
C. Simo
Laboratory of Plant Physiology, Department of Biological Sciences, Higher Teacher`s Training College, P.O. Box 47, Yaounde, Cameroon
P.F. Djocgoue
Laboratory of Plant Physiology, Department of Biological Sciences, Higher Teacher`s Training College, P.O. Box 47, Yaounde, Cameroon
H.D. Mbouobda
Laboratory of Plant Physiology, Department of Biological Sciences, Higher Teacher`s Training College, P.O. Box 47, Yaounde, Cameroon
P.O. EFFA
Laboratory of Plant Physiology, Department of Biological Sciences, Higher Teacher`s Training College, P.O. Box 47, Yaounde, Cameroon
T. Boudjeko
Department of Biochemistry, Faculty of Science, P.O. Box 812, Yaounde, Cameroon
D.N. Omokolo
Laboratory of Plant Physiology, Department of Biological Sciences, Higher Teacher`s Training College, P.O. Box 47, Yaounde, Cameroon
Plant Pathology Journal
Year: 2011 | Volume: 10 | Issue: 3 | Page No.: 89-98







ABSTRACT
The black pod disease caused by P. megakarya is responsible for about 30% of cocoa production loss in Cameroon and Polyphenoloxidase (PPO), which contribute to plant resistance, were analysed in response to stress due to cocoa pod inoculation by mycelium of Phytophthora megakarya. Two cocoa plant (Theobroma cacao) hybrid families (F79: ♀T79/467 x ♂ SNK13; F13: ♀ SNK13x ♂ T79/467) were used in this study. Quantitative and qualitative analyses of the specific activities of PPO in the healthy, scarified and inoculated pods were realised in function of the productivity and the necrosis surface. Finally the study of the resistance transmission of PPO vis-à-vis P. megakarya from parents to offsprings was also performed. In the ionically bounded (I) fractions, the increase in the activities of PPO is correlated with tolerance rate of genotypes within the two hybrid families. Heritability of the reciprocal crossing of the PPO activities is higher for the (I) fractions (0.88 for F79 and 0.83 for F13) and do not show any significant difference for the two families suggesting the absence of a maternal effect. The use of PPO as a biochemical marker has permitted to reveal that the bonded PPO are more heritable than the soluble ones and these bonded PPO are seemingly good indicators of the resistance. Over all, the present study has enabled to classify the different groups of genotypes as tolerant and less tolerant and so, a large scale distribution of the tolerant and productive genotypes to cocoa farmers will be beneficial.
PDF Abstract XML References Citation
Received: July 07, 2011;
Accepted: September 21, 2011;
Published: November 15, 2011
How to cite this article
C. Simo, P.F. Djocgoue, H.D. Mbouobda, P.O. EFFA, T. Boudjeko and D.N. Omokolo, 2011. Variation and Heritability of Polyphenoloxidasic Activities in Two Hybrid Families of Theobroma cacao L. after Cocoa Pods Inoculation with Phytophtora megakarya Bras. et Grif. Plant Pathology Journal, 10: 89-98.
DOI: 10.3923/ppj.2011.89.98
URL: https://scialert.net/abstract/?doi=ppj.2011.89.98
DOI: 10.3923/ppj.2011.89.98
URL: https://scialert.net/abstract/?doi=ppj.2011.89.98
INTRODUCTION
The cocoa plant (Theobroma cacao) is a humid tropical forest crop, originated from East and Central America (Wood and Lass, 2001; Motamayor et al., 2002; Niemenak et al., 2010). It is a cash crop for most tropical countries such as Brazil, Ghana, Ivory Coast and Cameroon. The mean annual world production of cocoa is 10.6 billion dollars (ICCO, 2010). The plant is a source of revenue for most Cameroon farmers. However, black pod disease caused by Phytophthora megakarya which is one of the most aggressive pathogenic agents of this plant, is responsible for high losses in the whole world. Those losses generally range from 20 to 30% but might attain 80% in some Central African Countries (Pokou et al., 2008). The losses can reach 100% of the total annual production if no proper measures of control are taken (Ndoumbe-Nkeng et al., 2004). Even if the chemical fight against black pod disease is effective, it still remains difficult, expensive and is a short term process for management of cocoa plantations. The use of tolerant and productive genotypes is cheaper for the farmer and does not pose any ecological problems related to the use of pesticides.
Studies dedicated to the observation of cocoa pods infection by P. megakarya in the farm reveal the existence of genetically heritable partial of tolerance (Djocgoue et al., 2006). The field observation of the genotypes performances under natural or artificial infection conditions and leaf or pod inoculation has enabled the identification of tolerant genotypes (Nyasse et al., 2002; Djocgoue et al., 2006). The world cocoa production comes mainly from hybrid genotypes. In effect, the highest world producers of cocoa are those that use the highest hybrid rate in the regeneration of plots. This rate is 76% for Indonesia, 69.2% for Ivory Coast, 96% for Malaysia, 63.2% for Ghana, 25.1% for Cameroon and 17.4% for Ecuador. Those hybrid varieties have high production and environmental adaptation capacities due to the additive effect of parent genes and can be used on a large scale (Tahi et al., 2000; Djocgoue et al., 2006; Boudjeko et al., 2007).
