
S. Boukhris-Bouhachem
Laboratoire de Protection des Vegetaux, Institut National de Recherche Agronomique de Tunisie, Rue Hedi Karray, 2049 Ariana, Tunis, Tunisia
J. Rouze-Jouan
Unite Mixte de Recherche, Biologie des Organismes et des Populations appliquee a la Protection des Plantes (BiO3P), INRA/Agrocampus Rennes, Domaine de la Motte B.P. 35327, F-35653 Le Rheu Cedex, France
R. Souissi
Laboratoire de Protection des Vegetaux, Institut National de Recherche Agronomique de Tunisie, Rue Hedi Karray, 2049 Ariana, Tunis, Tunisia
L. Glais
Unite Mixte de Recherche, Biologie des Organismes et des Populations appliquee a la Protection des Plantes (BiO3P), INRA/Agrocampus Rennes, Domaine de la Motte B.P. 35327, F-35653 Le Rheu Cedex, France
M. Hulle
Unite Mixte de Recherche, Biologie des Organismes et des Populations appliquee a la Protection des Plantes (BiO3P), INRA/Agrocampus Rennes, Domaine de la Motte B.P. 35327, F-35653 Le Rheu Cedex, France
Plant Pathology Journal
Year: 2011 | Volume: 10 | Issue: 1 | Page No.: 22-28







ABSTRACT
In the context of Potato virus Y epidemiological study, fourteen aphid species were selected to investigate their relative transmission efficiency in laboratory using tobacco plant tests Nicotiana tabacum var. Xanthi. These aphid species were the ones most often trapped in Yellow Water Traps (YWTs). Transmission efficiency was evaluated in both winged and wingless individuals in cages under controlled conditions. The transmission efficiencies obtained varied from 3 to 95%. Besides Myzus persicae (Sulzer), a highly efficient vector, 13 other aphid species were screened for their capability of transmitting PVYNTN. Three aphid species, Aphis spiraecola Patch, A. gossypii Glover and Brachycaudus helichrysi (Kaltenbach), appeared to propagate PVY greatly, with transmission efficiencies of 73, 71 and 68%, respectively. Even though Aphis fabae Scopoli was less efficient, with only a moderate efficiency of 43%, it is also suspected of being implicated in PVY dissemination. In 60% of the cases, results obtained from wingless and winged forms were very close. Consequently, five aphid species seem to represent a real risk for the spread of viruses given their abundance in traps.
PDF Abstract XML References Citation
Received: November 02, 2010;
Accepted: January 28, 2011;
Published: May 06, 2011
How to cite this article
S. Boukhris-Bouhachem, J. Rouze-Jouan, R. Souissi, L. Glais and M. Hulle, 2011. Transmission Efficiency of the Strain PVYNTN by Commonly Captured Aphids in Tunisian Potato Fields. Plant Pathology Journal, 10: 22-28.
DOI: 10.3923/ppj.2011.22.28
URL: https://scialert.net/abstract/?doi=ppj.2011.22.28
DOI: 10.3923/ppj.2011.22.28
URL: https://scialert.net/abstract/?doi=ppj.2011.22.28
INTRODUCTION
Some of the most damaging diseases in an economically important crop such as potatoes are caused by viruses particularly PVY (Valkonen, 2007) spread by aphid vectors (Buchen-Osmond, 2002). It is more difficult to control non-persistent viruses than persistent ones, since insecticides act too slowly to kill the aphids before they transmit a non-persistent virus because of both the short acquisition access period and inoculation access period (Matthews, 1991). During the aphid’s dispersal phase in search of new hosts, both potato-colonizing and non-potato-colonizing aphid species can transmit Potato virus Y (PVY). The availability of vectors and alternative hosts in potato regions explain the PVY propagation (Bostan and Haliloglu, 2004).
