
Askarne Latifa
Laboratoire De Biotechnologies et Valorisation Des Ressources Naturelles, Equipe De Phytopathologie et Lutte Integree, Ibn Zohr University, Faculty of Sciences, B.P 8106, Agadir, Morocco
Talibi Idriss
Laboratoire De Biotechnologies et Valorisation Des Ressources Naturelles, Equipe De Phytopathologie et Lutte Integree, Ibn Zohr University, Faculty of Sciences, B.P 8106, Agadir, Morocco
Boubaker Hassan
Laboratoire De Biotechnologies et Valorisation Des Ressources Naturelles, Equipe De Phytopathologie et Lutte Integree, Ibn Zohr University, Faculty of Sciences, B.P 8106, Agadir, Morocco
Serghini Mohammed Amine
Laboratoire De Biotechnologies et Valorisation Des Ressources Naturelles, Equipe De Phytopathologie et Lutte Integree, Ibn Zohr University, Faculty of Sciences, B.P 8106, Agadir, Morocco
Boudyach El Hassane
Laboratoire De Biotechnologies et Valorisation Des Ressources Naturelles, Equipe De Phytopathologie et Lutte Integree, Ibn Zohr University, Faculty of Sciences, B.P 8106, Agadir, Morocco
Ait Ben Aoumar Abdellah
Laboratoire De Biotechnologies et Valorisation Des Ressources Naturelles, Equipe De Phytopathologie et Lutte Integree, Ibn Zohr University, Faculty of Sciences, B.P 8106, Agadir, Morocco
Plant Pathology Journal
Year: 2011 | Volume: 10 | Issue: 3 | Page No.: 99-107







ABSTRACT
Control of Citrus blue mold, caused by Penicillium italicum, has been accomplished by postharvest application of synthetic fungicides. However, the development of resistant fungal strains and increasing public concern over food safety and the environment are driving a search for alternative disease control strategies. In vitro trials were conducted to evaluate the effect of several organic acid and salt compounds on mycelial growth, sporulation, spore germination and germ tube elongation of Penicillium italicum. Among 28 tested compounds, sodium carbonate, ammonium carbonate, copper sulfate, EDTA and sodium metabisulfite completely inhibited mycelial growth and sporulation of Penicillium italicum at only 0.02 M. The lowest Minimum Inhibitory Concentration (MIC), Minimum Fungicidal Concentration (MFC) and EC50 values were recorded in sodium metabisulfite treatment. All tested compounds, except Nicotinic acid, strongly inhibited spore germination and germ tube elongation in a dose-dependent manner. Results from this study provide an important basis for further study into the uses of salt compounds for the control of blue mold of Citrus fruit under semi-commercial conditions.
PDF Abstract XML References Citation
Received: July 04, 2011;
Accepted: August 10, 2011;
Published: November 15, 2011
How to cite this article
Askarne Latifa, Talibi Idriss, Boubaker Hassan, Serghini Mohammed Amine, Boudyach El Hassane and Ait Ben Aoumar Abdellah, 2011. Effects of Organic Acids and Salts on the Development of Penicillium italicum: The Causal Agent of Citrus Blue Mold. Plant Pathology Journal, 10: 99-107.
DOI: 10.3923/ppj.2011.99.107
URL: https://scialert.net/abstract/?doi=ppj.2011.99.107
DOI: 10.3923/ppj.2011.99.107
URL: https://scialert.net/abstract/?doi=ppj.2011.99.107
INTRODUCTION
Citrus is a major fruit crop that is grown around the World (Smilanik et al., 2005). In most producing countries the crop is largely grown for fresh fruit consumption. It is the first exporting agricultural sector in Morocco, with the Souss-Massa-Draa (SMD) Valley as the main area of both production and exportation of fresh fruit (Boubaker et al., 2009).