One of the priority objectives for cocoa farming in Cameroon is the selection of genotypes which are less vulnerable to black pod disease. Nevertheless, the rate of cocoa pods attack ranges from one genotype to another. Nyasse et al. (2002) have proven the existence of a correlation between cocoa resistance vis-à-vis to P. megakarya and the development of the lesion on cocoa pods obtained through artificial inoculation of the pathogenic agent by the lesion surrounded by living cells which have become resistant to microorganisms. Those cells have their cell walls reinforced by a layer of cellulose, lignin and other phenolic compounds (Housti et al., 2002; Majourhat et al., 2002). Numerous enzymes are activated notably Phenylalanine Ammonialyase (PAL), Peroxidases (Pox) and polyphenoloxidases (PPO); numerous biochemical pathways are stimulated and secondary metabolism increases. All these reactions constitute the hypersensitive response. It occurs after the regeneration of the pathogenic agent in the resistant plant. The PPO (EC.1.14.18.1) are actively involved in the defence mechanism of the plant against microbial and viral infections as well as unfavourable climatic conditions (Constabel et al., 2000; Housti et al., 2002; Wititsuwannakul et al., 2002). According to Omokolo et al. (2003), a sensitive increase in PPO activities simultaneously with lesion formation infection hypersensitive pods of P. megakarya was observed. The increase in this activity portrays the action of PPO in the oxidation of phenols for the production of quinones constituting therefore a molecular base for the resistance acquired by the plant in response to elicitines and proteinic signals emitted by the pathogenic agent (Herger and Klinghauf, 1990; Schneider and Ullrich, 1994). The PPO contributes to plant resistance by producing hydroxyphenols and quinones which are bactericide and fungicide compounds capable of undergoing polymerization. Moreover, they are involved in the oxidative detoxification of phytotoxins produced by pathogens (Vaughn et al., 1988). The PPO catalyse the oxidation of phenols to O-quinones which react with amino acids and proteins. The latter, after polymerization, give rise to complex compounds which are responsible for the black pods in tissues (Colak et al., 2005; Schweiggert et al., 2005).
In the present study, the assessment of heritability of polyphenoloxidasic activities in the resistance of T. cacao vis-à-vis P. megakarya was done; the quantitative and qualitative analyses of the specific activities of PPO in the healthy, scarified and inoculated pods was also realised in function of the productivity and the necrosis surface; finally the study of the resistance transmission of PPO vis-à-vis P. megakarya from parents to offsprings was also performed. The results obtained enabled to classify the different groups of genotypes as tolerant and less tolerant.
MATERIALS AND METHODS
The present investigation was carried out in the field laboratory of the Department of Biological Sciences, High Teacher’s Training College, University of Yaounde I in Cameroon during the period from May 2007 to October 2009.
Plant material: The plant material derived from the experimental station of the Cocoa Development Corporation (SODECAO) is constituted by 3-month-old cocoa pods that belong to two parental clones (the sensitive and productive SNK13, Trinitario group, a tolerant and less productive T79/467, Forastero group) as well as their hybrid progenies organized within the F13 (♀SNK13x♂T79/467) and F79 (♀T79/467x♂SNK13) populations. The fungal material is a local isolate of P. megakarya (TA121) obtained from the Research Institute for Agricultural Development (IRAD) of Nkolbisson (Yaounde, Cameroon).
Cocoa pods inoculation: The apparently healthy pods were harvested, washed with tap water, sterilized with 70° alcohol and divided in three groups. The first group consisted of healthy pod, second group of pods scarified and inoculated with sterilized agar disk, third group of pods scarified and inoculated with an agar disk containing P. megakarya mycelium obtained from 7-day-old PDA culture medium and incubated at 25-26°C in the dark in a humid chamber. The measurement of the necrosis surface area was done in 3, 4, 5 and 6 days after inoculation. The diameters of the more or less circular necrotic spots were measured and the surfaces calculated using Blaha and Lotode (1976) formula.
Estimation of the heritability: For the different parameters measured, heritability has been estimated according to Falconer (1974). This estimation was done by the regression slope between means of parents and means of progeny.
Genotypic identification for biochemical analysis: The parental and hybrid genotypes were identified for biochemical studies such as tolerant and productive (F1307, F1314, F7902, F7928), tolerant and less productive (F1315, F1313, F7926, F7907), less tolerant and productive (F1324, F1308, F7915, F7919) and less tolerant and less productive (F1321, F1326, F7904, F7911) (Djocgoue et al., 2010).
Preparation of crude enzyme extracts: At day 6 after inoculation, cortex samples were collected at 1 cm beyond the necrosis for biochemical analysis. Samples from pods of the same genotype and of the same treatment were combined and stored at -20°C until analysis.
