
Mujeebur Rahman Khan
Department of Plant Protection, Faculty of Agricultural Sciences, Aligarh Muslim University, Aligarh 202 002, India
Mohd Mahmud Khan
Department of Plant Protection, Faculty of Agricultural Sciences, Aligarh Muslim University, Aligarh 202 002, India
Plant Pathology Journal
Year: 2011 | Volume: 10 | Issue: 1 | Page No.: 1-12







ABSTRACT
Development of plant diseases depends largely on environmental factors besides the host susceptibility. Contamination of air due to release of phytotoxic pollutants such as SO2 may influence the composition of environment and consequently the host parasite relationship. SO2 enters in plants through open stomata, the gas reacts with moisture and is converted into acid. The acidic ions cause toxicity to the plant tissue as well as to pathogenic spores especially those present on the foliage. There are considerable reports, which indicate that plants show varied response to foliar pathogens under SO2 stressed air. In addition, root-attacking pathogens, such as plant nematodes may also be influenced due to host mediated effects of SO2. Generally, suppressive effects of the gas on pathogens have been reported at the concentrations above 200-300 ppb SO2. However, some studies have also revealed greater severity of diseases on plants exposed to lower concentration of the gas in the range of 50-100 ppb. The present paper has critically analyzed the information available on the topic and has attempted to provide explanation to the varied response of plants to pathogens under SO2 polluted environments and the mechanism involved in the interaction.
PDF Abstract XML References Citation
Received: October 29, 2010;
Accepted: November 05, 2010;
Published: May 06, 2011
How to cite this article
Mujeebur Rahman Khan and Mohd Mahmud Khan, 2011. Plants Response to Diseases in Sulphur Dioxide Stressed Environment. Plant Pathology Journal, 10: 1-12.
DOI: 10.3923/ppj.2011.1.12
URL: https://scialert.net/abstract/?doi=ppj.2011.1.12
DOI: 10.3923/ppj.2011.1.12
URL: https://scialert.net/abstract/?doi=ppj.2011.1.12
INTRODUCTION
Air is an important vital resource for the sustenance as well as development of living organism. The composition of its minor constituents often varies as a result of the emission or contaminants emerging from various activities. Environmental pollution is a direct consequence of rapid industrialization and urbanization which is associated with increased demand for energy as well as technological limitation on the emission control (Khan and Khan, 2000; Emberson et al., 2001; Yang et al., 2002). A huge amount of toxic materials originating from different kinds of industries and other human activities are released into air, which ultimately impure the atmosphere. In the developing countries, Sulphur Dioxide (SO2) is one of the most prevalent phytotoxic gaseous pollutants released due to combustion of fossil fuels (Khan and Khan, 1993a) and causes disorders in plants with specific symptoms (Barret and Benedict, 1970; Varshney and Garg, 1979; Khan and Khan, 1993b).
Sulphur dioxide at low concentration can stimulate physiology and growth of plants, especially in plants growing in sulphur deficient soil (Darrall, 1989) where the sulphate might be metabolized to fulfill the demand for sulphur as a nutrient (De Kok, 1990). Increased uptake of SO2, however, can cause toxicity and reduce growth and productivity of plants due to accumulation of sulphite and sulphate ions in excess (Darrall, 1989; Agrawal and Verma, 1997). Sulphur dioxide has been recognized as inanimate or abiotic pathogen as the gas harms plants by suppressing the plant growth and dry matter production and causing typical symptoms. These adverse effects occur through inhibition in the photosynthesis, impairment of vital physiological processes, reduction in chlorophyll pigments etc (Iqbal et al., 2000).
Contamination of air by SO2 and effects on plants and pathogens: A number of natural and anthropogenic sources release SO2 into the atmosphere. The primary source of SO2 are volcanoes and biomass burning, while the main anthropogenic sources of SO2 are burning of fossil fuels, metal smelting and processing of oil and natural gas (Cullis and Hirschler, 1980; Brimblecombe et al., 1989). Together natural and anthropogenic sources emit an estimated 194 million tonnes SO2 per annum, of which 83% is due to fossil fuel combustion (Watsan et al., 1990). Although considerable progress has been made in the development and implementation of SO2 control technologies in North America, Europe and some Asian countries such as Japan, China and Korea, ambient SO2 concentrations are still a significant problem in different parts of the world especially in Assia (Yunus et al., 1996; Innes and Haron, 2000).
| Table 1: | Emissions due to coal burning in power plants |
 | |
In many developing countries in Asia and Africa, of the mined coal is burnt in the power generating units (Haggestad et al., 1986). The burning of coal liberates high concentrations of oxides of sulphur, nitrogen carbon, particulate matter etc. into the environment (Table 1). The concentration of SO2 varies with the distance and direction of wind. Stacks of the power plants are generally equipped with pollution control devices, which are more efficient against particulate matter than gases, hence the surrounding environment of approximately 10-20 km diameter or much more may experience higher concentrations of gaseous pollutants, SO2 in particular (Khan et al., 1998). Ambient concentration of SO2 may range 59-356 μg m-3 around power plants in windward direction depending on the quality, amount and burning temperature of the coal and height of the stack, in addition to efficiency of air pollution devices fitted to the stack (Haggestad et al., 1986; Khan and Khan, 1994a).
Typical peak ambient SO2 concentrations vary from <1 ppb in remote areas to 1-30 ppb in rural areas and 30-200 ppb in moderately polluted areas and to 200-2000 ppb in heavily polluted areas (Finlayson-Pitts and Pitts, 1986). Once emitted, SO2 is transferred from the atmosphere onto surfaces by diffusion (dry as well as wet deposition) at variable rates which are strongly influenced by meteorological conditions. It is also important to note that SO2 in the atmosphere is also transformed to SO42¯ at variable rates and these SO42¯ particles are deposited onto surfaces by Brownian motion (dry deposition) and by precipitation (wet deposition). Any observed foliar injury or changes in plant growth and productivity due to SO2 exposures are the result of dry/wet deposition and subsequent uptake of sulphate and sulphite ions by the plants.