The most important diseases that cause commercially significant losses, in Morocco (Elkhamass et al., 1994) and worldwide (Eckert and Eaks, 1989; Holmes and Eckert, 1999; Zhu et al., 2006) are green mold, caused by Penicillium digitatum (pers.:Fr.) Sacc. and blue mold, caused by P. italicum wehmer. These Penicillium species are strict wound pathogens, they are ubiquitous and produce profuse amount of asexual conidia that are readily disseminated by air current (Holmes and Eckert, 1995, 1999; Boubaker et al., 2009). Therefore they can infect the fruit in the grove, the packinghouses and marketing, through wounds occurred during harvest and subsequent handling (Brown and Miller, 1999; Boubaker et al., 2009). Blue mold is more harmful because it spreads in the box and healthy fruits are directly attacked, regardless of injury. This disease is, also, more important under cold storage conditions. Currently, measures employed to manage such fungal diseases involve the application of chemical fungicides usually incorporated into waxes (Smilanick and Sorenson, 2001; Boubaker et al., 2009). However, the use of fungicides is becoming increasingly restricted due to stringent regulation, pathogen resistance development and growing public concern about chemical residues in fruit (Zhang and Swingle, 2003; Palou et al., 2008). Therefore, alternative methods for decay control are needed. Various alternative measures such as the application of microorganisms (El-Ghaouth et al., 2000; Taqarort et al., 2008), plant extracts (Tripathi and Dubey, 2004; Ameziane et al., 2007; Rani and Devanand, 2011), Essential oils (Barrera-Necha et al., 2008, 2009) or the use of effective natural substances like food additives (Kazemi et al., 2011; Shirzadeh and Kazemi, 2011) have been developed. Food additives have minimal adverse effect on the environment and health (Arslan et al., 2009) and exhibit a broad-spectrum antifungal activity (Corral et al., 1988). These alternative measures have been applied in combination or in a replacement for fungicide in order to reduce the input of fungicides and to prolong the shelf life of citrus fruit, particularly in a market where transport from producer to consumers may take several weeks (Boubaker et al., 2009). Several studies have dealt with the use of different salt compounds to control various post-harvest diseases of citrus and other crops (Arslan et al., 2006; Nigro et al., 2006; Arslan et al., 2009). Treatment of fruit with carbonate or bicarbonate salts was shown to reduce the incidence of post-harvest diseases of citrus fruit caused by Penicillium digitatum, Penicillium italicum or Geotrichum candidum (Zhang and Swingle, 2003; Smilanick et al., 2006; Smilanick et al., 2008). They have also been used to control the blue mold caused by Penicillium expansum and the gray mold caused by Botrytis cinerea in apple fruit (Droby et al., 2003; Palou et al., 2009). Sodium metabisulfite was shown to reduce potato silver scurf caused by Helminthosporiun solani (Olivier et al., 1998; Hervieux et al., 2002; Mills et al., 2006) and potato dry rot caused by Fusarium sambucinum (Mecteau et al., 2002). EDTA was used to control P. digitatum on oranges (Valencia-Chamorro et al., 2008) and B. cinerea on apple fruit (Droby et al., 2003).
The present work was performed to evaluate the efficacy of a range of organic acids and salts, for in vitro control of the mycelial growth, sporulation and spore germination of Penicillium italicum, the causal agent of blue mold of citrus fruit.
MATERIALS AND METHODS
Pathogen culture: P. italicum was isolated from naturally infected lemon fruit after storage of several weeks. This isolate was the most aggressive one in our collection and produced the largest lesions on inoculated fruit. This fungus was purified and maintained on Potato Dextrose Agar (PDA) and stored at 4°C, with periodic transfers through citrus fruit to maintain its aggressiveness.
In vitro screening of chemicals: The inhibitory effects of 28 Organic acids and salts (Table 1) on mycelial growth of Penicillium italicum were tested in vitro using the agar dilution technique. An aqueous solution of each compound was prepared in sterile distilled water and was added aseptically to molten (50°C) sterile PDA to achieve a final concentrations of 0.02 and 0.2 M before pouring the medium into petri plates (15 mL PDA/plate). Chemicals unamended plates served as control.
| Table 1: | Chemicals used in this study |
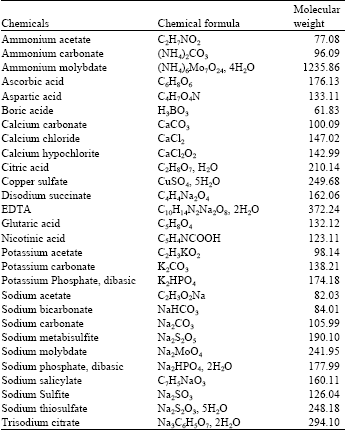 | |
Hyphal plugs (5 mm diameter) were cut from the periphery of actively growing colonies (7 to 10 day-old) and transferred aseptically, mycelium down, to three replicate Petri plates containing PDA medium supplemented with chemicals. The plates were sealed with parafilm and incubated in the dark at 25°C. Radial growth was measured daily at two perpendicular colony diameters until the growth in the control plates reached the edge of the Petri plates. The antifungal activity was expressed in terms of percentage of Mycelial Growth Inhibition (MGI) and calculated according to the following formula:
Compounds that allowed more than 50% of mycelial growth inhibitions in the first screening were further retained for determination of the effective concentration causing a 50% reduction (EC50) in the linear growth on PDA using probit analysis (POLO software). The Minimum Inhibitory Concentration (MIC) and Minimum Fungicidal Concentration (MFC) were also determined in parallel experiment.