For extract soluble PPO, 50 mM Tris-maleate buffer pH 7 containing 0.5 M mannitol was added to the original powder in the proportion 3:1 (v:w), stirred for 2 min and incubated at 4°C for 1 h and then centrifuged at 6,000 g for 30 min. The supernatant containing soluble PPO was collected and kept at -20°C until measurements. Ionically-bound PPO fraction was obtained by mixing the residue in 10 mM Tris-maleate buffer pH 7 containing 1 M NaCl. The homogenate was incubated at 4°C for 1 h and then centrifuged at 6,000 g for 30 min (Mbouobda et al., 2010).
Enzyme assay and protein determination: Enzyme activities were analysed in both soluble and ionically-bound fractions since their relative activities depend on the physiological state of the cells.
Assay of PPO was carried out using catechine as substrate. The standard assay contained 70 μL of the enzyme extract and 3 mL of 0.66 M phosphate buffer pH 7. The mixture was transferred quickly in a cuvette after adding 0.3 mL catechine 10 mM and the increase in absorbance at 330 nm over a 30 s period was determined using a spectrophotometer (U-2000 Hitachi). Enzyme activity is expressed as change in absorbance: Absorbance 30 s-1 g-1 fresh weight.
Proteins contents were estimated according to Bradford (1976). All the concentrations indicated are final concentrations in assay media.
Specific activity was estimated by the ratio of enzyme total activity over quantity of total protein.
PAGE gel staining: Isopolyphenoloxidases were analysed by PAGE under non-reducing conditions. Acrylamide (12 and 5%) was used for the resolving and stacking gel, respectively. Basic gel buffer was 0.4 M Tris pH 8.8 and the electrode buffer 0.03 M Tris, 0.2 M glycine pH 8.6. Samples contained about 20 μL proteins and 10% (v:v) glycerol. The gel was run at 4±1°C at a constant voltage (300 V) for 10 h and then stained in 100 mM sodium acetate buffer pH 5, 10 mM catechine and 5 mM DL DOPA for 30 min (Mbouobda et al., 2010).
Statistical analysis: Data from this study are presented in the form of Means±SD, for at least three independent experiments during three successive trips. Three measurements were recorded for each campaign. Analysis of Variance (ANOVA) and Duncan test were used to compare the susceptibility levels and polyphenoloxidasic activities of better progenies resulting from different crosses to access hybrid vigour, using SPSS 12 version for Windows. The p-value less than 0.05 was considered significant. Hierarchical classification were performed with SPAD version 4.1 windows software.
RESULTS
Soluble PPO
Variation in specific activities of the soluble PPO: The specific activities of soluble PPO in the healthy pods is high in the F1314 hybrids (108.42±1.70 IU mg-1 of protein), F7902 (111.08±2.94 IU mg-1 of protein) and F7907 (108.36±7.70 IU mg-1 of protein) (Table 1). Under lesion condition, the specific activity of the soluble PPO increases in almost all the genotypes where it varies from 20 to 480%, except the F1314 hybrid where it decreases of 5% in the F13 (Table 1). In the F79 family, it also increases in almost all the genotypes varying from 10 to 48% except few genotypes (F7926, F7907 and F7902) which indicates a decrease from 2% to 10% (Table 1). Under inoculation condition the specific activities of the soluble PPO in the F13 increases in almost all the hybrids and ranges from 4 to 438% (Table 1). Instead, in the F79 family, it increases in all the genotypes and varies from 11 to 135% (Table 1). At 95% of the homogeneity, the direct hierarchical classification of the genotypes obtained from the soluble PPO specific activities of the healthy, scarified and inoculated pods has enabled to distinguish four groups for each of the offsprings.
| Table 1: | Variation of the specific activity of soluble PPO (IU mg-1 of protein) in the mesocarp of parental and hybrid cortex |
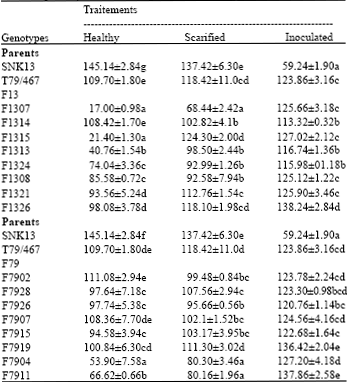 | |
| Values following by the same letter within column are not significantly different (p<0.05) | |
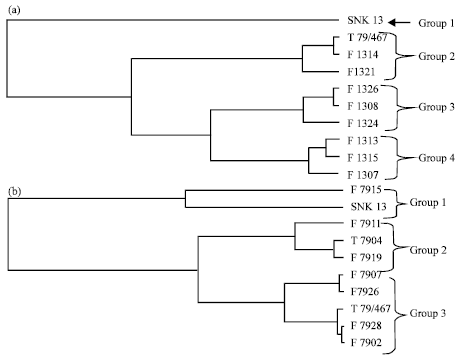
The first group of F13 offsprings is constituted by the parent SNK13. The second group is constituted by the T79/467, F1321, F1326 and F1314 which have a high soluble PPO specific activity. The third group is made up of F1324, F1315 and F1313 with an average specific activity of soluble PPO (Fig. 1a). In the F79 offsprings, the first group is composed of the SNK13 genotype. The second one is constituted by F7904 and F7911. The third group is made up of F7915, T79/467 and F7919 genotypes having a high soluble PPO activity. The fourth group is constituted by F7928, F7907 and F7902 having a high soluble PPO specific activity (Fig. 1b).