Exposure of plants to SO2 leads to development of visible foliar injury and symptoms. Sulphur dioxide has been recognized as inanimate or abiotic pathogen as the gas harms plants by suppressing their growth and dry matter production and causing specific symptoms.
Plants in nature may be attacked simultaneously by abiotic and biotic pathogens. When these two kinds of pathogens co-exist in a common environment may interact and develop some relationship, which may be as follows:
| • | If air pollutants, directly or indirectly through the host plant are toxic to biotic pathogens, they may suppress the disease and subsequently an antagonistic interaction would occur |
| • | The pollutants may predispose the host plant for greater invasion and damage by the pathogen(s) by modifying host physiology and/or injuring the host tissue so as to make the plant much susceptible or easily invadable by the pathogen. This may lead to a synergistic relationship between pollutant and pathogen or vice versa, thus causing greater damage to the host plant |
| • | Both pollutant and pathogen may act independently without influencing the activity of each others |
Exposures to SO2 may eventually leads to the accumulation of sulphur, increased acidity and other changes in plant physiology and biochemistry that may affect pathogenesis of microbes and subsequently the disease development. The environmental pollution may have a direct effect on plants and pathogens and it may alter the host parasite relationship influencing virulence of the pathogen and/or susceptibility of the host. Direct effect of SO2 on fungal spores has been investigated in ambient and simulated conditions as well as in vitro. Since long germicidal nature of SO2 has been known to man but probably such observation in ambient condition was first made by Kock (1935) when he noticed absence of powdery mildew fungus, Microspheaera alni on oak trees in the vicinity of a paper mill in Australia.
To understand development of plant diseases under SO2 stress environment and the mechanisms of interaction between the gas and pathogen it is essential to first know the effect of SO2 on the growth and development of plants which provide substrate to pathogens for infection and multiplication.
Effects on plants
SO2 uptake by plants: Under normal condition sulphur is prominently taken up by the roots in the form of sulphate ions (De Kok et al., 2002, 2004) assimilated into organic sulphur compounds used in various biochemical process (De Kok et al., 2002, 2004).
Sulphur dioxide after diffusion into mesophyll cells is converted into sulphite and sulphate ions. The absorbed SO2 in the mesophyll cells of the shoot may enter the sulphur reduction pathway either as sulphite or sulphate. Excess SO2 is transferred into the vacuole as sulphate, where it is slowly metabolized (Cram, 1990; Clarkson et al., 1993). Even at relatively low atmospheric concentration of SO2 results in an enhancement in the sulphur content of the foliage because of accumulation of sulphate in the vacuole (De Kok, 1990; De Kok and Tausz, 2001). Chinese cabbage is highly susceptible to sulphur dioxide as a strong linear relation between the rate of uptake of SO2 and the atmospheric concentrations (0.03-1.4 μL L-1) has been recorded (Yang et al., 2006).
Biomass of cabbage was reduced upon prolonged exposure to ≥0.1 μl L-1 SO2. The gas exposure resulted in an increase in the SO42¯, water-soluble non-protein thiols and total S content of the shoot at concentrations ≥0.1 μl L-1; however, the ratio of organic S to total S remained unaffected. The organic N content was not affected by SO2 exposure.
Sulphur dioxide injury in plants: The sulphite ions are about 30 times more toxic than sulphate ions (Thomas et al., 1943). Two general types of markings or symptoms designated as chronic and acute are produced by the plants due to the accumulation of sulphate and sulphite ions in the leaf tissue. Chronic injury occurs on exposure of plants to low concentration of SO2 (less than 100 ppb) at which the rate of accumulation of the ions is slow the cells oxidize the sulphite ions and injury occurs until sufficient sulphate ions accumulate. This type of chronic injury is characterized by a general chlorotic appearance of the leaves. Cells are not killed but the chlorophyll is bleached which appears as a mild chlorosis or yellowing of the leaf or a silvering or bronzing of the lower leaf surface without necrosis (Darley and Middleton, 1966). In an experiment, mustard cultivars were exposed intermittently to 2 (ambient), 25, 50 and 75 ppb SO2 for 5 h on alternate day for three months in open top exposure chambers. More than 90% of the germplasm sustained acute injury in the form of yellowing and mild chlorosis of leaves (Khan et al., 2007). Acute injury results from the absorption of lethal quantities of SO2. It appears as marginal or intercostal areas of dead tissues, which are at full grayish green water soaked in appearance. In most plant species, these areas become bleached in original colour, upon drying and dead or necrotic areas may fall out leaving a ragged appearance to the leaf. In case of severe injury abscission layer develops at the base of petiole and the leaves fall down (Jacobson and Hill, 1970).
Shaw et al. (1993) reported the effects of SO2 exposure at 34 and 58 μg m-3 on needle necrosis in Scots pine (Pinus sylvestris L.). Regression analysis indicated that the appearance of foliar injury was related to the mean SO2 concentration during a critical growth period, although injury did not become visible until 5 weeks later. SO2 at 58 μg m-3 caused foliar injury to a greater number of trees in 2 of the 3 survey years and foliar injury appeared on the same trees in consecutive years suggesting that the sensitivity was genetic. A subsidiary fumigation chamber experiment was performed to see if the injury symptoms observed in the field could be duplicated. Exposure to 655, 1,310 and 2,619 μg SO2 m-3 for 4 h on Scots pine seedlings produced no effects in any treatment. It was suggested that this may have been due to a low replicate number resulting in a few plants at the most sensitive stage of growth, and/ or low humidity during fumigation. Intermittent exposure of tomato plants (cv. Pusa Ruby) to SO2 at 286 μg m-3 (3 h every third day for 75 days) induced slight chlorosis of leaves, however, at 571 μg SO2 m-3 considerable chlorosis with browning developed on the foliage (Khan and Khan, 1993a).