To determine whether compounds have fungistatic or fungicidal effect on Penicillium italicum, Plugs from treatment with no growth were transferred to chemical unamended PDA, treatment in which mycelial growth did not occur after additional 9 days of incubation were considered fungicidal (Hervieux et al., 2002; Mecteau et al., 2002). The experiments were performed twice.
Effect of chemicals on sporulation: Sporulation was assessed using chemical amended PDA as described above, only for compounds with more than 50% of mycelium growth inhibition. Once fungal colony in control reached the edge of the plate, amended plates as well as control were flooded with 10 mL of sterile distilled water containing 0.05% (w/v) tween 80 and spores were gently dislodged from the medium using a bacteriological loop. The resulted suspension was filtered through a Buchner funnel and the spores were determined using a heamacytometer. The result was expressed as a percent sporulation inhibition using the following formula:
Three replicates were made for each chemical.
Effect of chemicals on spore germination and germ tube elongation: A spore-suspension was obtained from one-week-old culture of Penicillium italicum, grown on PDA at 25°C, by flooding the culture with 10 mL-1 of sterile distilled water containing 0.05% (v/v) Tween 80. The spores were gently dislodged from the surface and the suspension was filtered through two layers of sterile cheesecloth to remove hyphal fragments. The spore concentration of the suspension was adjusted to 2.105 spores mL-1 with the aid of a heamacytometer (Gabler and Smilanick, 2001). The germination and germ tube elongation of conidia of P. italicum were determined in concentrations of 0.002, 0.005, 0.01, 0.02 and 0.04 M for the compounds that allowed a mycelial growth inhibition superior to 50%. Aliquots (40 μL) of spore suspensions were aseptically transferred in triplicate to sterile depression slides containing 40 μL of 2% sterile and filtered oranges juice amended with different concentrations of salts (Droby et al., 2003). Inoculated slides were placed on moist filter paper in Petri plates and then incubated at 25°C for 18 h. Each slide was then fixed with acid fuchsine solution to stop further germination (Smilanick et al., 1999). Spore germination and germ tube elongation was determined in three microscopic fields using 10x40 ocular micrometers. At least 100 conidia within each replicate were observed. A spore was scored as germinated when the germ tube extended to at least twice the length of the spore itself (Punja and Gaye, 1993; Mills et al., 2004; Amiri and Bompeix, 2011). Germination and germ tube elongation were assessed microscopically at various concentrations of tested chemicals compared with control treatment. The results were expressed as percent spore germination or germ tube elongation inhibition. Each treatment included three replicates and the test was conducted twice.
Statistical analysis: All data were subjected to statistical Analysis of Variance (ANOVA) using STATISTICA software, version 6, Stat- Soft, 2001, France. Percentage values of inhibition were subjected to arcsine-square root transformation before analysis of variance. Duncan multiple range tests were used to segregate treatments which were significantly different at p<0.05. The EC50 values were calculated for each compound by probit analysis using POLO software.
RESULTS AND DISCUSSION
Effects on mycelium growth and sporulation: The in vitro antifungal activities of organic acids and salts were first examined at the concentrations of 0.02 and 0.2 M and showed a variable effects of tests compounds on P. italicum mycelial growth and sporulation (Table 2). Results also showed that after 7 days incubation at 25°C, most tested compounds reduced both mycelial growth and sporulation of P. italicum on PDA medium. It was also noticed that the reduction in growth and sporulation were correlated to the increase in compounds concentrations. Complete inhibition of growth and sporulation was achieved by sodium carbonate, ammonium carbonate, copper sulfate, sodium metabisulfite and EDTA at the lower tested concentration (0.02 M). Ammonium molybdate reduced also the mycelial growth and sporulation of the pathogen, on PDA medium amended with 0.02 M of this salt.
Boric acid and potassium carbonate reduced the mycelial growth by more than 50%. However, potassium carbonate appears to be more effective than boric acid when sporulation inhibition percentages were compared (68.89% versus 30.05%). The remaining compounds inhibited mycelial growth by less than 50%. In contrast, sodium acetate at 0.02 M enhanced the mycelial growth of P. italicum.