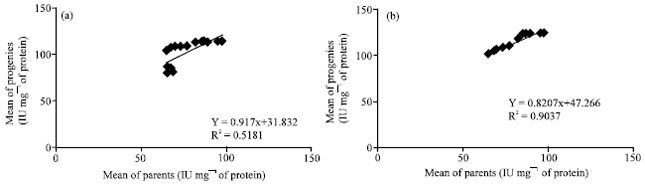
Heritability of specific activities of the soluble PPO: Heritability proper (h2) is estimated by the slope of the regression between the means and the performances of the parental genotypes and those of the progenies due to the specific activity of the soluble PPO (Fig. 2). The heritability value obtained in the F79 (♀T79/467x♂SNK13) is similar to that obtained in the F13 family (♀SNK13x♂T79/467). Those values are respectively 0.60 and 0.52. There is a strong correlation between the specific activities of the soluble PPO of the parents and those of the progenies (Fig. 2).
| Table 2: | Variation of the specific activity of bonded PPO (IU mg-1 of protein) in the mesocarp of parental and hybrid cortex |
 | |
| Values following by the same letter within column are not significantly different (p<0.05) | |
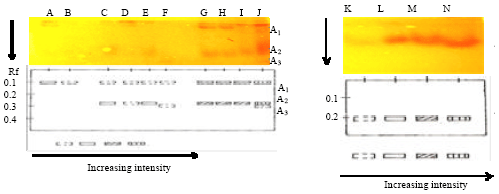
Analysis of soluble PPO profiles: The electrophoretic profiles of the soluble fraction presents one PPO isoform of the frontal ratio (Rf = 0.24) in the parental genotypes (SNK13 and T79/467) and in the F13 and F79 families under healthy, scarified and inoculated conditions (Fig. 3).
Bonded PPO
Variation in specific activities of the bonded PPO: The specific activity of the bonded PPO in the healthy pods is high in the F1307 (75.84±1.88 IU mg-1 of protein), F1315 (71.12±5.18 IU mg-1 of protein), F1326 (72.34±4.50 IU mg-1 of protein), F7915 (128.59±6.68 IU mg-1 of protein) and F7904 (70.04 ±2.18 IU mg-1 of protein) (Table 2). Under scarified condition, the specific activity of the bonded PPO increases almost in all the genotypes ranging from 4% to 86% in the F13 family (Table 2). On the contrary in the F79 family, it increases in all the hybrid genotypes and this increase ranges from 3% to 180%. Under inoculation condition in the F13 family, the specific activity of the bonded PPO increases in all the hybrids fluctuating between 24% and 156%. In the F79 family, the specific activity also increases in practically all the hybrid genotypes, except that of the F7915 where it drops by 16% (Table 2).
At 95% of homogeneity, the direct hierarchical classification of the genotypes obtained from the specific activities of the bonded PPO of healthy, scarified and inoculated pods enabled to respectively differentiate four and three groups for the F13 and F79 families.
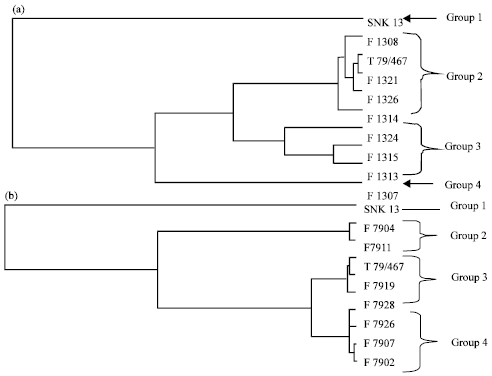 | |
| Fig. 1(a-b): | Direct hierarchical classification of genotypes of the F13 (a) and the F79 family (b) in relation to the specific activities of the soluble PPO |
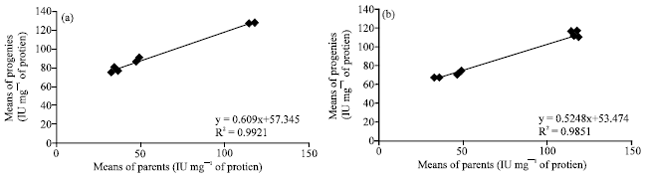 | |
| Fig. 2(a-b): | Estimation of the heritability proper (h2) of the soluble PPO activities in the (a) F13 family and (b) in the F79 family |
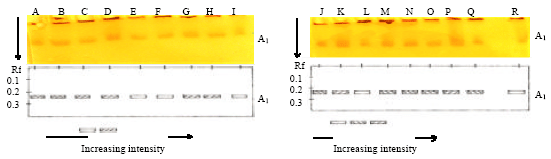 | |
| Fig. 3(a-b): | Electrophoretic profiles of soluble PPO in the cortex of the parental and hybrid pods submitted to different treatments (healthy, scarified and inoculated). A: Healthy SNK13; B: Scarified SNK13; C: Inoculated SNK13; D : Healthy T79/467; E: Scarified T79/467; F: Inoculated T79/467; G: Healthy F1321; H: Scarified F1321; I: Inoculated F1321. J: Healthy F1324; K: Scarified F1324; L: Inoculated F1324; M: Inoculated F1307; N: Inoculated F1315; O: Inoculated F1319; P: Inoculated F7902; Q: Inoculated F7911; R: Inoculated F7926 |
 | |
| Fig. 4(a-b): | Direct hierarchical classification of genotypes of the F13 family (a) and the F79 family (b) with respect to the specific activities of the bonded PPO |
 | |