PVY is a non-persistent aphid-transmitted virus and a very harmful pathogen in the potato crop. More than 70 additional aphid species are able to transmit PVY (Varveri, 2000; Halbert et al., 2003; Kerlan, 2006; Ragsdale et al., 2001; Robert et al., 2000). The green peach aphid, Myzus persicae (Sulzer), is by far the most efficient vector of PVY in potato crops worldwide (Ragsdale et al., 2001; Muthomi et al., 2009). Other aphid species are relatively less efficient (Sigvald, 1984; Van Harten, 1983). However, aphid species colonizing potato plants, like Aphis nasturtii Kaltenbach, Macrosiphum euphorbiae (Thomas) and Aulacorthum solani (Kaltenbach), should not be excluded from vector lists, despite their low transmission efficiency, since they too can be responsible for the transmission of PVY within a field. Non-colonizing species such as Brachycaudus helichrysi (Kaltenbach), Phorodon humuli (Schrank) (Rolot, 2005), Rhopalosiphum padi (Linnaeus) (Sigvald, 1989) and Acyrthosiphon pisum (Harris) (Halbert et al., 2003) are also reported to be involved in PVY epidemiology. Given that virus spread into and within a field depends on vector activity and virus sources, knowing aphid efficiency in transmitting PVY is a prerequisite to successfully controlling it.
Tunisian seed potato suffer of PVY contamination and every year we loss 12% of seed potato production (Groupement Inter-professionnel des Légumes (GIL, 2003) which exceed 5% of infected plants with viruses tolerated by the legislation from certified Elite seed >Spunta’ (2% infection) imported from France and Netherland. An epidemiology study was important to be conducted for a better sanitary control. This study is subsequent to the work which previously reported diversity of aphid species in seed potato production in Tunisia using yellow water traps and one suction trap (Boukhris-Bouhachem et al., 2007), 103 aphid species were identified and fourteen of them are numerous and known as PVY vectors. A work was also conducted about PVY strains; the prevalent type was PVYN (Boukhris-Bouhachem et al., 2004). It focuses on the strain PVYNTN, belonging to the PVYN group (Rolland et al., 2008), newly detected in Tunisia (Boukhris-Bouhachem et al., 2008). Aphid transmission of PVYNTN has never been studied before under these conditions and is rarely mentioned by Davis et al. (2005) especially when the changing of their virulence properties is recently reported by Volkov et al. (2009) and Chikh Ali et al. (2010). The objective of this research was to assess PVYNTN transmission efficiency under laboratory conditions of the prevalent aphid species, both colonizing and non-colonizing of potato plants, captured with traps in Tunisian certified seed potatoes in order to know how to control the main vectors and the spread of PVYNTN.
MATERIALS AND METHODS
Aphid species: Those aphid species were selected that are known for their capability to transmit PVY, are abundant in yellow water traps (Boukhris-Bouhachem et al., 2007) and are present on potato foliage (Boukhris-Bouhachem et al., 2004). Four water pan traps were placed in four seed potato production field (Batan, Bousalem, Sidi Mahmoud, Douala) from 2001 to 2006. The traps were weekly collected and aphids identified and counted under stereomicroscope using identification keys. During 2002 and 2003, every two weeks from marsh to may, one hundred potato leaves were collected in each potato field visited to identify colonizing aphids. All potato aphids were from the Cap Bon region (northeastern Tunisia) except Aphis gossypii Glover that was from the northern Manouba region. Non-colonizing aphids were collected from their respective host plants (Table 2) near a potato field in Cap Bon, except for R. padi that was collected from Jendouba in the northwest.
The aphids were reared since 2003, on their known host plants in hermetic closed cages at INRAT, under controlled conditions (temperature 23°C, photoperiod L: D 16: 8 h). All aphids rearing were started from a single parthenogenetic female, except for Aphis spiraecola Patch, Brachycaudus cardui (Linnaeus), B. helichrysi and Hyperomyzus lactucae (Linnaeus), which were reared from mixed populations.