Effect on physiology and plant growth: Seasonal variation in the sensitivity of plants to SO2 also occurs (Cormis, 1973). The physiological status and age of the plants are other important factors in this respect are important (Tamm and Cowling, 1976). Low concentration of SO2 reduces the net photosynthesis in plants. However, short and long exposure duration increase the rate of transpiration and dark respiration (Takemoto and Noble, 1982; Saxe, 1983a, b). Clapperton and Reid (1994) screened genotypes of timothy (Phleum pratense) for SO2 sensitivity in experiments conducted in closed fumigation chambers. In the first experiment, plants were exposed to 393 to 524 μg SO2 m-3 or 3 weeks, the plants developed chlorotic areas, browning and necrosis of the leaves. In a second experiment, plants exposed to 170 μg SO2 m-3 and the experiment was terminated when plants showed the first sign of damage in two weeks. Plants were considered tolerant when they exhibited no sign of visible injury and no significant decrease in shoot or root dry weights compared to unexposed plants. Foliar and flower injury occurred in Calendula officianalis (Padhi et al., 1995) and Zinnia (Rath et al., 1995) and the intensity of symptoms increased with SO2 concentration and duration of exposure. Rakwal et al. (2003) observed distinctive reddish brown necrotic spots and interveinal browning appeared on the leaf surface of rice seedling cv. Nipponbare after exposure to SO2 over control, partly reminiscent of the hypersensitive reaction lesions. Intermittent exposure of SO2 at 200 and 300 μg m-3 caused chlorosis of the leaves of pumpkin with or without inoculation of M. javanica. A mild chlorosis appeared only in the infected plants at 100 μg SO2 m-3 (Khan et al., 1995). Sulphur dioxide (0.1 ppm) induced foliar chlorosis on two cultivars of cowpea viz., V-38-1 and V-218 which appeared earlier in the presence of root-knot nematode (M. incognita) (Khan and Khan, 1996a).
Sulphur dioxide has been reported to cause significant suppression in the plant growth and/or yield of a number of crop plants viz., tomato (Khan and Khan, 1991; Khan and Khan, 1994b), egg plant (Khan and Khan, 1997), cowpea (Khan and Khan, 1996a), okra (Khan and Khan, 1994a), soybean (Singh et al., 1996, 1997), bottle gourd (Khan and Khan, 1991; Khan et al., 1998), pea (Singh et al., 1995; Prakash et al., 1997) and mustard (Khan et al., 2007).
Effects of Sulphur dioxide on plant diseases: When SO2 and plant pathogens co-occur in a given ecosystem, the gas may prove toxic to pathogen and suppress its pathogenicity. Exposures to SO2 may eventually leads to the accumulation of sulphur, increased acidity and other changes in plant physiology and biochemistry. Any alterations in plant physiology and biochemistry may affect pathogenesis of microbes and subsequently the disease development. The review of researches carried out on pathogen pollutant (SO2) interaction has shown considerable information on the effect of plant diseases caused by fungi and nematodes, whereas plant pathogenic viruses and bacteria have almost been neglected in this regard.
Effect of SO2 on plant diseases caused by fungi: Considerable information is available on effect of SO2 on plant pathogenic fungi and the diseases they cause. However, the effects are conflicting and far from generalization. These informations have emerged from the experiments carried under in vitro and in vivo conditions. Most of these studies show inhibitory or stimulatory effects of SO2 on the pathogens (Table 2) and are summarized as under.
| Table 2: | Effect of sulphur dioxide on plant pathogenic fungi |
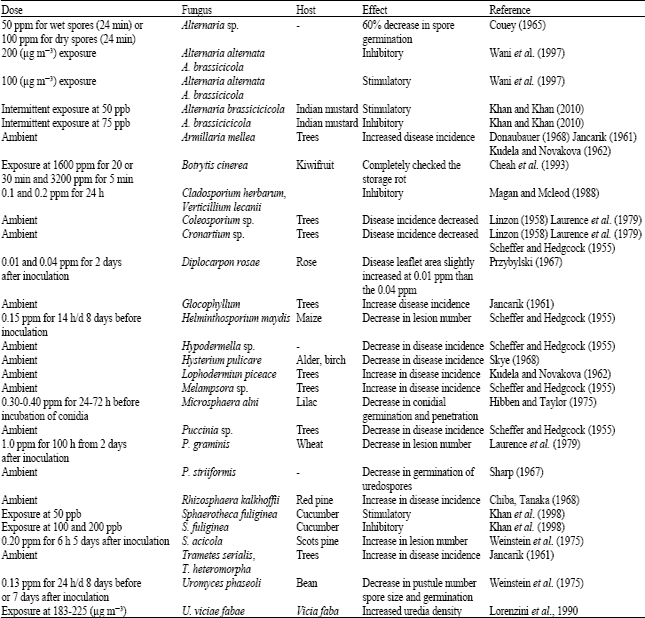 | |
In vitro condition
Inhibition of pathogens: In vitro studies have revealed that SO2 inhibits fungal parasitism but the mechanism involved is not known (Heck et al., 1986). Spores as well as mycelium of most of the fungi appear to be resistant to direct exposure to SO2 and even massive doses have little effect on spore germination. The germination of uredospores of Puccinia striiformis was less in ambient air with a higher concentration of SO2 (Sharp, 1967). Magan and McLeod (1988) reported significant inhibitory effect of 0.1 and 0.2 ppm SO2 for 24 h on spore germination and germ tube extension of Verticillium lecanii and Cladosporium herbarum. Conidial germination of powdery mildew viz., Erysiphe trifolii, E. pisi, E. polygoni, Microsphaera alphitoides, Phyllactinia dalbergiae, Sphaerotheca fuliginea and S. cassia were suppressed on exposure to 0.1 and 0.2 ppm SO2 (Khan and Kulshreshtha, 1991). Exposures at 200 ppb SO2 suppressed the conidial germination of S. fuliginea.