| Fig. 5(a-b): | Estimation of the heritability proper (h2) of the specific activities of bonded PPO in the F13 family (a) and in the F79 family (b) |
The first group of the F13 family is constituted by the SNK13 parent. The second one is constituted by the T79/467 parent, F1314 and F1321 genotypes which have an average specific activity of bonded PPO. The third group is constituted by F1326 and F1324 genotypes which have a low specific activity of bonded PPO. Finally, the fourth group is constituted by F1313, F1315 and F1307 genotypes which have a high specific activity of bonded PPO (Fig. 4a). In the F79 family, the first group is constituted by the SNK13 and F7915 hybrid having an average specific activity of bonded PPO. The second group encloses the F7911 and F7904 genotypes which have a low specific activity of bonded PPO. The third group includes the T79/467 parent and the F7907, F7926, F7928 and F7902 having a high specific activity of bonded PPO (Fig. 4b).
Heritability of specific activities of the bonded PPO: The heritability proper (h2) is estimated by the slope of the regression between the means of the performances of parental genotypes and those of offsprings due to the specific activity of the bonded PPO (Fig. 5). The heritability value obtained in the F79 family (♀T79/467x♂SNK13) is similar to that obtained in the F13 family (♀SNK13x♂T79/467). Those values are respectively 0.91 and 0.82. There is a strong correlation between the specific activities of the bonded PPO of parents and those of the progenies of the F13 family. This correlation is stronger between the specific activities of the bonded PPO of parents and those of progenies of the F79 family (Fig. 5).
 | |
| Fig. 6 (a-b): | Electrophoretic profiles of bonded PPO in the cortex of parental and hybrid pods at the different treatments (healthy, scarified and inoculated). A: Inoculated F1324; B: Inoculated F1321; C: Healthy F1315; D: Scarified F1315, E: Inoculated F1315; F: Inoculated SNK13; G: Inoculated T79/467; H: Scarified T79/467; I: Healthy T79/467; J: Inoculated F1307. K: Inoculated F7911; L: Inoculated F7926; M: Inoculated F7919; N: Inoculated F7902 |
Analysis of bonded PPO profiles: The electrophoretic profile of the bonded fraction shows three PPO isoforms in the F13 family and one in the F79 family. The parental clone SNK13, sensitive and productive under inoculation reveals two isoforms of frontal ratio (Rf = 0.11 et 0.31). In the parental clone T79/467, tolerant and less productive under healthy, scarified and inoculated conditions, two isoforms of frontal ratio (Rf = 0.11 and 0.28) are respectively observed. In the F13 family, the F1307 genotype more tolerant and more productive presents three isoforms of frontal ratio (Rf = 0.11; 0.28 and 0.31) under conditions of inoculation. On the contrary, in the F1315 genotype less tolerant and more productive under healthy, scarified and inoculation conditions, there are two respective observables isoforms of frontal ratio (Rf = 0.11 and 0.28). In the F1324 genotype less tolerant and more productive under inoculation condition, there is one isoform (Rf = 0.11). In the F1321 less tolerant and less productive under inoculation condition, there is also one isoform (Rf = 0.11). In the F79 family under inoculation condition one isoform of frontal ratio (Rf = 0.21; 0.21; 0.21 and 0.21) for the F7902 genotype more productive and more tolerant, F7926 more tolerant and less productive, F7919 less tolerant and less productive and F7911 less tolerant and less productive was observed, respectively (Fig. 6).