Virus source: PVYNTN is the prevalent strain in Tunisia on potato var. “Spunta” and “Nicola,” characterized on the basis of test plant reactions (N. tabacum var. Xanthi). It is confirmed by RT-PCR protocols with specific primers based on the polymorphism in the P1 genes (Glais et al., 2001) and the three recombinant sites specific to PVYNTN isolates RJ1 and RJ2 (Nie and Singh, 2003) RJ3 (Glais et al., 2005). Among these isolates, only the PVYNTNC1-3 isolated from potato cv. >Spunta’ and cultivated at Cap Bon was used for all of the transmission tests. This virus is maintained in the laboratory, on Nicotiana tabacum var. Xanthi and transmitted mechanically from tobacco plant to tobacco plant. Tobacco seedlings at the 4-leaf stage were inoculated with plant sap. Two weeks after inoculation, the plants developed vein necrosis and leaf distortion, characteristic symptoms of isolates belonging to the PVYN strain group.
Transmission of PVYNTN by aphids: Transmission tests were conducted between 2004 and 2007 both with wingless and winged aphids from tobacco plant to tobacco plant (N. tabacum var. Xanthi) for practical reasons, as was done by many other authors (Basky and Almasi, 2005; Boiteau et al., 1998; Derron and Goy, 1990; Harrington et al., 1986; Kanavaki et al., 2006; Van Hoof, 1980; Volkov et al., 2009). After rearing, 100 wingless individuals were collected in Petri dishes and starved for 2 h. The aphids were then allowed to probe on PVY-infected tobacco leaves for 2 min, called the Acquisition Access Period (AAP). The aphids inoculated twenty tobacco plants. Five aphids were placed on each plant with four true leaves over a 24 h period, called the Inoculation Access Period (IAP). After 24 h, the aphids were killed with an insecticide, Imidacloprid (Confidor, Bayer CropScience), following the manufacturer’s instructions. Three separate repetitions of 20 tobacco plants were carried out for each aphid species.
The protocol for the winged aphids was different. One hundred winged aphids were transferred onto 4 infected tobacco plants, which were put into a cage with 20 healthy tobacco plantlets with 4 true leaves during a 24 h IAP.
Two weeks later, all inoculated plants were tested for the presence of PVY with DAS-ELISA according to Clark and Adams (Clark and Adams, 1977), using a commercial diagnosis kit (PVYN, Bioreba, AG, Reinach, Switzerland) as followed in Boonham et al. (2002). After incubation at room temperature, the absorbance of the samples was read at 405 nm using an automatic microplate reader (Multiscan Ascent Labsystems, Waltham, MA, USA). Tobacco samples with an extinction value exceeding twice the average value of the healthy control were considered PVY positive.
Statistical analyses: All statistical analyses were done using SPSS software. Aphid transmission rates obtained were compared using Duncan’s test. Transmission rates between wingless and winged forms were then analyzed with analyses of variation (ANOVA). The test was significant when p = 0.05.
RESULTS AND DISCUSSION
Five aphid species were identified on potato leaves, Aphis fabae, A. gossypii, Aulacorthum solani, M. persicae and Macrosiphum euphorbiae. The most abundant species was A. gossypii while the least one was A. solani (Table 1).
Major aphid species captured in yellow pan traps were listed in Table 2. Among potato aphids, A. fabae, A. gossypii, M. persicae were the numerous ones compared to M. euphorbiae and A. solani. A. spiraecola and Aphis sp. were the most abundant non colonizing potato aphids especially in Batan and Bousalem. However, high levels of Aphis sp. were observed in Douala. A. pisum was caught with important numbers in three regions: Bousalem, Sidi Mahmoud and Douala. Results indicate that many aphid species were present on potato fields and they may play a role in PVY dissemination.