By and large, fungal spores are quite resistant to SO2 but they may become sensitive under moist and humid condition. Germination of wet conidia of Alternaria sp. was decreased by 60% from exposure to 50 ppm SO2 for 24 min, however, 110 ppm produced similar effects on dry spores (Couey, 1965). Wani et al. (1997) have recorded inhibitory effect of 100 and 200 μg SO2 m-3 on the colonization of Alternaria alternata and A. brassicicola. Length and number of cells of a spore increased due to exposure to 100 μg SO2 m-3 and remained uninfluenced at 200 μg concentration.
Stimulation of pathogens: Sporulation and spore germination of Alternaria brassicae and A. brassicicola were stimulated on exposure to 100 μg SO2 m-3 for 3-9 h (Wani et al., 1997). Khan et al. (1998) have reported that intermittent exposures of SO2 at 50 ppb increased the germination of conidia of Sphaerotheca fuliginea.
Simulated condition: Considerable researches have been conducted under artificial treatment conditions in closed-top or open-top exposure chambers to evaluate the effect of SO2 on plant diseases caused by fungi. These researches, however, have shown varied response of the plant pathogenic fungi (Table 2).
Inhibition of pathogens: An open-air fumigation system was utilized to assess the development of mycoflora (yeast and filamentous fungi) on winter barley (Hordeum vulgare L.) cv. Igri in the presence of SO2 or two fungicides (Magan and McLeod, 1991). Exposures to 37, 76 or 123 μg SO2 m-3 in the first year or 37, 73 or 100 μg SO2 m-3 throughout the growing season in the second year experiment resulted in consistently lower number of colony forming units on flag leafs while the fungicides had no consistent effect. Significant decrease was observed in populations of pink yeast (Sporobolomyces roseus) under high SO2 concentration plots in both years. White yeast (Cryptococcus spp.) populations also decreased with SO2 exposure while populations of Cladosporium spp. remained unaffected. Both fungicide treatments significantly reduced Cladosporium spp. populations in the first year. A significantly greater percentage of green flag leaf area was found in the SO2 treated plots compared to the fungicide treated plots. It was concluded that yeast and Cladosporium species were important component of cereal mycoflora and act as antagonists to foliar pathogens. Thus, a reduction in their populations by SO2 could reduce plant defense to pathogen attack.
Cheah et al. (1993) studied the effect of sulphur dioxide fumigation on storage-rot caused by Botrytis cinerea and residues in kiwifruit (Actinidia deliciosa). Fumigation with 1600 ppm for 20 or 30 min and 3200 ppm for 5 min almost completely checked the storage-rot. Absorption of SO2 by fruit exposed to 1600 ppm gradually increased with time from 5 to 30 min of fumigation. The rate of absorption was greater at 3200 ppm than at 1600 ppm. Total SO2 residues were proportional to exposure time and treatment rate. The residues declined sharply from a maximum of 4.0 to 1.6 ppm after 24 h and to 1.3 ppm 5 days after fumigation. No SO2 injury was observed on fruit at any concentration-time combination tested and fruit firmness was not affected by the treatments.
Khan et al. (1998) reported that air polluted with higher levels of SO2 may directly alter the environment around the plant and pathogen. To test the hypothesis that plants may respond differently to foliar pathogens in polluted environments, effects of intermittent exposures of SO2 at 286 and 571 μg m-3 were investigated on the development of powdery mildew of cucumber (Cucumis sativa L.) caused by Sphaerotheca fuliginea, using pre, post and concomitant inoculation exposures in closed-top chambers. Sulphur dioxide and the fungus alone caused chlorosis and/or necrosis and mildew colonization on leaves and subsequently reduced the plant growth and yield of cucumber. Fungus colonization was greatly suppressed at higher concentrations. Gas injury on fungus-infected plants was also less in the other treatments. Conidia of S. fuliginea collected from exposed plants varied in size. Higher concentrations, 286 and 571 μg SO2 m-3, however, suppressed the germination of conidia collected from the exposed plants or exposed on glass slides. The number of fibrosin bodies declined at all the concentrations. Sulphur dioxide at 571 μg m-3 and powdery mildew infection acted antagonistically and their combined effects on plant growth were less than the additive effects.
Khan and Khan (2010) investigated the effect of intermittent exposure of SO2 on the leaf blight caused by Alternaria brassicicola on Indian mustard. The gas at 75 ppb inhibited the pathogenesis of A. brassicicola. Interaction of 75 ppb SO2 concentration and A. brassicicola was found to be antagonistic. At 75 ppb SO2, the sporulation was suppressed. The study has shown that the effects of SO2 on Alternaria blight were suppressive at 75 ppb SO2.
Stimulation of pathogens: Vicia faba cv. Aguadulce was grown in fumigation chambers ventilated with filtered air or air containing SO2 at controlled concentrations and inoculated with the rust fungus Uromyces viciae-fabae to investigate the effect of SO2 on the parasitism of rust fungus on Vicia faba (Lorenzini et al., 1990). Exposures to SO2 levels around 70-90 η1 L-1 enhanced uredia density. Pre-inoculation fumigations at 70-86 η1 L-1 increased uredia density, whilst post-inoculation fumigations stimulated urediospore production. Lower concentrations increased diffusion resistance of the host plant and slightly increased uredia density. Concentrations between 100 and 130 η1 L-1 were somewhat inhibitory for the parasite rust fungi. Total sulphur accumulation in leaf tissue was reduced in presence of the fungal infection. SO2 pollution neither decreased germination ability nor modified morphological parameters of the urediospores. However, elongation of the germination tube of spores coming from filtered air was adversely affected by the pollutant.
