
V. Pensuk
Program in Plant Production Technology, Faculty of Technology, Udonthani Rajabhat University, Udonthani 41000, Thailand
S. Jogloy
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
A. Patanothai
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
Plant Pathology Journal
Year: 2010 | Volume: 9 | Issue: 4 | Page No.: 188-193







ABSTRACT
The difference in PBND incidence in the rainy and dry seasons in Thailand has led to a hypothesis that temperature and relative humidity might be the causal factors. The objective of this study was to investigate the effects of temperature and relative humidity on the effectiveness of mechanical inoculation of PBNV on peanut. Two experiments were conducted in the rainy and dry seasons during 2004 to 2005, using the susceptible peanut genotype Tainan 9. The first experiment evaluated the effect of low temperature on the effectiveness of PBNV transmission by mechanical inoculation. The second experiment compared two temperature and relative humidity conditions in the rainy season and in the dry season. The results showed that exposing the PBNV-inoculated plants to low temperature (25°C, 90% RH) during a 12 h light period and 20°C, 90% RH during a 12 h dark period) for all the time during the experimental period did not increase the infected plants. Two climate conditions representing the dry season (daytime, 32°C, 44% RH, night-time, 22°C, 65% RH) and the rainy season (daytime, 36°C, 46% RH, night-time, 25°C, 70% RH) in Thailand also showed no difference in the infected plants. These results indicated that low temperature or relative humidity did not affect the transmissibility of PBNV. The difference in disease incidences in the rainy and the dry seasons in Thailand could not be explained by the difference in climatic conditions but could possibly be accounted for by the difference in vector infestation.
PDF Abstract XML References Citation
Received: July 06, 2010;
Accepted: August 16, 2010;
Published: November 12, 2010
How to cite this article
V. Pensuk, S. Jogloy and A. Patanothai, 2010. Effects of Temperature and Relative Humidity on the Effectiveness of Peanut bud necrosis virus Inoculation on Peanut. Plant Pathology Journal, 9: 188-193.
DOI: 10.3923/ppj.2010.188.193
URL: https://scialert.net/abstract/?doi=ppj.2010.188.193
DOI: 10.3923/ppj.2010.188.193
URL: https://scialert.net/abstract/?doi=ppj.2010.188.193
INTRODUCTION
Peanut bud necrosis disease (PBND) caused by Peanut bud necrosis virus (PBNV) and transmitted by Thrips palmi Karny, is a serious disease of peanut in many countries in Asia (Satyanarayana et al., 1996; Reddy et al., 1995). The disease can cause substantial yield losses in peanut (Dwivedi et al., 1995). PBNV is now recognized as a distinct species in the genus Tospovirus (Reddy et al., 1992; Van Regenmortel et al., 2000). Currently, there is no practical control measure for PBNV on peanut. However, the disease incidence can be reduced by some cultural practices such as adjustment of planting date to the period with low levels of vector activity, intercropping with fast growing cereals (Reddy et al., 2000) and close spacing (Basu, 1995; Buiel, 1996; Wongkaew, 1995). Irrigation may effect the distribution of thrips and consequently can reduce PBND incidence (Bhatnagar et al., 1995). Insecticide application to control the vector failed to control the disease and the rapid acquisition of resistance to insecticides by thrips had been observed (Boiteux et al., 1993). Therefore, cultivar resistance appears to be the most effective strategy for control disease.
Successful inoculation of PBNV is necessary to evaluate the levels of resistance of genotypes. Although, field screening of peanut genotypes for resistance to the disease has been reported, a low disease incidence of a genotype in field screening could not be discerned whether it was due to the effect of resistance to the virus or resistance to the vector or the collective effect of both (Dwivedi et al., 1995). Therefore, an efficient method for mechanical transmission of the virus is required in breeding for PBNV resistance. Sap inoculation is a standard method for resistance evaluation. However, the method has been reported to yield different results for the same genotype. Dwivedi et al. (1995) reported PBND incidences on a susceptible check in the range of 78-94% by mechanical inoculation. Buiel and Parlevliet (1996) studied the occurrence of mature plant resistance and found the incidences between 12 and 96% on a susceptible peanut genotype depending on the age of the plants. In, screening peanut genotypes for resistance to PBNV, Pensuk et al. (2002) obtained the transmission efficiency between 60-100% on susceptible genotypes by the mechanical inoculation under greenhouse conditions. The inconsistency of results from mechanical inoculation has made the PBNV resistance evaluation less efficient and has caused difficulties in the genetic studies of PBNV resistance. To improve the efficiency of mechanical transmission of the virus, a better understanding of the factors causing the variation of the results is needed.