Except Aphis sp. these aphid species were all used on transmission tests. All of the tested aphid species were vectors of the PVYNTNC1-3 isolate (Table 3). Duncan’s test, applied to transmission efficiency results for wingless aphid species, showed five significantly distinct classes of vectors. As expected, M. persicae was found to be the most efficient vector with a transmission efficiency of 95%; it was therefore grouped into class 1. Following that came A. spiraecola, A. gossypii and B. helichrysi with a transmission efficiency ranging between 68 and 73% which were grouped into class 2. Rhopalosiphum maidis (Fitch), Aphis fabae Scopoli and H. lactucae, with a virus transmission of almost 50%, were grouped into class 3. B. cardui, R. padi and Lipaphis erysimi (Kaltenbach) transmitted PVY at a lower level, ranging between 25 and 34% and were grouped into class 4. Finally, Hyalopterus pruni (Geoffroy), A. pisum, Sitobion avenae (Fabricius) and M. euphorbiae, with a transmission efficiency of less than 10% were the poorest vectors and were grouped into class 5.
The results obtained with winged aphids showed three significant classes using Duncan’s test. No differences were observed between M. persicae, A. spiraecola and A. gossypii (class 1), R. maidis, A. fabae and H. lactucae (class 2) and R. padi, H. pruni, A. pisum and M. euphorbiae (class 3).
The ANOVA showed no significant difference between winged and wingless forms of the same species in six of the ten cases compared. M. persicae was the most efficient PVY vector in both winged and wingless forms with no significant difference.
| Table 1: | Total potato aphids per 100 leaves related to sites and years during March to May |
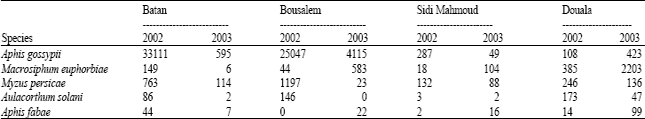 | |
| Table 2: | Mean aphid species per yellow water traps during 2002-2006 |
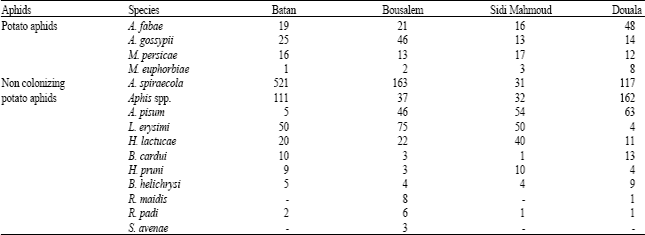 | |
| Table 3: | Transmission efficiency of PVYNTN by different aphid species in percentage |
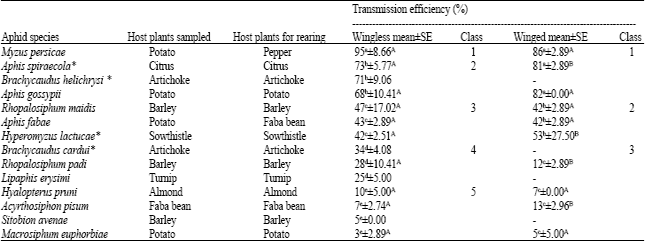 | |
| The aphids were starved for two hours before being subjected to a two-minute AAP and a 24 h IAP; five aphids per plant were used and three replications of 20 tobacco plants (four true leaves) were performed; (*) population origin; (-) not tested; values of aphid transmission efficiency with different lower case letters are significantly different (p = 0.05, Duncan’s test); ANOVA was used t o study significant differences in wingless and winged forms (capital letters) | |
However, the winged forms of A. spiraecola, H. lactucae and A. pisum were significantly more efficient than the wingless forms. In contrast, the winged R. padi was a significantly less efficient vector.
This assay indicates that M. persicae, A. spiraecola, B. helichrysi and A. gossypii were the most important vectors under controlled conditions.
Of the 14 aphid species tested, it was demonstrated that all of them transmitted PVYNTN whether or not they colonized potato plants. PVYNTN is a new prevalent strain in potato fields in Tunisia. Not much data is currently available about the transmission efficiency of PVYNTN by other aphid vectors, as it is for Aphis glycines Matsumura (Davis et al., 2005).