Tested at 0.2 M, 14 out of 28 studied compounds completely inhibited the mycelial growth and sporulation of P. italicum. Among these chemicals, ammonium carbonate, ammonium molybdate, sodium phosphate, sodium sulfite, EDTA, sodium metabisulfite and sodium salicylate were, also, fungicidal. Indeed, mycelial Plugs transferred from chemical amended PDA to unamended PDA failed to grow after additional 9 days incubation at 25°C (Table 3).
| Table 2: | In vitro effects of various test compounds on mycelial growth and sporulation of Penicillium italicum |
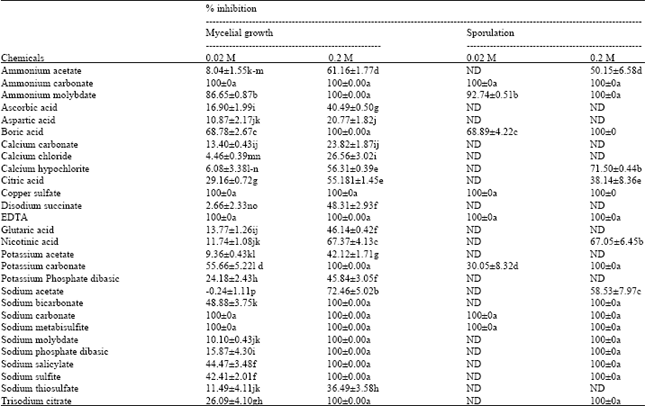 | |
| ND: Not determined. Values expressed are mean of three replicates. Values within each column followed by different letters are significantly different at p<0.05. Means followed by the same letter in each column do not differ significantly according to Duncan multiple range tests at p<0.05 | |
| Table 3: | EC50, Minimum inhibitory concentration (MIC)) and Minimum fungicidal concentration (MFC) values of various compounds tested against P. italicum |
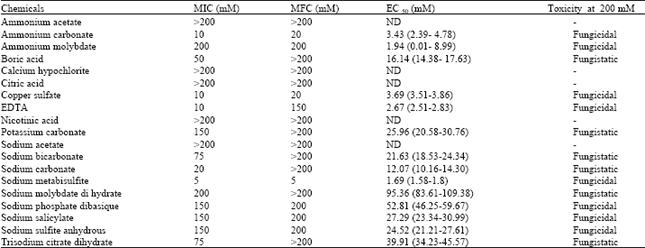 | |
The remaining compounds reduced the pathogen growth; MGI values varied from 20.77% recorded with aspartic acid to 72.46% recorded with sodium acetate.
The obtained results demonstrate that several organic acids and salts can inhibit significantly the growth of P. italicum and reduce or completely inhibit its spore production. This is in agreement with the finding of Mills et al. (2004) and Hervieux et al. (2002) who reported complete inhibition of mycelial growth was generally associated with complete inhibition of sporulation. In this study, copper sulfate strongly inhibits mycelial growth and sporulation of P. italicum. However, when tested against Phytophthora erythroseptica, this compound reduced mycelial growth without inhibiting sporulation (Mills et al., 2004).
Results of MIC and MFC study are shown in Table 3. The lowest MIC and MFC values were recorded for Sodium metabisulfite tested at 5 mM. The second lowest MIC value was recorded for EDTA, ammonium carbonate and copper sulfate treatments (10 mM). However, the MFC of EDTA is widely greater than the one of Copper sulfate and ammonium carbonate; since those salts showed a fungicidal activity at only 20 mM while EDTA at 150 mM. The highest MIC and MFC values were recorded for ammonium acetate, citric acid, sodium acetate, nicotinic acid and calcium hypochlorite.