DISCUSSION
Pathogens infections are a major limiting factor in plant production and fungi are known as the most causal pathogens on plants and particularly on cocoa plant (Ammar and El-Naggar, 2011; Djocgoue et al., 2010); Valizadeh et al., 2011; Moradi and Jafarpour, 2011; Moradi, 2011). The identification of tolerant and productive cocoa genotypes is a crucial step towards achieving the objective of the economic control. Tolerance in this case can be defined as the ability of a genotype to produce a good crop even when it is infected with a pathogen. Nevertheless, different sources of resistance are recommended for enhanced the durability of resistance (Ochola and Tusiime, 2011; Aliyu and Balogun, 2011). The main goal of the present study was to analyse the heritability of the soluble and bonded PPO in the cortex of healthy, scarified and inoculated pods of parental clones SNK13 and T79/467 and hybrids resulting from the reciprocal crossing of ♀SNK13x♂T79/467 in function of the productivity and the resistance of cocoa pod vis-à-vis P. megakarya.
The main results show that among the healthy plants, the parental clones and 75% of the hybrid genotypes have specific activities of the high PPO with respect to those in the F1315, F1313, F7904 and F7911 genotypes in the soluble fractions (S). Those activities are high in the genotypes of the within the bonded fractions (I). There is a remarkable increase in those activities under conditions of scarification. However, this increase is more significant for a majority of the genotypes under conditions of inoculation in the soluble and bonded fractions. In the (S) fractions, this increase is more pronounce in the F13 family where one notes a 638% increase. On the contrary in the (I) fractions it is more important in the F79 family where one observes a 241% increase. Nevertheless, there is no correlation between the increase rate and PPO activities, the productivity and tolerance of the genotypes in the (S) fraction, contrary to those of the (I) fractions where an increase in PPO activity is correlated with the degree of tolerance of the genotypes within the two hybrid families. Those results are in agreement with those published by Kim et al. (2001), Constabel et al. (2000), Wititsuwannakul et al. (2002), Omokolo et al. (2003) and Dogbo et al. (2007, 2008) who measured the activity of PPO after lesion and/or elicitation. They revealed that the PPO was implicated in numerous functions, among which the defence mechanism of plants against pathogenic agents.
The parental clone T79/467 and all the hybrid genotypes were strongly stimulated by the P. megakarya in the (S) and (I) fractions apart from the parental clone SNK13 and the hybrid genotype F7919 in the (I) fraction. The PPO activity after the inoculation of the plants with P. megakarya proves their implication in the defence reactions of the plants. The works of some authors agree with this hypothesis. In effect, abiotic and biotic effect can activate new proteins as chitinases and other pathogenesis-related proteins and activate many of enzymes which have a role of disease resistance (El-Hersh et al., 2011). The activity of the PPO assessed in the cocoa pods at different treatment conditions has enabled to confirm the increase in this activity under lesion conditions and a more significant increase under conditions of inoculation in practically all of the genotypes. The more tolerant and more productive hybrid genotypes (F1307, F1314, F7902, F7928) and the more tolerant genotypes (F1315, F1313, F7926, F7907) with respect to the best parent in the bonded fraction accumulate a greater quantity of PPO activity in the (I) fraction with small necrotic lesion surfaces thus implication of PPO in the resistance of the plant. Those results are in conformity with those reported by Chen et al. (2000).
As concerns heritability, the PPO activity values obtained in the (S) fraction of the F13 (♀SNK13x♂T79/467) and in the F79 family (♀T79/467x♂SNK13) are relatively low. They are respectively 0.6 and 0.52. On the contrary, although quite similar in both families, those values are high in the (I) fractions and are respectively 0.91 and 0.82 in the F13 and the F79. Those high values in the (I) fractions are evidence of a strong additive variance in the transmission of resistance character to black pod disease. Those results are contrary to those documented by Djocgoue et al. (2007) in the same plant. The manifestation of hybrid vigour and PPO heritability values which have been inherited by the offsprings is an indication of a good general aptitude to parental clone combination and suggest that these results may be under the control of additive gene action. This agrees with the report of Denton and Nwangburuka (2011) and Parthiban et al. (2011) who showed that the characters of plant height at maturity, total weight of fruits per plan are under the influence of additive gene effect. Whatsoever the studied fraction, the absence of a significant difference between the heritability values of reciprocal crossing portrays the absence of parental heritability. This observation suggests that heritability of the PPO is nuclear and non-cytoplasmic. Such observations have already been reported by Djocgoue et al. (2007) studying phenolic compounds on T. cocoa leaves.
The results in the present work generally reveal that the PPO are present both in the healthy pods and the treated ones; they are thus constitutive and have no significant correlations neither with the sensitive P. megakarya nor with the productivity. This suggests a regulation of the expression of this enzyme during the development of the pod. These results are in agreement with those published by Yedidia et al. (2000) after the inoculation of cucumber roots with Trichoderma harzynum; Kasran et al. (2000) after inoculation of T. cacao with Onchobasidium theobromae and Omokolo et al. (2003) after the inoculation of T. cocoa by P. megakarya.