A. gossypii, the cotton aphid with high population levels on potato plants, was an efficient vector under experimental conditions and transmitted PVYNTN with more efficiency than was reported by many authors who obtained a 12 to 31% transmission rate with PVYN (Fereres et al., 1993; Raccah et al., 1985).
The green citrus aphid, A. spireacola, seemed to be a very good vector of PVYNTN under laboratory conditions, in contrast with the transmission rate of 6.2% with PVYN (Raccah et al., 1985).
The leaf-curling plum aphid, B. helichrysi, mentioned by Harrington and Gibson (1989), Piron (1986) and Powell et al. (1992) as a PVY vector with low transmission efficiency (4.8 to 12.5%) was found here to be a good vector (71%). Moreover, it is also considered an important vector in Belgium (Rolot, 2005).
With a moderate efficiency of virus transmission in Tunisia, the black bean aphid, A. fabae (43%) and the currant-sowthistle aphid, H. lactucae (42%), were reported as poor vectors, with a transmission efficiency of 7.6 to 24% and 0.4 to 17.4%, respectively (Harrington and Gibson, 1989; Piron, 1986). The cereal aphid, R. maidis (47%), had not been tested until now (Ragsdale et al., 2001).
The bird-cherry aphid, R. padi, with a transmission rate of 28%, was more efficient in this study than reported before where it varied from 0.5 to 11.5% (Harrington and Gibson, 1989; Harrington et al., 1986; Piron, 1986; Van Hoof, 1980) and less than 40% (Sigvald, 1984). In the same class 4, L. erysimi, the turnip aphid, had a transmission efficiency of 25%, percentage higher than 10% previously mentioned by Ragsdale et al. (2001).
Class 5 aphid species were poor vectors. H. pruni was found to have a transmission efficiency of 8.5%, less efficient than the rate of 13.9% reported by Perez et al. (1995). The transmission efficiency of A. pisum was between 7 and 13%, quite similar to the results (3.8-14%) mentioned by Fereres et al. (1993), Harrington and Gibson (1989), Harrington et al. (1986), Piron (1986) and Raccah et al. (1985). S. avenae also proved to be a poor vector both in this study and in those done by many other authors (Harrington and Gibson, 1989; Perez et al., 1995; Piron, 1986; Sigvald, 1984). S. avenae was therefore not considered a vector. The colonizing potato aphid, M. euphorbiae, transmitted PVYNTN under the conditions here with an efficiency of 4%. This was the same result as that obtained by Kostiw (1980) but different from other authors who found a transmission efficiency of 17 to 29% (De Bokx and Van der Want, 1987).
Potato aphids were present in important numbers on potato fields exceeding the threshold, 3-10 aphids per 100 leaves, recommended for seed potato production (Muthomi et al., 2009; Capinera, 2001). Non-colonizing winged aphids increased opportunity for movement of viruliferous aphids around the crop (Harrington et al., 2003) and are then implicated in PVY epidemiology.
In comparing wingless and winged forms, no significant difference was noted between the two forms of the efficient vectors M. persicae, A. gossypii, A. fabae and R. maidis, except for A. spiraecola. This may be due to similar genetic properties (e.g., virion receptors in the mouthparts) between the two forms of the same species in relation to virus transmission. Nevertheless, transmission efficiency by the winged form was slightly superior in all of the species. This heterogeneity could be attributed to the methodology used, the experimental conditions or the age and activity of winged aphids that increase transmission probability because of flight. In fact, it is unknown if flight behavior is similar for all species or if winged aphids touched on all plants during the IAP. Another factor involved in transmission rates was virus concentration.
PVY transmission variability between the aphid’s species was confirmed by the results obtained here and also reported by Kennedy et al. (1962), Sigvald (1984), Van Harten (1983) and Mirmomeni et al. (2008). Biotype variability of the same species (Verbeek et al., 2010) was also confirmed. Moreover, a difference in transmission efficiency was reported for several other strains of PVY. It has been demonstrated that M. persicae transmit PVYN with more efficiency than PVYO (Basky and Almasi, 2005; Sigvald, 1984; Van Hoof, 1980).