Mandal et al. (2001) reported that the common factors causing the variability in mechanical transmission of Tomato spotted wilt virus (TSWV) were the inoculum source, the buffer composition, the additives used in the preparation of inoculum, the growth stage of assay plants, the methods of inoculum application to the assay plants and the environment after inoculation. These factors were assumed to cause the variability in mechanical transmission of PBNV as well. However, in the studies of Pensuk et al. (2002), the same source of inoculum, the same method of inoculum preparation, the same growth stage of the assay plants and the same method of inoculation were used. Yet, the results were still variable and a near 100% disease incidence could not be achieved at will. It was hypothesized that environmental conditions would have been the cause of the variability in PBNV transmission. In Thailand, it was also found that the incidence of the disease was much lower in the rainy season than in the dry season (Wongkaew, 1995). Because crop grown in the dry season experiences low temperature and low relative humidity at the early growth stage, these factors were suspected to favor disease development resulted in a high disease incidence in the dry season. This study aimed to investigate the effects of temperature and relative humidity on the effectiveness of PBNV inoculation on peanut in order to develop a protocol that enhances the efficiency and reliability of mechanical transmission of the virus.
MATERIALS AND METHODS
This study consisted of two experiments in the rainy and dry seasons during 2004 to 2005. The first experiment (Experiment 1) evaluated the effect of low temperature on the effectiveness of PBNV transmission by mechanical inoculation. The second experiment (Experiment 2) compared two temperature and relative humidity conditions that represented the conditions in the rainy season during 8 September 2004 and the dry season during 25 February 2005 in Thailand on the success of mechanical transmission of PBNV to peanut.
In both studies, the susceptible peanut genotype Tainan 9 was used. The test plants were grown in plastic pots and filled with a soil mixture (soil, sand and compost at 2:1:1 v/v/v). Each pot contained four plants. Each treatment consisted of 3 or 4 replicates depending on the trial, with 8 plants (two pots) per replicate. The test plants were inoculated at 7 days after planting. Two new unexpanded quadrifoliate leaves were inoculated per plant.
In preparing the inoculum, leaves of infected peanut plants showing typical symptom of PBND were initially collected from a field. A virus clone was then isolated from a single lesion that appeared as primary symptom on an infected peanut plant. The isolated virus was propagated in Tainan 9 peanut plants that were kept in a screened house. Identity of the virus was confirmed by ELISA test using PBNV antiserum as a reference (Do Nascimento et al., 2006). The inoculum was prepared by grinding systemically infected peanut leaves in 0.05 M phosphate buffer, pH 7.0, containing 0.2% 2-mercaptoethanol (1:10 w/v) using a chilled pestle and mortar. Debris was removed by squeezing the ground extract through a pad of nonabsorbent cotton. Celite was added to the plant extract to a final concentration of 1%. The extract was kept chilled during inoculation. Inoculation was done by dripping 200 μL of the inoculum onto the unfolding leaves of each plant and rubbed thoroughly. Prior to inoculation, the plants were maintained under dark condition for 3 h. The plants were sprayed with distilled water after inoculation.
For experiment 1, the treatments included maintaining the inoculated plants in an environmental growth chamber (Contherm Model CAT 610) that was set at 25°C, 90% RH during a 12 h light period (a light intensity of 1,000 lk) and 20°C, 90% RH during a 12 h dark period and outside the growth chamber at room temperature. The test plants maintained in the growth chamber were divided into 4 groups by the length of time they were kept in the growth chamber after inoculation, i.e., 24, 48, 72 h and all the period after inoculation until the test was terminated. The plants were visually scored for PBNV symptoms at 14 and 28 days after inoculation. Those showing systemic symptoms were considered as infected plants. At 28 days after inoculation, a newly formed leaflet from each plant was assayed by direct antigen coating enzyme-linked immunosorbent assay (DAC-ELISA) as described by Wongkaew (1993) to confirm PBNV infection. The primary antiserum specific to PBNV was obtained from S. Wongkaew (Khon Kaen University, Thailand) and was cross absorbed with healthy plant extract at a 1:1000 (v/v) dilution. Goat-anti-rabbit IgG conjugated to alkaline phosphatase (Sigma 99H9235) at a dilution of 1:2000 was used as the secondary antibody. Extracts from healthy and infected Tainan 9 peanut leaves were used as the negative and positive controls, respectively. Absorbance (405 nm) at least 2 folds higher than those of the healthy control was considered positive for the virus. The data were obtained using an ELISA plate reader Model Microplate Manager® 4.0 Bio-Rad Laboratories, Inc. The experiment was repeated 5 times. Trail II was conducted in 3 replicates, while other trials were conducted in 4 replicates. The disease incidence was determined as the infected plants. Data were analyzed statistically after being transformed by an arc sine transformation.
