitat (Hjeljord and Tronsmo, 2003). Perello et al. (2009) reported that T. harzianum and T. koningii significantly reduced tan spot of wheat caused by Pyrenophora tritici-repentis under field conditions.
Results of the present study are very encouraging to establish Trichoderma spp. In particular, Trichoderma viride as an effective tool to control leaf spot disease of mustard whereas; A. niger does not appear to be effective. Further studies are conducted to understand their adaptable microclimate and environmental requirements.
ACKNOWLEDGMENTS
The authors are thankful to University Grants Commission, New Delhi for granting fellowship and Dr. Mujeebur Rahman Khan (Fellow of National Academy of Agricultural Sciences, New Delhi) Associate Professor, Department of Plant Protection, Aligarh Muslim University, Aligarh. The authors are also grateful to the Department of Plant Protection and the Aligarh Muslim University for providing necessary facilities. Both the author has equal authorship and equal contribution in this study.
REFERENCES
- Abeysinghe, S., 2009. Effect of combined use of Bacillus subtilis CA32 and Trichoderma harzianum RU01 on biological control of Rhizoctonia solani on Solanum melongena and Capsicum annuum. Plant Pathol. J., 8: 9-16.
CrossRefDirect Link - Abarca, M.L., F. Accensi, J. Cano and F.J. Cabanes, 2004. Taxonomy and significance of black aspergilli. Antonie Van Leeuwenhoek, 86: 33-49.
PubMed - Abd-El-Kareem, F., 2007. Induced resistance in bean plants against root rot and alternaria leaf spot diseases using biotic and abiotic inducers under field conditions. Res. J. Agric. Biol. Sci., 3: 767-774.
Direct Link - Adejumo, T.O., T. Ikotun and D.A. Florini, 1999. Biological control of Protomycopsis phaseoli, the causal agent of leaf smut of cowpea. J. Phytopathol., 147: 371-375.
CrossRefDirect Link - Barbosa, M.A.G., K.G. Rehn, M. Menezes and R. de L.R. Mariano, 2001. Antagonism of Trichoderma species on Cladosporium herbarum and their enzymatic characterization. Braz. J. Microbiol., 32: 98-104.
Direct Link - Brain, P.W. and J.G. McGowan, 1945. Viridin: A highly fungistatic substance produced by Trichoderma viride. Nature, 156: 144-145.
CrossRefDirect Link - Brunner, K., S. Zeilinger, R. Ciliento, S.L. Woo, M. Lorito, C.P. Kubicek and R.L. Mach, 2005. Improvement of the fungal biocontrol agent Trichoderma atroviride to enhance both antagonism and induction of plant systemic disease resistance. Applied Environ. Microbiol., 71: 3959-3965.
CrossRefDirect Link - Conn, K.L., J.P. Tewari and R.P. Awasthi, 1990. A disease assessment key for Alternaria blackspot in rapeseed and mustard. Can. Plant Dis. Surv., 70: 19-22.
Direct Link - Dennis, C. and J. Webster, 1971. Antagonistic properties of species-groups of Trichoderma. II. Production of volatile antibiotics. Trans. Br. Mycol. Soc., 57: 41-48.
CrossRef - El-Sherif, A.G. and A.F.A. Ismail, 2009. Integrated management of Meloidogyne incognita infecting soybean by certain organic amendments, Bacillus thuringiensis, Trichoderma harzianum and oxamyl with reference to NPK and total chlorophyll statuus. Plant Pathol. J., 8: 159-164.
CrossRefDirect Link - Elad, Y., 1990. Reasons for the delay in development of biological control of foliar pathogens. Phytoparasitica, 18: 99-105.
CrossRefDirect Link - Elad, Y., I. Chet and Y. Henis, 1982. Degradation of plant pathogenic fungi by Trichoderma harzianum. Can. J. Microbiol., 28: 719-725.
CrossRefDirect Link - Elad, Y., G. Zimand, Y. Zaqs, S. Zuriel and I. Chet, 1993. Use of Trichoderma harzianum in combination or alternation with fungicides to control cucumber grey mould (Botrytis cinerea) under commercial greenhouse conditions. Plant Pathol., 42: 324-332.
CrossRefDirect Link - Fokkema, N.J., 1993. Opportunities and problems of control of foliar pathogens with micro-organisms. Pestic. Sci., 37: 411-416.
CrossRefDirect Link - Fujimoto, Y., H. Miyagawa, T. Tsurushima, H. Irie, K. Okamura and T. Ueno, 1993. Structures of antafumicins A and B, novel antifungal substances produced by the fungus Aspergillus niger NH-401. Biosci. Biotechnol. Biochem., 57: 1222-1224.
Direct Link - Mir, G.H., L.S. Devi, S. Ahmad, V.M. Kumar and P. Williams, 2011. Antagonistic potential of native isolates of Trichoderma viride on corm rot pathogen complex of saffron (Crocus sativus) in Kashmir. Plant Pathol. J., 10: 73-78.
CrossRefDirect Link - Hjeljord, L.G. and A. Tronsmo, 2003. Effect of germination initiation on competitive capacity of Trichoderma atroviride P1 conidia. Phytopathology, 93: 1593-1598.