Over a three-year period, Mansfield et al. (1991) examined the effects of SO2 exposure in an open-air fumigation system on the development of fungal diseases of winter barley (Hordeum vulgare L.) cv. Igri. Exposures at 24 (ambient), 55, 84 and 113 μg SO2 m-3 were applied in 1984-85, 18 (ambient), 37, 76 and 126 μg SO2 m-3 in 1985-86 and 13 (ambient), 64, 73 and 100 μg SO2 m-3 in 1986-87, throughout the growing season. A general increase in powdery mildew infection in plots exposed to SO2 and decreased leaf blotch infection was recorded at the time of sampling. However, there was no effect of SO2 treatment in any year on the incidence of sharp eyespot, Fusarium foot rot, brown rust, glume blotch and net blotch on the flag leaves but variable effects were observed for eyespot and black ear moulds. Khan et al. (1998) investigated the effects of intermittent exposures of SO2 at 143 μg m-3 on the development of powdery mildew of cucumber (Cucumis sativa L.) caused by Sphaerotheca fuliginea, using pre, post and concomitant inoculation exposures in closed-top chambers. Fungus colonization was relatively greater on the plants exposed to 143 μg SO2 m-3. Gas injury on fungus-infected plants was also less in the other treatments. Conidia of S. fuliginea collected from exposed plants varied in size. Conidial germination was considerably greater at 143 μg SO2 m-3. The number of fibrosin bodies declined at all the concentrations. Synergistic relationship between 143 μg SO2 m-3 and S. fuliginea was recorded on plant growth and yield of cucumber. Intermittent exposures of mustard plants to SO2 at 25 and 50 ppb promoted pathogenesis of A. brassicicola resulting to greater blight and plant growth reductions (Khan and Khan, 2010).
Ambient condition
Inhibition of pathogens: Since long germicidal nature of SO2 has been known to man but probably such observation in ambient condition was first made by Kock (1935) when he noticed absence of powdery mildew fungus, Microspheaera alni on oak trees in the vicinity of a paper mill in Australia. The disease was quite prevalent on oaks in an area which was agroclimatically identical but away from the smokes (SO2) of the mill. Industrial emissions containing SO2 may decrease incidence of the disease caused by various fungi viz. Alternaria solani, Hypodermella juniper, Hysterium pulicara, Rhystisma acerinum, Sphaerotheca fulginea, Venturia inequalis etc. (Przybylski, 1967). Scheffer and Hedgcock (1955) observed low intensity and incidence of the diseases caused by species of Coleosporium, Cronartium, Melampsora, Peridermium or Puccinia on trees grown in SO2 polluted area (near an ore smelter) and sustained pollution injury. With a gradual increase in distance from the smelter, there was a corresponding decrease in plant injury and increase in the incidence of fungal diseases. Lilacs grown in polluted air (SO2) of New York City and other urban areas of USA developed substantially less infection of the powdery mildew fungus than the lilacs grown in rural areas (Hibben and Walker, 1966). Similar inhibitory effects of SO2 have also been reported in India (Rai, 1987; Khan et al., 1991). Incidence and severity of early blight of potato caused by Alternaria solani were low in fields about 3-4 km away from a coal fired thermal power plant compared to unpolluted area (Rai, 1987). Khan et al. (1991) observed a low intensity and incidence of powdery mildew caused by Sphaerotheca fulginea on cucurbits grown around ceramic and pottery industries at Khurja, India (upto 2 km left to Khurja-Delhi road). With increasing distance from the source, SO2 injury gradually decreased but incidence of powdery mildew increased.
Stimulation of pathogens: Some researchers have reported diseases in severe form on plants growing in SO2 stressed environments (Heagle, 1973). The incidence of Armillaria mellea was remarkably greater on trees injured by ambient SO2 (Kudela and Novakova, 1962; Donaubauer, 1968). An increased incidence of wood rot (Jancarik, 1961) and needle rot caused by Lophodemium piecae (Kudela and Novakova, 1962) was recorded in localities polluted with SO2. Chiba and Tanaka (1968) also observed high disease incidence of Rhizosphaera kalkhoffii on red pine in ambient condition. Greater infection of Trametes heteromorpha, T. serialis, Glocophyllum and Melampsora species was noticed on different trees grown in SO2 polluted areas (Scheffer and Hedgcock, 1955; Jancarik, 1961).
Plant diseases caused by nematodes: Plant parasitic nematodes constitute an important group of parasites that are considered as a constraint in the cultivation of agricultural crops. Plant parasitic nematodes are typically obligate parasites and can not survive for a longer duration in the absence of host plant. Majority of plant parasitic nematodes are root feeders but a few, like Anguina sp., Aphelenchoides spp., Ditylenchus spp. etc., attack and invade above ground parts such as stem, leaf or inflorescence. Plant nematodes on the basis of parasitism are categorized as ectoparasites and endoparasites. Ectoparasites feed on the surface of root or shoot whereas endoparasites penetrate the plant tissue and feed internally (Khan, 2008). Both groups of plant nematodes are agriculturally important, but endoparasites such as Meloidogyne spp. and Pratylenchus spp. are more damaging and inflict tremendous yield losses to susceptible crops. Effects of SO2 on the diseases caused by nematodes have been tested in some studies however, they are not adequate enough to categorize under simulated and ambient condition, hence are clubbed together and presented under following subheadings of inhibitory and stimulatory effects (Table 3).
Inhibition of nematode pathogenesis: Enough evidences exist to indicate that SO2 may inhibit parasitism of nematodes (Table 3). The effect of SO2 may be indirect either through altered host physiology or through some chemical alterations in soil as SO2 is unlikely to penetrate the soil and affect the inhabiting nematodes directly.