In experiment 2, the inoculated plants were kept in two growth chambers that were set to the conditions representing the average climatic conditions in the rainy and the dry seasons in Thailand, respectively. For the condition representing the dry season, the growth chamber was set to 32°C, 44% RH, during an 11 h 30 min light period and 22°C, 65% RH, during a 12 h 30 min dark period. For the condition representing the rainy season, the environmental growth chamber was set to 36°C, 46% RH, during a 12 h 30 min light period and 25°C, 70% RH, during an 11 h 30 min dark period. The inoculated plants were kept in the growth chambers for four weeks then were visually checked for PBNV symptoms. Only plants showing clear systemic symptoms were considered to be infected. The experiment was repeated 3 times, all with 4 replicates. The disease incidence was determined as the percentage of infected plants and the differences between means of the two treatments were test statistically by t-test.
RESULTS
The experiment 1 showed no difference among treatments in the number of infected plants (Table 1), this indicated that keeping PBNV inoculated plants in low temperature for a certain period did not increase the percentage of infected plants over the conditions of room temperatures at different times of the year. Variations in the percentages of infected plants were still obtained in different trials. High percentages of infected plants were obtained in Trials II and III, i.e., 100% for all treatments in Trial II and 88 to 100% in Trial III, but lower percentages were showed in other trials. Trials I, II and V were conducted during the late-rainy season while Trials III and IV were conducted in the rainy seasons. The temperatures and relative humidity under room conditions in these trials also differed, so as the percentages of infected plants under room conditions. However, no association was observed between the temperature and relative humidity under room condition with the percentage of PBNV infected plants. The same was found for the treatments where the inoculated plants were kept under low temperature in the growth chamber for a certain period. In the treatment in which the inoculated plants were kept under the same control condition all the time after inoculation, the PBNV infected plants still varied in the different trials. These results indicated that, temperature and relative humidity were not the factor causing the variability of PBNV transmission by mechanical inoculation.
Experiment 2 was performed to compensate for the fact that in experiment 1 the environmental conditions outside the growth chamber could not be controlled and these conditions might affect the symptom expression of the disease.
| Table 1: | Incidences of PBNV infected plants obtained from mechanical inoculation under different environmental conditions |
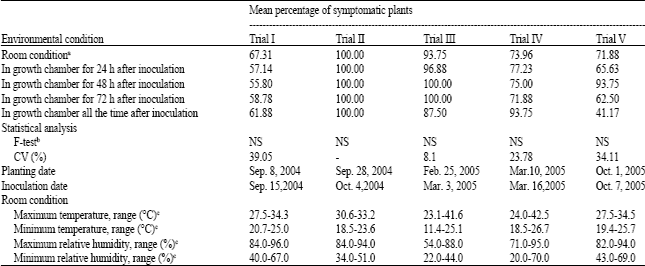 | |
| aGrowth chamber condition: 25°C, 90% RH during a 12 h light period and 20°C, 90% RH during a 12 h dark period. bData in trial I, IV and V were transformed using the arcsine transformation before analysis; NS = Not significant.cData were recorded during the experimental period, i.e., 28 days after inoculation | |
| Table 2: | Incidences of PBNV infected plants obtained from mechanical inoculation under the environmental conditions representing the conditions in rainy and the dry seasons in Thailand |
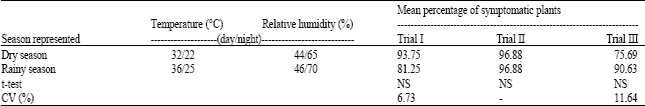 | |
| NS: Not significant | |
In this experiment, the environmental conditions representing the average temperature and relative humidity conditions in the rainy season and the dry season were set up using growth chambers (Table 2). The results from three repeated trials showed that the two temperature and relative humidity conditions did not cause a difference in the number of symptomatic plants. Again, variations in the infected plants were observed among trials. The warm and humid condition representing the rainy season tended to produce less symptomatic plants than the cool and less humid condition representing the dry season in Trial I, but the opposite was observed in Trial III. In Trial II, however, 97% infection was obtained in both conditions. These results also confirmed the results of experiment I in that temperature and relative humidity were not the causal factor for the variability in PBNV transmission by mechanical transmission.