Furthermore, it was clear that M. persicae was the best vector of PVYNTN, with a very high average of transmission efficiency, nearly 100% under laboratory conditions. In general, according to previous studies, PVYNTN is transmitted with more efficiency than PVYN, thus increasing the risk of the spread of PVYNTN in potato crops and making sanitary control more complex.
Four species (Table 1) were very efficient vectors in this experiment (with a transmission rate of >68%). However, a greater number of inefficient vectors may be more important than a fewer number of efficient vectors in the epidemiology of virus diseases (DiFonzo et al., 1997). Their flight behavior and/or high numbers could compensate low transmission efficiencies of some species during the potato-growing season (Sigvald, 1990). Such is the case for the non-colonizing aphids, A. pisum and R. padi, which may be involved in PVY epidemics (Halbert et al., 2003; Sigvald, 1984). These species were trapped in high numbers in the YWTs in Tunisia during the late season.
Colonizing aphid species such as M. persicae and A. gossypii probably play a role in the epidemiology of PVY since they transmit PVY rather efficiently. A. fabae, a moderately efficient vector, is frequently captured in YWTs (Boukhris-Bouhachem et al., 2007) and is likely to contribute to the dissemination of PVYNTN as well.
Non-colonizing winged aphid species, such as A. pisum, A. spiraecola, B. helichrysi, B. cardui, Dysaphis sp., H. lactucae, H. pruni, L. erysimi, R. padi and S. avenae, were also found on potato plants during the growing season. Among these, A. pisum, A. spiraecola, B. cardui and H. lactucae were commonly captured in YWTs (Boukhris-Bouhachem et al., 2007). All of these species have been found to transmit PVY and seem to contribute greatly to the PVY epidemic during the season compared to their number. In fact, A. pisum, A. spiraecola, B. helichrysi, H. lactucae, L. erysimi and R. padi are often caught in large numbers in YWTs, meaning that these non-colonizing aphid species can be held responsible for the epidemic spread of PVYNTN (DiFonzo et al., 1997; Halbert et al., 1981).
Simulating the situation occurring in the field with winged aphids in cages gave better practical results than the laboratory experiments with wingless individuals obligated to feed on tobacco which gave transmission efficiencies that were too high for those aphid species that do not colonize potato plants. By taking the vector efficiency of a particular aphid species into account together with its abundance in the traps, the potential risk of infection can be evaluated for potato crops (Sigvald, 1986). The use of tobacco in laboratory transmission tests gives an indication of the vector’s behavior and its capacity to transmit PVYNTN under controlled conditions. Based on these results (Table 3) and aphid abundance (Boukhris-Bouhachem et al., 2007), it can be assumed that M. persicae, A. spiraecola, A. gossypii, A. fabae and A. pisum are the key vectors of PVYNTN in potato crops in Tunisia.
Transmission efficiencies for PVYNTN strains seem to be higher within the others PVY strain and seem determined by aphid species, by clone used and virus concentration in the source leaf. These results will lead to a better understanding of PVYNTN epidemiology and to a better control of aphid vectors in order to improve seed potato sanitary conditions. However, the transmission method must be improved by using potato plants and it must be standardized in order to use it as a routine technique to evaluate the vector capacity of aphids in transmitting PVYNTN in the field.
ACKNOWLEDGMENTS
A sincere thank you to Dr. I. Chaib from the National Institute of Agronomic Research of Tunisia (Institut National de la Recherche Agronomique de Tunisie) for his help with the statistical analyses. Thanks must also be given to P. Piron, from Plant Research International B.V. Droevendaalsesteeg 1, The Netherlands, for his interesting remarks. Thanks C. Chaar for careful revision of the English.