Bassus (1968) was probably the first to correlate nematode population to atmospheric pollution. He reported increased population of saprophagous and predaceous nematodes in the soil of pine trees severely damaged by SO2 and alkaline particulate material compared to the soil of those pines away from the source and exhibiting a mild injury of SO2. Weber et al. (1979) reported that intermittent exposures of soybean plants to 655 μg SO2 m-3 had adverse effects on Heterodera glycines and Paratrichodorus minor. Some more studies have also been conducted to evaluate the effect of coal smoke pollutants (primarily SO2) on the pathogenesis of nematodes (Khan and Khan, 2000). At a site, 2 km away from the stack of a coal fired thermal power plant having 233 (43-348) μg SO2 m-3 and 112 (29-174) μg NO2 m-3, disease intensity (galls) and reproduction (egg masses) of root-knot nematode, Meloidogyne incognita on tomato, were significantly low compared to plants grown at unpolluted site with same soil.
| Table 3: | Effect of sulphur dioxide with plant parasitic nematodes |
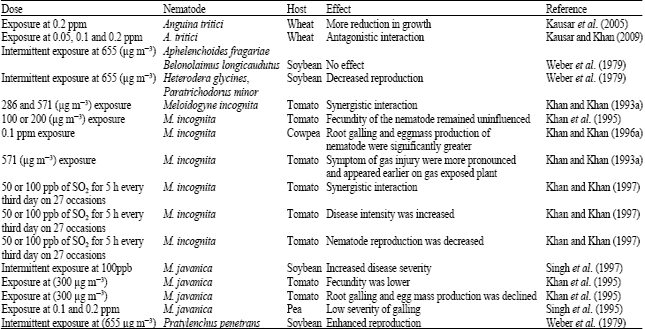 | |
Under artificial treatment condition in close top exposure chambers the reproduction of M. incognita on tomato was inhibited at 571 μg m-3 for 3 h every third day for 75 days. Severity of root-knot (galling) caused by M. javanica was also low on pea plants exposed to 0.1 and 0.2 ppm SO2 exposures (Singh et al., 1995). Root galling and egg mass production of M. javanica were declined by 23 and 24% at 300 μg SO2 m-3 exposure, respectively. The fecundity was 16% lower at 300 μg SO2 m-3 (Khan et al., 1995).
Kausar et al. (2005) observed the interaction of SO2 (0.2 ppm) and seed gall nematode, Anguina tritici on five wheat varieties. LOK-1, HD-2329, Raj-3765, PPW-154, UP-2328 in artificial treatment condition. SO2 alone significantly reduced plant growth of all the varieties. Reduction in growth was more in nematode inoculated and SO2 exposed sets compared to nematode alone-inoculated sets. However, highest reduction in yield was observed in nematode inoculated sets. Effect of different concentrations of SO2 (0.05, 0.1 and 0.2 ppm) on the mortality of seed gall nematode, A. tritici at different time intervals (1, 2, 3 and 4th days) has also been studied under artificial treatment condition (Kausar and Khan, 2009). All concentrations of SO2 were found harmful to juveniles of A. tritici. Further, investigation of the interactive effect of different concentration of SO2 and inoculum levels (2,500, 5,000 and 10,000) of nematode on wheat, Triticum aestivum revealed that plant growth, yield, photosynthetic pigments, seed carbohydrate and seed protein were reduced significantly. Both the pathogens and SO2 were antagonistically interacted.
Stimulation of nematode pathogens: Considerable numbers of reports have also revealed stimulatory effect of SO2 on the activity of plant parasitic nematode (Table 3). Weber et al. (1979) reported that intermittent exposures of soybean plants to 655 μg SO2 m-3 enhanced the reproduction of Pratylenchus penetrans, but Aphelenchoides fragariae and Belonolaimus longicaudatus remained unifluenced. SO2 also had stimulatory effect on the development and reproduction of Pratylenchus penetrans on tomato (Shew et al., 1982). Significantly greater number of galls and egg masses (reproduction) of M. incognita were recorded on eggplant (Khan and Khan, 1994b), okra (Khan and Khan, 1994a) and tomato (Khan and Khan, 1996b) grown in clay plots at a site about 1 km away from the stack of a coal fired thermal power plant, where daily mean concentration of SO2 and NO2 was 147 (16-269) and 88 (19-231) μg m-3, respectively. Interactive effects of root-knot nematodes and coal smoke were invariably synergistic, leading to a greater reduction of plant growth and yield. Similar stimulatory effects of coal smoke (containing higher levels of SO2), emanating from ceramic and pottery industries at Khurja on root-knot disease and reproduction of M. arenaria, M. incognita and M. javanica have been noticed on cucurbits (Khan et al., 1991). Gall formation and egg mass production of these nematode species gradually declined with increasing distance from the pottery units.
Khan and Khan (1993a) studied the interaction of SO2 and root-knot nematodes on tomato (cv. Pusa Ruby). In a close top exposure chambers intermittent exposures at 286 μg m-3 (3 h every third day for 75 days) induced considerably greater chlorosis with browning on the foliage of tomato plants infected with M. incognita race 1 (Khan and Khan, 1993b). Synergistic (positive) interactions between SO2 and M. incognita occurred in concomitant and post inoculation exposures at 286 and 571 μg m-3, respectively. Root galling and egg mass production of M. javanica were enhanced by about 11% and 6% at 100 μg m-3. The galling, egg masses production and fecundity remained unaffected at 200 μg SO2 m-3 (Khan et al., 1995). However, the root galling and egg mass production of M. incognita on cowpea cultivars were significantly greater on plants exposed to 0.1 ppm SO2 over control (Khan and Khan, 1996a, b).