CrossRefPubMedDirect Link - Imtiaj, A. and T.S. Lee, 2008. Antagonostic effect of three Trichoderma species on the Alternaria porri pathogen of onion blotch. World J. Agric. Sci., 4: 13-17.
Direct Link - Jackson, A.M., J.M. Whipps, J.M. Lynch and M.J. Bazin, 1991. Effects of some carbon and nitrogen sources on spore germination, production of biomass and antifungal metabolites by species of Trichoderma and Gliocladium virens antagonistic to Sclerotium cepivorum. Biocontrol Sci. Technol., 1: 43-51.
CrossRefDirect Link - Jegathambigai, V., R.S.W. Wijeratnam and R.L.C. Wijesundera, 2010. Effect of Trichoderma sp. on Sclerotium rolfsii, the causative agent of collar rot on Zamioculcas zamiifolia and an on farm method to mass produce Trichoderma species. Plant Pathol. J., 9: 47-55.
CrossRefDirect Link - Khan, M.R. and A.M. Anwer, 2007. Molecular and biochemical characterization of soil isolates of Aspergillus niger aggregate and an assessment of their antagonism against Rhizoctonia solani. Phytopathologia Mediterranea, 46: 304-315.
Direct Link - Kolte, S.J., R.P. Awasthi and Vishwanath, 1987. Assessment of yield losses due to Alternaria blight in rapeseed and mustard. Indian Phytopathol., 40: 209-211.
Direct Link - Kumar, S., J.P. Upadhyay and S. Kumar, 2005. Biocontrol of Alternaria leaf spot of Vicia faba using antagonistic fungi. J. Biol. Control, 20: 247-251.
Direct Link - Latha, P., T. Anand, N. Ragupathi, V. Prakasam and R. Samiyappan, 2009. Antimicrobial activity of plant extracts and induction of systemic resistance in tomato plants by mixtures of PGPR strains and Zimmu leaf extract against Alternaria solani. Biol. Control, 50: 85-93.
CrossRefDirect Link - Mohiddin, F.A., M.R. Khan, S.M. Khan and B.H. Bhat, 2010. Why Trichoderma is considered super hero (super fungus) against the evil parasites? Plant Pathol. J., 9: 138-148.
Direct Link - Mondal, G., P. Dureja and B. Sen, 2000. Fungal metabolites from Aspergillus niger AN27 related to plant growth promotion. Indian J. Exp. Biol., 38: 84-87.
PubMedDirect Link - Nair, M.G. and B.A. Burke, 1988. A new fatty acid methyl ester and other biologically active compounds from Aspergillus niger. Phytochemistry, 27: 3169-3173.
CrossRef - Pandey, R.R., D.K. Arora and R.C. Dubey, 1993. Antagonistic interaction between fungal pathogens and phyloplane fungi of guava. Mycopathology, 124: 31-39.
Direct Link - Papavizas, G.C., 1985. Trichoderma and Gliocladium: Biology, ecology and potential for biocontrol. Ann. Rev. Phytopathol., 23: 23-54.
Direct Link - Patni, C.S. and S.J. Kolte, 2006. Effect of some botanicals in management of Alternaria blight of rapeseed-mustard. Ann. Plant Prot. Sci., 14: 151-156.
Direct Link - Reino, J.L., R.F. Guerrero, R. Hernandez-Galan and I.G. Collado, 2008. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev., 7: 89-123.
CrossRefDirect Link - Sen, B., K. Mukherjee, C. Chattopadhyay, A.K. Patibanda and J. Sharma, 1995. Aspergillus niger, A potential biocontrol agent for soil borne plant pathogen. Proceedings of the Global Conference on Advances in Research on Plant Pathogens and their Management, January 18-20, 1995, Society of Mycology and Plant Pathology, Rajasthan Agricultural University, Udaipur, pp: 19.
- Shah, S. and S. Nasreen, 2011. Evaluation of bioagents against the infection of green mold (Trichoderma spp.) in Pleurotus sajor-caju cultivation. Int. J. Plant Pathol., 2: 81-88.
CrossRef - Shakeri, J. and H.A. Foster, 2007. Proteolytic activity and antibiotic production by Trichoderma harzianum in relation to pathogenicity to insects. Enzyme Microbial. Technol., 40: 961-968.
Direct Link - Singh, R.B. and R.N. Singh, 2005. Fungal pathology: Fungicidal management of foliar diseases of mustard in mid-eastern India. Indian Phytopathol., 58: 51-56.
Direct Link - Singh, R.P., A.K. Singh and R.N. Singh, 2003. Effect of neem products on the growth of Alternaria triticina. Ann. Plant Prot. Sci., 11: 384-386.
Direct Link - Singh, S.K. and R.H. Singh, 2003. Effect of culture filtrate of bio-agents and bio-pesticides on Fusarium udum. Ann. Plant Prot. Sci., 11: 387-388.
Direct Link - Upadhyay, J.P. and A.N. Mukhopadhyay, 1986. Biological control of Sclerotium rolfsii by Trichoderma harzianum in sugarbeet. Trop. Pest Manage., 32: 215-220.
CrossRefDirect Link - Vishwanath and S.J. Kolte, 1997. Variability in Alternaria brassicae: Response to host genotypes, toxin production and fungicides. Indian Phytopathol., 50: 373-381.
Direct Link