REFERENCES
- Basky, Z. and A. Almasi, 2005. Differences in aphid transmissibility and translocation between PVYN and PVYO isolates. J. Pest. Sci., 78: 67-75.
Direct Link - Boiteau, G., M. Singh, R.P. Singh, G.C.C., Tai and T. R. Turner, 1998. Rate of spread of PVYN by alate Myzus persicae (Sulzer) from infected to healthy plants under laboratory conditions. Potato Res., 41: 335-344.
CrossRefDirect Link - Boukhris-Bouhachem, S., N. Khamassy, L. Glais and C. Kerlan, 2008. Occurrence in Tunisia of potato tuber necrotic ringspot disease (PTNRD) caused by variant PVYNTN of Potato virus Y. Plant Pathol., 57: 388-388.
CrossRef - Boukhris-Bouhachem, S., R. Souissi, E. Turpeau, J. Rouze-Jouan, M. Fahem, N.B. Brahim and M. Hulle, 2007. Aphid (Hemiptera: Aphidoidea) diversity in Tunisia in relation to seed potato production. Annales Societe Entomologique France, 43: 311-318.
CrossRefDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Davis, J.A., E.B. Radcliffe and D.W. Ragsdale, 2005. Soybean aphid, Aphis glycines Matsumura, a new vector of potato virus Y in potato. Am. J. Potato Res., 82: 197-201.
CrossRef - De Bokx, J.A. and J.P.H. Van der Want, 1987. Viruses of Potatoes and Seed-Potato Production. 2nd Edn., Pudoc, Wageningen, ISBN: 90-220-0859-2 pp: 259.
Direct Link - DiFonzo, C.D., D.W. Ragsdale, E.B. Radcliffe, N.C. Gudmestad and G.A. Secor, 1997. Seasonal abundance of aphid vectors of potato virus Y in the Red River Valley of Minnesota and North Dakota. J. Econ. Entomol., 90: 824-831.
Direct Link - Fereres, A., P. Perez, C. Gemeno and F. Ponz, 1993. Transmission of Spanish pepper and potato PVY isolates by aphid (Homoptera: Aphididae) vectors: Epidemiological implications. Environ. Entomol., 22: 1260-1265.
Direct Link - Glais, L., M. Tribodet and C. Kerlan, 2001. Molecular detection of particular PVY isolates: PVYNTN and PVYNW. Proceedings of the 11th EAPR Virology Section Meeting, Oct. 7-13, Halickuv Brod-Trest, Czech, pp: 70-71.
Direct Link - Glais, L., M. Tribodet and C. Kerlan, 2005. Specific detection of the PVYN-W variant of Potato virus Y. J. Virol. Meth., 125: 131-136.
CrossRef - Halbert, S.E., D.L. Corsini and M.A. Wiebe, 2003. Potato virus Y transmission efficiency for some common aphids in Idaho. Am. J. Potato Res., 80: 87-91.
Direct Link - Halbert, S.E., M.E. Irwin and R.M. Goodman, 1981. Alate aphid (Homoptera: Aphididae) species and their relative importance as field vectors of soybean mosaic virus. Ann. Applied Biol., 97: 1-9.
CrossRef - Harrington, R. and R.W. Gibson, 1989. Transmission of potato virus Y by aphids trapped in potato crops in southern England. Potato Res., 32: 167-174.
CrossRefDirect Link - Harrington, R., N. Katis and R.W. Gibson, 1986. Field assessment of the relative importance of different aphids species in the transmission of potato virus Y. Potato Res., 29: 67-76.
CrossRefDirect Link - Kanavaki, O.M., J.T. Margaritopoulos, N.I. Katis, P. Skouras and J.A. Tsitsipis, 2006. Transmission of potato virus Y in tobacco plants by Myzus persicae nicotianae and M. persicae s.str. Plant Dis., 90: 777-782.