Khan and Khan (1997) reported that in infected plants, the diffusive intake of gaseous pollutants would be greater and the interaction between the nematode and pollutant(s) would be governed by the degree of stomatal opening. Tomato plants infected with the root-knot nematode, M. incognita were exposed to air containing 0, 50 or 100 ppb of SO2 or O3 for 5 h every third day on 27 occasions in during two years. Plants exposed to the gases at 100 ppb had chlorotic and/or necrotic leaves, smaller shoots and roots, reduced leaf pigment levels and low yield, compared to untreated plants. Greater foliar injury developed on plants exposed to SO2 + O3 mixture. Gas injury symptoms were greater on nematode-infected exposed plants. M. incognita alone reduced tomato yield by 14.4% and induced a 3.6% increase in the width of stomatal pores and a 15.6% increase in the transpiration rate. A positive correlation was observed between stomatal pore width and rate of transpiration. Interaction between SO2 and O3 depended on the presence (significant) or absence (insignificant) of nematodes. Effects of nematode infection and gas exposures (especially mixtures) were mostly synergistic. Disease intensity (galls per root system) was increased, but nematode reproduction (egg masses per root system, eggs per egg mass) decreased on plants exposed to SO2 and/or O3.
Singh et al. (1997) evaluated the concomitant effects of SO2 on soybean in the presence or absence of root knot nematode and/or root nodule bacteria. The study revealed that root nodule bacteria enhanced the plant growth yield, pigment, protein, nitrogen and oil contents of soybean. However, these parameters were suppressed significantly by M. javanica. Joint inoculation of M. javanica and Bradyrhizobium japonicum caused reduction in concerned parameters, less than root nodule bacteria inoculated but more than nematode inoculated ones. Severity of the nematode disease was higher on 100 ppb exposed soybean plants.
CONCLUSION
Despite the deleterious consequences of burning of fossil fuels, coal is still being extensively used as a vital energy source for various industrial operations. Poor quality of coal, with regard to higher sulphur and ash contents and low calorific value, further aggravates pollution problems. Prevalence and phytotoxicity of SO2, have been realized well, but little efforts have been done in the developing countries to equip industries with pollution control devices so as to keep the ambient SO2 levels within the recommended limits. In India, majority of the power generating plants release SO2 at concentrations high enough to affect the surrounding vegetation. The cash crops, being usually grown in adjoining areas for easy access to market, may sustain pollution injuries due to expanded and green foliage (Khan et al., 1998; Khan and Khan, 2010).
In addition to causing direct damage, SO2 may also influence plant performance indirectly by influencing activity of microorganisms associated with the plant, especially pathogens as they draw nutrition form host plants. Researches conducted on the effect of SO2 on host parasite interaction have provided ample evidences that the pollutant may modify the host response and pathogenicity of the organism. In most of the cases, the gas has proved inhibitory to the fungus leading to suppression of the disease but in a few instances, SO2 promoted the disease severity and multiplication/reproduction of the pathogen. The varied response, however, has been found to be concentration dependent. Moderate levels of SO2 somewhere close to critical levels (80 ppb 24 h mean) have been found to synergise plant pathogenic fungi and nematodes whereas higher concentrations (above 100 ppb) caused adverse effects on the pathogens.
At the present state of our research and knowledge we are unable to convince ourself on the very basic question on variable response of pathogens to SO2 and the mechanism(s) involved in the interaction that governs the pollutant-pathogen relationships. To get convincing explanation it is greatly needed to devise and conduct experiments addressing these questions. Secondly, the host parasite relationships, where disease severity increases as a result of SO2 exposures, are required to be investigated more thoroughly. The report on greater susceptibility of plants to pathogens in SO2 stressed environment with lower concentration of SO2 such as 50-100 ppb of certain nematodes is alarming, though at this stage it is hard to make a generalization as the evidences are inadequate. More scientific efforts are needed to be diverted in this direction. If the predisposing effect of SO2 is established and it occurs invariably in the ambient condition, the critical levels of SO2 considered safer for plants has to be re-evaluated.
ACKNOWLEDGMENTS
Financial assistance from the Department of Science and Technology, Government of India, New Delhi through a research project entitled “Studies on the effect of sulphur dioxide and Alternaria species singly and jointly on the indigenous germplasm of mustard” (SP/SO/A-04/2001) is gratefully acknowledged which has greatly supported our overall research programme on pollutant-pathogen interaction.
REFERENCES
- Agrawal, M. and M. Verma, 1997. Amelioration of sulphur dioxide phytotoxicity in wheat cultivars by modifying NPK nutrients. J. Environ. Manage., 49: 231-244.
CrossRef - Cheah, L.H., A.W. Hunt and P. Lorentz, 1993. Effect of sulphur dioxide fumigation on botrytis storage rot and residues in kiwifruit. Postharvest Biol. Technol., 3: 11-16.
CrossRef - Clapperton, M.J. and D.M. Reid, 1994. Effects of sulphur dioxide (SO2) on growth and flowering of SO2 tolerant and non-tolerant genotypes of Phleum pratense. Environ. Pollut., 86: 251-258.
PubMedDirect Link - Clarkson, D.T., M.J. Hawkesford and J.C. Davidian, 1993. Membrane and Long Distance Transport of Sulphate. In: Sulphur Nutrition and Assimilation in Higher Plants: Regulatory Agricultural and Environmental Aspects, De Kok, L.J., I. Stulen, H. Rennenberg, C. Brunold and W.E. Rauser (Eds.). SPB Academic Publishing, The Hague, The Netherlands, pp: 3-19.
- Cullis, C.F. and M.M. Hirschler, 1980. Atmospheric sulphur: Natural and man-made sources. Atmos. Environ., 14: 1263-1278.
CrossRef - Darley, E.F. and J.T. Middleton, 1966. Problems of air pollution in plant pathology. Annu. Rev. Phytopathol., 4: 103-118.
Direct Link - De Kok, L.J., 1990. Sulphur Metabolism in Plants Exposed to Atmospheric Sulphur. In: Sulphur Nutrition and Sulphur Assimilation in Higher Plants: Fundamental, Environmental and Agricultural Aspects, Rennenberg, H., C. Brunold, L.J. De Kok and I. Stulen (Eds.). SPB Academic Publishing, The Hague, The Netherlands, pp: 111-130.
- Emberson, L.D., M.R. Ashmore, F. Murray, J.C.I. Kuylenstierna and K.E. Percy et al., 2001. Impacts of air pollutants on vegetation in developing countries. Water Air Soil Pollut., 130: 107-118.