DISCUSSION
Little is known about the interactions between the environments and the symptomatic expression of the recently described PBNV that is now recognized as a member of the genus Tospovirus. PBND in Thailand is prevalent mainly during the dry season (Wongkaew, 1995), thus, we hypothesized that the environmental factors prevailing in dry season might favor the disease incidence. To test this hypothesis, the inoculated plants were exposed to a warm temperature, i.e., under room condition and a cooler temperature, i.e., under growth chamber condition, after inoculation. Different times of incubating the inoculated plants in the growth chamber were also examined, as, if it works, the shortest incubating time would be preferred to speed up the process of peanut breeding lines evaluation for PBNV resistance. However, the disease incidences obtained from all treatments were not significantly different in all the 5 repeated trials. These results indicated that low temperature did not favor the PBNV transmission. These results differed somewhat from the study of Mandal et al. (2001) on the transmission of TSWV. They reported that the maximum rate of transmission of TSWV from tobacco to peanut achieved at 15 days postinoculation (DPI) under greenhouse condition was 66.6%, whereas a 100% transmission was achieved by 10 DPI under growth chamber condition. Llamas-Llamas et al. (1998) studied the effect of temperature on symptom expression and accumulation of TSWV in Nicotiana tabacum and reported that, in general, virus accumulation in the inoculated leaves of all plants was higher at a lower temperature. Llamas-Llamas et al. (1998) also reported that TSWV replication was higher at 20°C than at 36°C, but the disease symptoms were more severe at 36°C. These finding has led to their conclusion that there is not necessarily a direct relationship between virus accumulation and symptom expression.
Since, low temperature may not be the only factor that favors bud necrosis incidence, a second experiment was conducted in which both temperature and relative humidity that characterize the average conditions of the rainy and the dry seasons in Thailand were controlled. The inoculated plants were kept in the growth chamber throughout the experiment to avoid exposing the plants to varying environmental conditions. The results in three repeated trials indicated that the disease incidence was not affected by the environmental conditions under which the inoculated plants were kept. Variation in the percentages of infected plants among the trials also could not be accounted for by temperature and relative humidity as the same conditions of the two treatments were maintained in all the 3 repeated trials. The difference in the incidences of PBND in the rainy and the dry seasons in peanut production areas in Thailand, therefore, could not be accounted for by the difference in temperature and relative humidity in the two seasons. However, Tsai et al. (1995) reported that T. palmi was able to tolerate a low temperature (56% mortality when held at 0°C for 15 h) much better than a high temperature (100% mortality at 45°C for 15 h). Tsai et al. (1995) also showed that T. palmi could not complete its development at 35°C or higher temperatures. This might partially explain why T. palmi populations are higher in the winter and spring and lower in the summer in Florida (Tsai et al., 1995). As temperatures during the rainy season in Thailand are generally above 35°C, T. palmi populations in peanut production areas of the country are expected to be low and this could account for the low PBND incidence in these areas.
The results of the present study indicated that low temperature or low relative humidity did not affect the transmissibility of PBNV by mechanical inoculation. A 100% PBND incidence from mechanical inoculation was obtained in some trials but not the others. The less than perfect infection by mechanical inoculation might be attributed to the disease escape and/or the subliminal infection as suggested by Do Nascimento et al. (2006). The low PBND incidence occurring in the rainy season in Thailand could not be accounted for by the high temperature and high relative humidity conditions prevailing in the season, but might be explained by the low vector population and hence, low vector infestation in the rainy season.
ACKNOWLEDGMENTS
This study was funded by the Senior Research Scholar Project of Prof. Dr. A. Patanothai under the Thailand Research Fund and the work was carried out with the assistance of the Peanut Project, Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khon Kaen University, Khon Kaen, Thailand.
REFERENCES
- Bhatnagar, H., M.V. Reddy, J.N. Rao and P. Singh, 1995. Confirmation of the effects of plant density and irrigation on peanut bud necrosis incidence. Int. Arachis Newslett., 15: 52-53.
Direct Link - Boiteux, L.S., T. Nagata, W.P. Dutra and M.E.N. Fonseca, 1993. Sources of resistance to Tomato spotted wilt virus (TSWV) in cultivated and wild species of Capsicum. Euphytica, 67: 89-94.