CrossRefDirect Link - Muthomi, J.W., J.N. Nyaga, F.M. Olubayo, J.H. Nderitu and J.N. Kabira et al., 2009. Incidence of aphid-transmitted viruses in farmer-based seed potato production in Kenya. Asian J. Plant Sci., 8: 166-171.
CrossRefDirect Link - Nie, X. and R.P. Singh, 2003. Specific differentiation of recombinant PVYN:O and PVYNTN isolates by multiplex RT-PCR. J. Virol. Methods, 113: 69-77.
CrossRefDirect Link - Perez, P., J.L. Collar, C. Avillza, M. Duque and A. Fereres, 1995. Estimation of vector propensity of potato virus Y in open-field pepper crops of central Spain. J. Econ. Entomol., 88: 986-991.
Direct Link - Piron, P.G.M., 1986. New aphid vectors of potato virus YN. Eur. J. Plant Pathol., 92: 223-229.
CrossRefDirect Link - Raccah, B., A. Gal-on and V.F. Eastop, 1985. The role of flying aphid vectors in the transmission of cucumber mosaic virus and potato virus Y to peppers in Israel. Ann. Appl. Biol., 106: 451-460.
CrossRef - Robert, Y., J.A.T. Woodford and D.G. Ducray-Bourdin, 2000. Some epidemiological approaches to the control of aphid-borne virus diseases in seed potato crops in Northern Europe. J. Virus Res., 71: 33-47.
CrossRef - Sigvald, R., 1984. The relative efficiency of some aphid species as vectors of potato virus Yo (PVYo). Potato Res., 27: 285-290.
CrossRef - Sigvald, R., 1989. Relationship between aphid occurrence and spread of potato virus Y° (PVY°) in field experiments in Southern Sweden. J. Applied Entomol., 108: 35-43.
CrossRefDirect Link - Sigvald, R., 1990. Aphids on potato foliage in Sweden and their importance as vectors of potato virus YO. Acta Agric. Scand., 40: 53-58.
Direct Link - Van Harten, A., 1983. The relation between aphid flights and the spread of Potato virus YN in the Netherlands. Potato Res., 26: 1-15.
CrossRef - Varveri, C., 2000. Potato Y potyvirus detection by immunological and molecular techniques in plants and aphids. Phytoparasitica, 28: 141-148.
CrossRef - Verbeek, M., P.G.M. Piron, A.M. Dullemans, C. Cuperus and R.A.A. van der Vlugt, 2010. Determination of aphid transmission efficiencies for N, NTN and Wilga strains of Potato virus Y. Ann. Applied Biol., 156: 39-49.
CrossRefDirect Link - Volkov, Y.G., N.N. Kakareka, L.A. Balabanova, Z.N. Kozlovskaya and M.V. Sapotsky, 2009. Characterization of a novel far-eastern potato virus y isolates. Plant Pathol. J., 8: 62-67.
CrossRef - Mirmomeni, M.H., A. Sharifi and S. Sisakhtnezhad, 2008. Rapid detection of potato Y potyvirus in potato farms of kermanshah using RT-PCR amplification of the P1-protease gene and its cloning. Pak. J. Biol. Sci., 11: 1482-1486.
CrossRefPubMedDirect Link - Bostan, H. and K. Haliloglu, 2004. Distribution of PLRV, PVS, PVX and PVY (PVYN, PVYo and PVYc) in the seed potato tubers in Turkey. Pak. J. Biol. Sci., 7: 1140-1143.
CrossRefDirect Link - Boonham, N., K. Walsh, M. Hims, S. Preston, J. North and I. Barker, 2002. Biological and sequence comparisons of Potato virus Y isolates associated with potato tuber necrotic ring spot disease. Plant Pathol., 51: 117-126.
Direct Link - Chikh Ali, M., T. Maoka, T. Natsuaki and K.T. Natsuaki, 2010. PVYNTN-NW, a novel recombinant strain of Potato virus Y predominating in potato fields in Syria. Plant Pathol., 59: 31-41.
CrossRef