CrossRefDirect Link - Haggestad, H.E., J.H. Bennett, E.H. Lee and L.W. Douglass, 1986. Effects of increasing doses of sulphur dioxide and ambient ozone on tomatoes: Plant growth, leaf injury, elemental composition, fruit yields and quality. Phytopathol., 76: 1338-1344.
Direct Link - Heagle, A.S., 1973. Interaction between air pollutants and plant parasites. Annu. Rev. Phytopathol., 11: 365-388.
Direct Link - Hibben, C.R. and M.P. Taylor, 1975. Ozone and sulphur dioxide effects on the lilac powdery mildew fungus. Environ. Pollut., 9: 107-114.
CrossRef - Kausar, S., A.A. Khan and D. Raghav, 2005. Interaction of sulphur dioxide and seed gall nematode, Anguina tritici, on wheat. J. Food Agric. Environ., 3: 130-132.
Direct Link - Khan, M.R. and M.W. Khan, 1993. The interaction of SOM2 and root-knot nematode on tomato. Environ. Pollut., 81: 91-102.
CrossRef - Khan, M.R. and M.W. Khan, 1994. Single and interactive effects of root-knot nematode and coal smoke on okra. N. Phytol., 126: 337-345.
Direct Link - Khan, M.R. and M.W. Khan, 1994. Smoke pollution and the effects of root-knot nematode on the growth of eggplants. Plant Pathol., 43: 683-693.
CrossRefDirect Link - Khan, M.R. and M.W. Khan, 1996. Interaction of Meloidogyne incognita and coal smoke pollutants on tomato. Nematropica, 26: 47-56.
Direct Link - Khan, M.R. and M.W. Khan, 1997. Effect of root-knot nematode, Meloidogyne incognita, on the sensitivity of tomato to sulphur dioxide and ozone. Environ. Exp. Bot., 38: 117-130.
CrossRef - Khan, M.R., M.W. Khan and M.J. Pasha, 1998. Effects of sulphur dioxide on the development of powdery mildew of cucumber. Environ. Exp. Bot., 40: 265-273.
CrossRef - Khan, M.W. and M. Kulshrestha, 1991. Impact of sulphur dioxide exposure on conidial germination of powdery mildew fungi. Environ. Pollut., 70: 81-88.
CrossRef - Lorenzini, G., A. Panicucci and L. Guidi, 1990. Growth dynamics of wheat (Triticum aestivum L.) exposed to sulphur dioxide pollution. Bull. Environ. Contaminat. Toxicol., 45: 408-414.
CrossRefDirect Link - Magan, N. and A.R. McLeod, 1991. Effect of open-air fumigation with sulphur dioxide on the occurrence of phylloplane fungi on winter barley. Agric. Ecosyst. Environ., 33: 245-261.
CrossRef - Magan, N. and A.R. McLeod, 1988. In vitro growth and germination of phylloplane fungi in atmospheric sulphur dioxide. Trans. Br. Mycol. Soc., 90: 571-575.
CrossRef - Mansfield, P.J., J.N.B. Bell, A.R. McLeod and B.E.J. Wheeler, 1991. Effects of sulphur dioxide on the development of fungal diseases of winter barley in an open-air fumigation system. Agric. Ecosyst. Environ., 33: 215-232.
CrossRef - Rakwal, R., G.K. Agrawal, A. Kubo, M. Yonekura, S. Tamogami, H. Saji and H. Iwahashi, 2003. Defense/stress response elicited in rice seedlings exposed to the gaseous air pollutant sulphur dioxide. Environ. Exp. Bot., 49: 223-235.
CrossRef - Saxe, H., 1983. Long term effects of low levels of SO2 on bean plants (Phaseolus vulgaris). I. Emission response pattern of net photosynthesis and transpiration during life long continuous measurements. Physiol. Plant., 57: 101-107.
CrossRefDirect Link - Saxe, H., 1983. Long term effects of low levels of SO2 on bean plants (Phaseolus vulgaris). II. Emission response effects on biomass production: Quantity and quality. Physiol. Plant., 57: 108-113.
CrossRefDirect Link - Sharp, E.L., 1967. Atmospheric ions and germination of uredospores of Puccinaia striiformis. Sci., 156: 1359-1360.
PubMedDirect Link - Shaw, P.J.A., M.R. Holland, N.M. Darrall and A.R. McLeod, 1993. The occurrence of SO2-related foliar symptoms on Scots pine (Pinus sylvestris L.) in an open-air forest fumigation experiment. N. Phytol., 123: 143-152.
Direct Link - Singh, K., M.W. Khan and M.R. Khan, 1996. Plant growth and root-knot disease of soybean under the stress of SO2 and O3 gases mixture. Nematologia Mediterranea, 24: 261-264.
Direct Link - Skye, E., 1968. Lichens and air pollution: A study of cryptogamic epiphytes and environment in the Stockholm region. Acta Phytogeogr. Suecica, 52: 119-123.
Direct Link - Takemoto, B.K. and R.D. Noble, 1982. The effects of short term SO2 fumigation on photosynthesis and respiration in soybean Glycine max. Environ. Pollut. Ser. A, 28: 67-74.
CrossRef - Thomas, M.D., R.H. Hendrick, T.R. Collier and G.R. Hill, 1943. The utilization of sulphate and sulphur dioxide for the nutrition of alfalfa. Plant Physiol., 18: 345-371.
Direct Link - Weinstein, L.H., D.C. McCune, A.I. Aluisio and P. Lenken, 1975. The effect of sulphur dioxide on the incidence and severity of bean rust and early blight of tomato. Environ. Pollut., 9: 145-155.
CrossRef - Yang, L., I. Stulen and L.J. De Kok, 2006. Sulphur dioxide: Relevance of toxic and nutritional effects for Chinese cabbage. Environ. Exp. Bot., 57: 236-245.
CrossRef