The EC50 values for tested compounds were further determined (Table 3). The lowest values were obtained for sodium metabisulfite (1.69 mM), ammonium molybdate (1.94 mM), EDTA (2.67 mM), ammonium carbonate (3.43 mM ) and copper sulfate (3.68 mM). The data show that P. italicum has differential sensitivity to bicarbonate and carbonate salts, as demonstrated by its varying rates for complete inhibition of colony growth and sporulation. Sodium carbonate and sodium bicarbonate are common food additives permitted with no restrictions for many applications in North American regulation including organic agriculture (Smilanick et al., 1999). According to their MIC and EC50 values, ammonium and sodium carbonate were more toxic than the other tested carbonic salts. Olivier et al. (1998) reported that sodium carbonate and potassium carbonate were more effective than the respective bicarbonate salts, as sodium and potassium carbonate completely inhibited radial growth of Helminthosporium solani at 0.1 and 0.2 M. This finding is similar to the level of inhibition reported by Hervieux et al. (2002) and five to ten time higher than the level of inhibition observed in this study. The present results are consistent with previous studies that demonstrate better disease control with carbonate salts than with bicarbonate salts (Biggs et al., 1994; Palmer et al., 1997). Palou et al. (2001) demonstrated that sodium carbonate and sodium bicarbonate had fungistatic rather fungicidal activity against P. italicum which is consistent with present data. Whereas, ammonium carbonate exhibited a fungicidal activity at 200 mM. Ammonium salts displayed also an inhibitory effect on P. italicum which concurs with the finding of Hervieux et al. (2002) on H. solani. Palmer et al. (1997) demonstrated that ammonium salts were effective under alkaline conditions where ammonia gas NH3 (additional active compounds) is favored over the ineffective NH4+ form. Montesinos-Herrero et al. (2011) reported that postharvest green mold and blue mold, caused by Penicillium digitatum and Penicillium italicum, respectively, were effectively controlled by fumigation of lemons and oranges with ammonia gas. Sodium metabisulfite, a compound with antimicrobial activity (Hervieux et al., 2002; Mills et al., 2004), has been shown to completely inhibit in vitro mycelial growth and sporulation of H. solani (Hervieux et al., 2002), Fusarium sambucinum (Mecteau et al., 2002) and a wide range of potato postharvest pathogens (Mills et al., 2004) at a concentration of 0.2 M. Present results indicate that Sodium metabisulfite completely inhibited mycelial growth and sporulation of P. italicum at a concentration as low as 0.02 M. At biochemical level, the mode of action of this compound lies in the inhibition of cellular intermediary metabolism, energy production, protein biosynthesis, DNA replication and membranes synthesis (Mills et al., 2004).
Effect of chemicals on spore germination and germ tube elongation: It is evident from the Table 4 that out of 28 salt compounds tested against P. italicum, 27 compounds showed a reduction or complete inhibition of spore germination and germ tube elongation in a dose-dependent manner. In contrast, nicotinic acid allowed spore germination but slightly reduced germ tube elongation. The toxicity of ammonium molybdate, copper sulfate, calcium hypochlorite, sodium metabisulfite and sodium carbonate to spores was higher to that of the other test compounds. Since percent germination in salts amended medium (diluted oranges juice) ranged between 96.27 and 100% at only 2 mM. Results revealed that the compounds which inhibited spore germination also inhibited the mycelial growth of P. italicum. In contrast, calcium hypochlorite affects slightly the mycelial growth of P. italicum at 0.02 M (Table 2) but it strongly inhibited the spore germination and germ tube elongation at low concentration (Table 4). Mills et al. (2004) showed that copper sulfate completely inhibited the spore germination of various potato pathogens. This result agrees with the level of inhibition observed in the present study. While 5 mM concentration of sodium metabisulfite and copper sulfate was required for complete inhibition of spore germination and germ tube elongation, 20 mM concentration was required for complete inhibition of mycelial growth. This confirms that mycelium growth in vitro is less sensitive to these compounds compared with spore germination.