CrossRef - Buiel, A.A.M. and J.E. Parlevliet, 1996. Mature plant and tissue resistance in the Groundnut-Peanut bud necrosis virus system. Euphytica, 91: 213-217.
Direct Link - Dwivedi, S.L., S.N. Nigam, D.V.R. Reddy, A.S. Reddy and G.V. Ranga Rao, 1995. Progress in Breeding Groundnut Varieties Resistant to Peanut Bud Necrosis Virus and its Vector. In: Recent Studies on Peanut Bud Necrosis Disease, Buiel, A.A.M., J.E. Parlevliet and J.M. Lenne (Eds.). International Crops Research Institute for the Semi-Arid Tropics, Patancheru, India, pp: 35-40.
- Llamas-Llamas, M.E., E. Zavaleta-Mejia, V.A. Gonzales-Hermandez, L. Cervates-Diaz, J.A. Santizo-Rincon and D.L. Ochoa-Martinez, 1998. Effect of temperature on symptom expression and accumulation of Tomato spotted wilt virus in different host species. Plant Pathology., 47: 341-347.
CrossRefDirect Link - Mandal, B., H.R. Pappu and A.K. Culbreath, 2001. Factors affecting mechanical transmission of Tomato spotted wilt virus to peanut (Arachis hypogaea). Plant Dis., 85: 1259-1263.
CrossRef - Pensuk, V., N. Daengpluang, S. Wongkaew, S. Jogloy and A. Patanothai, 2002. Evaluation of screening procedures to identify peanut resistance to Peanut bud necrosis virus (PBNV). Peanut Sci., 29: 47-51.
CrossRef - Reddy, D.V.R., A.A.M. Buiel, T. Satyanarayana, S.L. Dwivedi and A.S. Reddy et al., 1995. Peanut Bud Necrosis Disease: An Overview. In: Recent Studies on Peanut Bud Necrosis Disease, Buiel, A.A.M., J.E. Parlevliet and J.M. Lenne (Eds.). International Crops Research Institute for the Semi-Arid Tropics, Patancheru, India, pp: 3-7.
Direct Link - Reddy, D.V.R., A.S. Ratna, M.R. Sudarshana, F. Poul and I. Kiran Kumar, 1992. Serological relationship and purification of bud necrosis virus, a tospovirus occurring in peanut. Ann. Applied Biol., 120: 279-286.
CrossRef - Reddy, A.S., L.J. Reddy, N. Mallikarjuna, M.D. Abdurahman, Y.V. Reddy, P.J. Bramel and D.V.R. Reddy, 2000. Identification of resistance to Peanut bud necrosis virus (PBNV) in wild Arachis germplasm. Ann. Applied Biol., 137: 135-139.
Direct Link - Satyanarayana, T., S.E. Mitchell, D.V.R. Reddy, S. Brown and S. Kresovich et al., 1996. Peanut bud necrosis virus, purification of nucleocapsids and sequence homology of nucleocapsid protein and glycoprotein precursor with other tospovirus. Acta Hortic., 431: 228-236.
Direct Link - Tsai, J.H., B. Yue, S.E. Webb, J.E. Funderburk and H.T. Hsu, 1995. Effects of host plant and temperature on growth and reproduction of Thrips palmi (Thysanoptera: Thripidae). Environ. Entomol., 24: 1598-1603.
Direct Link - Van Regenmortel, M.H.V., C.M. Fauquet, D.H.L. Bishop, E.B. Carstens and M.K. Estes et al., 2000. Virus Taxonomy: Classification and Nomenclature of Viruses: Seventh Report of the International Committee on Taxonomy of Viruses. 1st Edn., Academic Press, New York, ISBN-10: 0123702003.
Direct Link - Do Nascimento, L.C., V. Pensuk, N.P. Da Costa, F.M. De Assis Filho, G. Pio-Ribeiro, C.M. Deom and J. Sherwood, 2006. Evaluation of peanut genotypes for resistance to Tomato spotted wilt virus by mechanical and thrips inoculation. Pesq. Agropec. Bras., 41: 937-942.
Direct Link
Dr Suresh Kunkalikar Reply
Good observation. We too have obtained similar results during our work on GBNV since last 7-8 years at MAHYCO research center, India. In rainy season disease was more on mechanically inoculated plants possibly due to temp of 27-30 dgrees and fairly high humidity.