The PPO electrophoretic profiles show that the increase in their activity after inoculation is due not only to the synthesis of new isoforms but mainly to the activation of pre-existing forms. The isoforms (F1307) and (T79/467) of the soluble fractions might be directly related to the resistance. As concerns the other hybrid genotypes in the soluble and bonded fractions, the increase in activity of the PPO can be attributed to an activation of pre-existing forms. The increase in PPO activation contributes generally to the creation of a fungicide environment often associated with stress (Yedidia et al., 2000). The reduction of those activities in the tissues might lead to a reduction in oxidation of metabolites which are toxic to the pathogen. This agrees to the report of Gorji et al. (2011). Those biochemical changes involve the capacity of the plant to stimulate the pathway of the phenylpropanoids.
CONCLUSION
Heritability was examined in healthy, scarified and inoculated pods. In the (S) fractions, the PPO activity is correlated neither to the sensitivity nor to the productivity of the different genotypes. But in the (I) fractions, the productive and tolerant genotypes (F1307, F1314, F7902 and F7929) and the tolerant and less productive genotypes (F1315, 1315, F7926, F7907) show high PPO proportions after inoculation. Nevertheless, in the same (I) fractions, the less tolerant and productive genotypes (F1324, F1308, F7915, F7919) and the less tolerant and less productive genotypes (F1321, F1326, F7904 and F7911) display low PPO proportions after inoculation. The PPO heritability assessment enables, on one hand, to reveal that the PPO in the (S) fractions are less heritable than those obtained in the (I) fractions and on the other hand, to believe that the PPO in the (I) fractions is a good indicator of resistance. Over all, the present study has enabled to classify the different groups of genotypes as tolerant and less tolerant and so, a large scale distribution of the tolerant and productive genotypes to cocoa farmers will be beneficial.
ACKNOWLEDGMENTS
The authors are gratefully to Cameroon Cocoa Development Corporation (SODECAO) for the field used.
REFERENCES
- Ammar, M.I. and M.A. El-Naggar, 2011. Date palm (Phoenix dactylifera L.) fungal diseases in Najran, Saoudi Arabia. Int. J. Plant Pathol., 2: 126-135.
CrossRefDirect Link - Boudjeko, T., P.F. Djocgoue, J.D. Nankeu, H.D. Mbouobda, N.D. Omokolo and I. El Hadrami, 2007. Luteolin derivatives and heritability of resistance to Phytophthora megakarya in Theobroma cacao. Australas. Plant Pathol., 36: 56-61.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chen, C., R.R. Belanger, N. Benhamou and T.C. Paulitz, 2000. Defense enzymes induced in cucumber roots by treatment with Plant Growth-Promoting Rhizobacteria (PGPR) and Pythium aphanidermatum. Physiol. Mol. Plant Pathol., 56: 13-23.
CrossRefDirect Link - Colak, A., A. Ozen, B. Dincer, S. Guner and F.A. Ayaz, 2005. Diphenolases from two cultivars of cherry laurel (Laurocerasus officinalis Roem.) fruit at an early stage of maturation. Food Chem., 90: 801-807.
CrossRef - Constabel, C.P., L. Yip, J. Patton and M.E. Christopher, 2000. Polyphenol oxidase from hybrid poplar. Cloning and expression in response to wounding and herbivory. Plant. Physiol., 124: 285-296.
Direct Link - Denton, O.A. and C.C. Nwangburuka, 2011. Heritability, genetic advance and character association in six yield related characters of Solanum anguivi. Asian J. Agric. Res., 5: 201-207.
CrossRefDirect Link - Djocgoue, P.F., T. Boudjeko, H.D. Mbouobda, D.J. Nankeu, M.I.B. Efombagn, S. Nyasse and N.D. Omokolo, 2006. Comparative assessment of resistance of cocoa (Theobroma cacao L.) progenies from SNK10xSNK413; ICS84xICS95 to Phytophthora megakarya in Cameroon by measuring size of necrotic lesion along the midrib. J. Plant Pathol., 5: 329-335.
- Djocgoue, P.F., T. Boudjeko, H.D. Mbouobda, D.J. Nankeu, I. El Hadrami and N.D. Omokolo, 2007. Heritability of phenols in the resistance of Theobroma cacao against Phytophthora megakarya, the causal agent of black pod disease. J. Phytopathol., 155: 519-525.
CrossRefDirect Link - Djocgoue, P.F., C. Simo, H.D. Mbouobda, T. Boudjeko, D.J. Nankeu and N.D. Omokolo, 2010. Assessment and heritability of productivity and tolerance level to Phytophthora megakarya in two hybrid populations of Theobroma cacao. J. Plant Pathol., 92: 607-617.
Direct Link - El-Hersh, M.S., K.M. Abd El-Hai and K.M. Ghanem, 2011. Efficiency of molybdenum and cobalt elements on the lentil pathogens and nitrogen fixation. Asian J. Plant Pathol., 5: 102-114.
CrossRefDirect Link - Kasran, R., M. Mahmood, R. Muse and A. Sipat, 2000. Peroxidase, polyphenoloxidase, chitinase and β-1,3 Glucanase in relation to vascular streak dieback disease resistance in Theobroma cacao L. Proceedings of the 13th International Cocoa Research Conference, October 9-14, 2000, Kota Kinabalu, Sabah, Malaysia, pp: 23-30.