Hwang and Klotz (1938) indicated that the in vitro activity of several carbonate salts against germinated or ungerminated spores of P. italicum was higher than that of bicarbonate salts.
| Table 4: | In vitro effects of various test compounds on spore germination and germ tube elongation of Penicillium italicum |
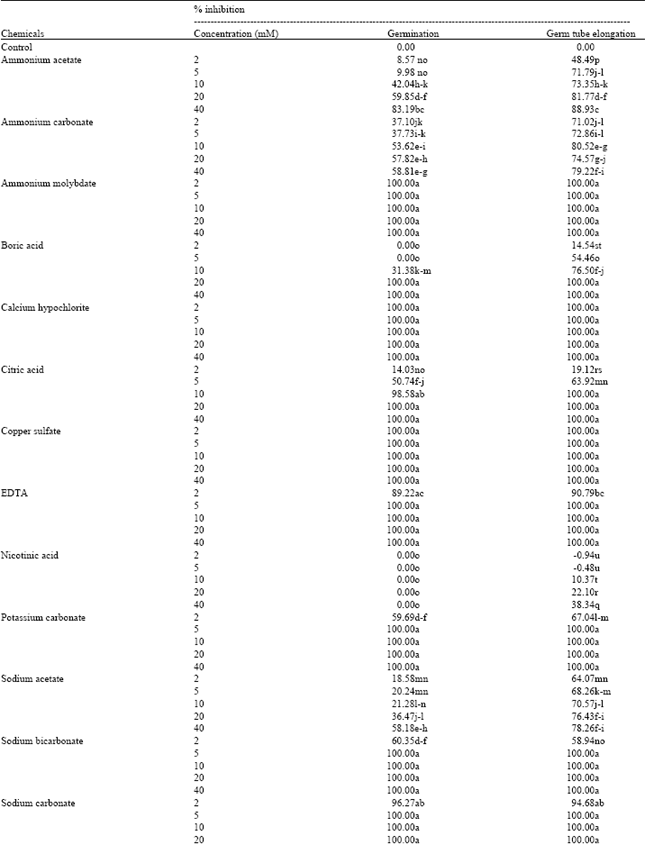 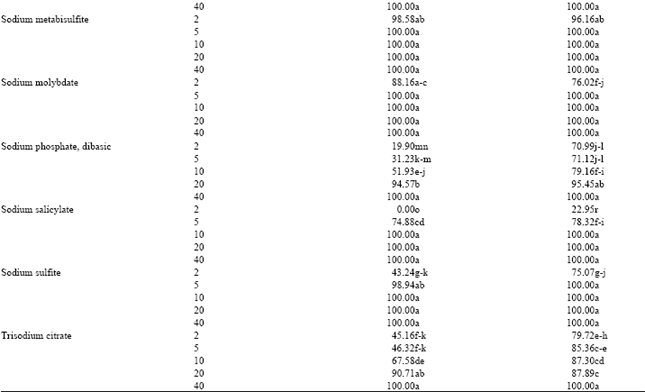 | |
| Germination of at least 100 conidia was assessed microscopically after 18 h incubation in the dark in dilute orange juice. Value was the mean of three replicates. Means followed by the same letter in each column do not differ significantly according to Duncan multiple range tests at p<0.05 | |
In contrast, no significant difference was found in the inhibitory effect between the two salts. The inhibitory effect of salt compounds on spore germination and germ tube elongation of various fungal pathogens have been previously documented (Mecteau et al., 2002; Mills et al., 2004; Arslan et al., 2006). Fallik et al. (1997) reported that the inhibitory effect of sodium bicarbonate on microorganisms may be due to a reduction of cell turgor pressure with collapse and shrinkage of hyphae and spore. However, the mechanisms by which fungi are tolerant or sensitive to salt compounds are poorly understood.
Present results confirmed high antifungal activities of sodium metabisulfite and copper sulfate on various asexual developmental stages of the life cycle of P. italicum in vitro. According to Errampalli (2004), antifungal compounds that significantly inhibits germination of conidia, mycelial growth and subsequently, rot formation should reduce the ability of pathogen to cause disease. As citrus fruit are commonly contaminated by postharvest fungal pathogens during processing, a salt compound with strong effect on spore germination and germ tube elongation is highly desirable to reduce the initial load of spores and subsequent deterioration of fruits. To our knowledge, this is the first report on the in vitro use of sodium metabisulfite and copper sulfate to control P. italicum development.
CONCLUSION
The results of this study suggest that salt compounds may act as an alternative to synthetic fungicides. However, the potential use of organic acid and salts to control blue mold of citrus requires a detailed examination of their biological activity in vivo and the development of formulation which inhibits the growth of the pathogen at non-phytotoxic concentrations.
REFERENCES
- Amiri, A. and G. Bompeix, 2011. Control of Penicillium expansum with potassium phosphite and heat treatment. Crop Prot., 30: 222-227.
CrossRefDirect Link - Arslan, U., K. Ilhan and O.A. Karabulut, 2006. Evaluation of food additives and low-toxicity compounds for the control of bean rust and wheat leaf rust. J. Phytopathol., 154: 534-541.
CrossRef - Arslan, U., K. Ilhan, C. Vardar and O.A. Karabulut, 2009. Evaluation of antifungal activity of food additives against soilborne phytopathogenic fungi. World J. Microbiol. Biotechnol., 25: 537-543.
CrossRef - Barrera-Necha, L.L., S. Bautista-Banos, H.E. Flores-Moctezuma and A.R. Estudillo, 2008. Efficacy of essential oils on the conidial germination, growth of Colletotrichum gloeosporioides (Penz.) penz. and sacc and control of postharvest diseases in papaya (Carica papaya L.). Plant Pathol. J., 7: 174-178.
CrossRefDirect Link - Barrera-Necha, L.L., C. Garduno-Pizana and L.J. Garcia-Barrera, 2009. In vitro antifungal activity of essential oils and their compounds on mycelial growth of Fusarium oxysporum f. sp. gladioli (Massey) snyder and hansen. Pak. J. Nutr., 8: 17-21.
CrossRefDirect Link - Biggs, A.R., M.M. El-Kholi and S.M. El-Neshawy, 1994. Effect of calcium salts on growth, pectic enzyme activity and colonization of peach twigs by Leucostoma persoonii. Plant Dis., 78: 886-890.
Direct Link - Boubaker, H., B. Saadi, E.H. Boudyach and A. Ait Benaoumar, 2009. Sensitivity of Penicillium digitatum and P. italicum to imazalil and thianbendazole in morocco. Plant Pathol. J., 8: 152-158.
Direct Link - Corral, L.G., L.S. Post and T.J. Montville, 1988. Antimicrobial activity of sodium bicarbonate. J. Food Sci., 53: 981-982.
CrossRef - Droby, S., M. Wisniewski, A. El-Ghaouth and C. Wilson, 2003. Influence of food additives on the control of postharvest rots of apple and peach and efficacy of the yeast-based biocontrol product Aspire. Postharvest Biol. Technol., 27: 127-135.
CrossRefDirect Link - Eckert, J.W. and I.L. Eaks, 1989. Postharvest Disorders and Diseases of Citrus Fruits. In: The Citrus Industry: Crop Protection, Postharvest Technology and Early History of Citrus Research in California, Calavan, E.C. and G.E. Carman (Eds.). Vol. 5, University of California Press, Berkeley, pp: 179-269.
- El-Ghaouth, A., J.L. Smilanick and C.L. Wilson, 2000. Enhancement of the performance of Candidasaitoana by the addition of glycolchitosan for the control of postharvest decay of apple and citrus fruit. Postharvest Biol. Technol., 19: 103-110.
CrossRefDirect Link - Elkhamass, M., B. Oulahcen, A. Lekchiri, A. Sebbata and Y. Charhabaili, 1994. Strategies used to Control Postharvest Diseases of Citrus Fruit. In: Postharvest Pathology and Technology for Horticultural Commodities: Recent Advances, Ait Oubahou, A. and M. El-otmani (Eds.). Institut Agronomique et Veterinaire Hassan II, Agadir, Maroc, pp: 388-398.
- Errampalli, D., 2004. Effect of fludioxonil on germination and growth of Penicillium expansum and decay in apple cvs. Empire and Gala. Crop Prot., 23: 811-817.
CrossRefDirect Link - Fallik, E., O. Ziv, S. Grinberg, S. Alkalai and J.D. Klein, 1997. Bicarbonate solutions control powdery mildew (Leveillula taurica) on sweet red pepper and reduce the development of postharvest fruit rotting. Phytoparasitica, 25: 41-43.
CrossRef - Gabler, F.M. and J.L. Smilanick, 2001. Postharvest control of table grape gray mold on detached berries with carbonate and bicarbonate salts and disinfectants. Am. J. Enol. Viticult., 52: 12-20.
Direct Link - Holmes, G.J. and J.W. Eckert, 1995. Relative fitness of imazalil-resistant and sensitive biotypes of Penicillium digitatum. Plant Dis., 79: 1068-1073.
Direct Link - Holmes, G.J. and J.W. Eckert, 1999. Sensitivity of Penicillium digitatum and P. italicum to postharvest citrus fungicides in Californa. Phytopathology, 89: 716-721.
CrossRef - Kazemi, M., M. Aran and S. Zamani, 2011. Effect of calcium chloride and salicylic acid treatments on quality characteristics of kiwifruit (Actinidia deliciosa cv. Hayward) during storage. Am. J. Plant Physiol., 6: 183-189.
CrossRefDirect Link - Hervieux, V., E.S. Yaganza, J. Arul and R.J. Tweddell, 2002. Effect of organic and inorganic salts on the development of Helminthosporium solani, the causal agent of potato silver scurf. Plant Dis., 86: 1014-1018.
CrossRefDirect Link - Mills, A.A.S., H.W. Plat and R.A.R. Hurta, 2006. Sensitivity of Erwinia spp. to salt compounds in vitro and their effect on the development of soft rot in potato tubers in storage. Post Harvest Biol. Technol., 41: 208-214.
CrossRef - Montesinos-Herrero, C., J.L. Smilanick, J.S. Tebbets, S. Walse and L. Palou, 2011. Control of citrus postharvest decay by ammonia gas fumigation and its influence on the efficacy of the fungicide imazalil. Posth. Biol. Technol., 59: 85-93.
CrossRef - Nigro, F., L. Schena, A. Ligorio, I. Pentimone, A. Ippolito and M.G. Salerno, 2006. Control of table grape storage rots by pre-harvest applications of salts. Postharvest Biol. Technol., 42: 142-149.
CrossRefDirect Link - Olivier, C., D.E. Halseth, E.S.G. Mizubuti and R. Loria, 1998. Postharvest application of organic and inorganic salts for suppression of silver scurf on potato tubers. Plant Dis., 82: 213-217.