- Kim, J.Y., Y.S. Seo, J.E. Kim, S.K. Sung, K.J. Song, G. An and W.T. Kim, 2001. Two polyphenol oxidases are differentially expressed during vegetative and reproductive development in the Fuji apple. Plant Sci., 161: 1145-1152.
CrossRef - Mbouobda, H.D., Fotso, P.F. Djocgoue, N.D. Omokolo, I. El-Hadrami and T. Boudjeko, 2010. Benzo-(1,2,3)-thiadiazole-7-carbothioic S-methyl ester (BTH) stimulates defense reactions in Xanthosoma sagittifolium. Phytoparasitica, 38: 71-79.
CrossRef - Valizadeh, M., J. Valizadeh and M. Jafari, 2011. Identification, distribution and incidence of important tomato and cucurbits viruses in Southeast of Iran. Am. J. Plant Physiol., 6: 242-251.
CrossRefDirect Link - Moradi, Z. and B. Jafarpour, 2011. First report of coat protein sequence of Cucumber green mottle virus in cucumber isolated from Khorasan in Iran. Int. J. Virol., 7: 1-12.
CrossRef - Motamayor, J.C., A.M. Risterucci, P.A. Lopez, C.F. Ortiz, A. Moreno and C. Lanaud, 2002. Cacao domestication I: The origin of the cacao cultivated by the Mayas. Heredity, 89: 380-386.
CrossRefPubMedDirect Link - Ndoumbe-Nkeng, M., C. Cilas, E. Nyemb, S. Nyasse, D. Bieysse, A. Flori and I. Sache, 2004. Impact of removing diseased pods on cocoa black pod caused by Phytophthora megakarya and on cocoa production in Cameroon. Crop Prot., 23: 415-424.
CrossRefDirect Link - Niemenak, N., C. Cilas, C. Rohsius, H. Bleiholder, U. Meier and R. Lieberei, 2010. Phenological growth stages of cacao plants (Theobroma sp.): Codification and description according to the BBCH scale. Ann. Applied Biol., 156: 13-24.
CrossRef - Parthiban, K.T., R. Kirubashankkar, M. Paramathma, V. Subbulakshmi and P. Thiyagarajan et al., 2011. Genetic association studies among growth attributes of jatropha hybrid genetic resources. Int. J. Plant Breed. Genet., 5: 159-167.
CrossRef - Pokou, N.D., J.A.K.N. Goran, I. Kebe, A. Eskes, M. Tahi and A. Sangare, 2008. Levels of resistance to Phytophthora pod rot in cocoa accessions selected on-farm in Cote d'Ivoire. Crop Prot., 27: 302-309.
CrossRef - Schweiggert, U., A. Schieber and R. Carle, 2005. Inactivation of peroxidase, polyphenoloxidase and lipoxygenase in paprika and chilli powder after immediate thermal treatment of the plant material. Innov. Food Sci. Emerg. Technol., 6: 403-411.
CrossRef - Schneider, S. and W.R. Ullrich, 1994. Differential induction of resistance and enhanced enzyme activities in cucumber and tobacco caused by treatment with various abiotic and biotic inducers. Physiol. Mol. Plant Pathol., 45: 291-304.
CrossRef - Aliyu, T.H. and O.S. Balogun, 2011. Effects of variety and planting density on the incidence of common viral diseases of Cowpea (Vigna unguiculata) in a Southern Guinea Savannah Agro-ecology. Asian J. Plant Pathol., 5: 126-133.
CrossRefDirect Link - Vaughn, K.C., A.R. Lax and S.O. Duke, 1988. Polyphenol oxidase: The chloroplast oxidase with no established function. Physiol. Plant, 72: 659-665.
CrossRefDirect Link - Wititsuwannakul, D., N. Chareonthiphakorn, M. Pace and R. Wititsuwannakul, 2002. Polyphenol oxidases from latex of Hevea brasiliensis: Purification and characterization. Phytochemistry, 61: 115-121.
CrossRef - Yedidia, I., N. Benhamou, Y. Kapulnik and I. Chet, 2000. Induction and accumulation of PR proteins activity during early stages of root colonization by the mycoparasite Trichoderma harzianum strain T-203. Plant Physiol. Biochem., 38: 863-873.
CrossRefDirect Link - Moradi, Z., 2011. Diagnosis and molecular variability of watermelon mosaic virus isolates from North, East, North-East and North-West regions of Iran. Asian J. Plant Pathol., 57: 115-125.
CrossRefDirect Link - Ochola, D. and G. Tusiime, 2011. Pathogenicity of rice yellow mottle virus and the potential sources of resistance against the disease in Eastern Uganda. Asian J. Plant Pathol., 5: 1-15.
CrossRef - Gorji, A.H., Z. Zonoori, M. Zolnoori and A. Jamasbi, 2011. Inheritance of antioxidant activity of triticale under drought stress. Asian J. Plant Sci., 10: 220-226.
CrossRefDirect Link