CrossRefDirect Link - Palmer, C.L., R.K. Horst and R.W. Langhans, 1997. Use of bicarbonates to inhibit in vitro colony growth of Botrytis cinerea. Plant Dis., 81: 1432-1438.
CrossRefDirect Link - Palou, L., J.L. Smilanick and C.H. Crisosto, 2009. Evaluation of food additives as alternative or complementary chemicals to conventional fungicides for the control of major postharvest diseases of stone fruit. J. Food Prot., 174: 1037-1046.
PubMedDirect Link - Palou, L., J.L. Smilanick and S. Droby, 2008. Alternatives to conventional fungicides for the control of citrus postharvest green and blue moulds. Stewart Postharvest Rev., 4: 1-16.
Direct Link - Palou, L., J.L. Smilanick, J. Usall and I. Vinas, 2001. Control of postharvest blue and green molds of oranges by hot water, sodium carbonate and sodium bicarbonate. Plant Dis., 85: 371-376.
Direct Link - Punja, Z.K. and M.M. Gaye, 1993. Influence of postharvest handling practices and dip treatments on development of black root rot on fresh market carrots. Plant Dis., 77: 989-995.
Direct Link - Rani, P.U. and P. Devanand, 2011. Efficiency of different plant foliar extracts on grain protection and seed germination in maize. Res. J. Seed Sci., 4: 1-14.
CrossRefDirect Link - Shirzadeh, E. and M. Kazemi, 2011. Effect of malic acid and calcium treatments on quality characteristics of apple fruits during storage. Am. J. Plant Physiol., 6: 176-182.
CrossRefDirect Link - Smilanick, J.L., M. Mansour, G.G. Mlikota and W.R. Goowdwine, 2006. The effectiveness of pyrimethanil to inhibit germination of spores of Penicillium digitatum and to control citrus green mold after harvest. Postharv. Biol. Technol., 42: 75-85.
Direct Link - Smilanick, J.L., M.F. Mansour, F.M. Gabler and D. Sorenson, 2008. Control of citrus postharvest green mold and sour rot by potassium sorbate combined with heat and fungicides. Postharvest Biol. Technol., 47: 226-238.
CrossRefDirect Link - Smilanick, J.L., M.F. Mansour, D.A. Margosan, F. Mlikota-Gabler and W.R. Goodwine, 2005. Influence of pH and NaHCO3 on the effectiveness of imazalil to inhibit germination of spores of Penicillium digitatum and to control green mold on citrus fruit. Plant Dis., 89: 640-648.
Direct Link - Smilanick, J.L., D.A. Margosan, F. Mlikota, J. Usall and I.F. Michael, 1999. Control of citrus green mold by carbonate and bicarbonate salts and the influence of commercial postharvest practices on their efficacy. Plant Dis., 83: 139-145.
CrossRefDirect Link - Smilanick, J.L. and D. Sorenson, 2001. Control of postharvest decay of citrus fruit with calcium polysulfide. Postharvest Biol. Technol., 21: 157-168.
Direct Link - Taqarort, N., A. Echairi, R. Chaussod, R. Nouaim and H. Boubaker et al., 2008. Screening and identification of epiphytic yeasts with potential for biological control of green mold of citrus fruits. World J. Microbiol. Biotechnol., 24: 3031-3038.
CrossRef - Tripathi, P. and N.K. Dubey, 2004. Exploitation of natural products as an alternative strategy to control postharvest fungal rotting of fruit and vegetables. Postharvest Biol. Technol., 32: 235-245.
CrossRefDirect Link - Valencia-Chamorro, S.A., L. Palou, M.A. del Rio and M.B. Perez-Gago, 2008. Inhibition of Penicillium digitatum and Penicillium italicum by hydroxypropyl methylcellulose-lipid edible composite films containing food additives with antifungal properties. J. Agric. Food Chem., 56: 11270-11278.
CrossRefPubMedDirect Link - Zhu, J.W., Q.Y. Xie and H. Li, 2006. Occurrence of imazalil-resistant biotype of Penicillium digitatum in China and the resistant molecular mechanism. J. Zhejiang Univ. Sci., 7: 362-365.
CrossRefDirect Link - Mecteau, M.R., J. Arul and R.J. Tweddell, 2002. Effect of organic and inorganic salts on the growth and development of Fusarium sambucinum, a causal agent of potato dry rot. Mycol. Res., 106: 688-696.
CrossRefDirect Link - Mills, A.A.S., H.W. Platt and R.A.R. Hurta, 2004. Effect of salt compounds on mycelial growth, sporulation and spore germination of various potato pathogens. Postharvest Biol. Technol., 34: 341-350.
CrossRefDirect Link