
K.M. Abd El-Hai
Department of Leguminous and Forage Crops Diseases, Egypt
M.A. El-Metwally
Department of Mycological Research and Plant Diseases Survey,Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
S.M. El-Baz
Department of Mycological Research and Plant Diseases Survey,Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
Plant Pathology Journal
Year: 2010 | Volume: 9 | Issue: 4 | Page No.: 149-161







ABSTRACT
The causal pathogens of root rot and stalk rot of soybean were isolated from infected plants. Fusarium oxysporum, F. solani, Macrophomina phaseolina and Rhizoctonia solani were the main causal pathogens of pre- and post-emergence damping-off. The isolated fungi especially R. solani caused anatomically deformation in the basal portion of infected stem including complete disruption in epidermal cells and degradation of primary cell wall in cortical cells. The growth substances; Ethrel, Indole Butyric acid (IBA) and Cycocel (CCC) were tested in vitro on the growth of the pathogenic fungi. Ethrel at 200 ppm was the most effective in reducing fungal growth. During two successive growing seasons (2008 and 2009) of soybean under different salinity stress (1000 to 3000 ppm), the application of growth substances led to significant reduction in pre- and post-emergence damping-off as well as root and stalk rot diseases. Ethrel, CCC and IBA increased significantly branches number plant-1, photosynthetic pigments, total phenol, proline content, yield and seed quality (oil and protein %). Ethrel and CCC at 200 ppm were the best treatments. Soaking soybean seed in Ethrel, CCC or IBA at 200 ppm is recommended to be incorporated into the production program of soybean to decrease root and stalk rots and increase productivity and seed quality under salt stress conditions.
PDF Abstract XML References Citation
Received: May 27, 2010;
Accepted: July 14, 2010;
Published: May 08, 2019
How to cite this article
K.M. Abd El-Hai, M.A. El-Metwally and S.M. El-Baz, 2010. Reduction of Soybean Root and Stalk Rots by Growth Substances Under Salt Stress Conditions. Plant Pathology Journal, 9: 149-161.
DOI: 10.3923/ppj.2010.149.161
URL: https://scialert.net/abstract/?doi=ppj.2010.149.161
DOI: 10.3923/ppj.2010.149.161
URL: https://scialert.net/abstract/?doi=ppj.2010.149.161
INTRODUCTION
Soybean (Glycine max L. Merrill) is one of the main fabaceous crops all over the world. It is an important crop providing the abundant proteins and vegetable oils for human nutrition. Its highly digestible oil contains unsaturated fatty acids and no cholesterol. Its protein content is superior in substantial levels of most essential amino acids. The carbohydrate content is about 30%, with total soluble sugars equal to about 10%. Also, soybean can be considered as a friendly crop to the environment related to its supply of the soil by nitrogen fixation. Additionally, it is a good dietary source of calcium and phosphorus that is why soybean is a remarkable plant (Singh et al., 1987; Nassiuma and Wasike, 2002; Akande et al., 2007).
Salinity and soil-borne fungal diseases are limiting factors of soybean growth and productivity. Soybean has been recognized as a salt-sensitive plant according to the relative growth ratio under high-salt condition (Greenway and Munnus, 1980). High levels of salinity significantly reduce germination, seedling growth, shoot and root growth, photosynthetic pigments, protein content and productivity for many plant species including soybean (Muthukumarasam and Panneerselvam, 1997; Wang et al., 2001), because of the high-salt environments cause molecular damages as hyperosmotic stress, homeostasis disruption and ionic toxicity to plant cell that become a resultant limiting factor for plant development and crop productivity (Aoki et al., 2005).
Root rot and charcoal rot diseases are among the most destructive diseases attacking soybean seeds, seedlings and roots as well as lower part of the stem causing serious damage (Hassanien, 1985; Singh et al., 1987; Sinclair and Backman, 1989). Rhizocronia solani Kuchn causes foot and root rots of young soybean plants. Signs of infection are reddish-brown decay of the outer cortical tissues and later sunken reddish cankers that girdle the stem at the soil line (Sinclair and Backman, 1989). Moreover, charcoal rot caused by Macrophomina phaseolina (Tassi) Goid causes damping-off of seedlings, which characterized by reddish-brown lesions girdling the stem near the soil. The fungus turns stellar tissues of the root to dark brown and as the infected plant approach senescence, numerous blackish microsclerotia of the fungus from below the epidermis of the underground parts, giving the affected portion a charcoal appearance (Oyekan and Niak, 1987). Fusarium species have been reported to cause wilt in soybean, which characterized by the browning of the vascular tissues of roots and stems of infected plants and yellowing and withering of the foliage (Sinclair and Backman, 1989).
Many authors studied the effect of growth substances on overcoming the depressing effect of salinity (Zaidi and Singh, 1995; Galley and Fletcher, 1997; Abd El-Hai, 2001) and on linear growth, sporulation and sclerotial formation of the pathogens and they reported that auxins (IAA, IBA and NAA) are potential antifungal (Michiewiez and Rozej, 1987; Khalifa, 2003; Metwally et al., 2006). Under such adverse conditions, plants are more susceptible and secreted low yield.
Therefore, reduction of soybean root and stalk rots under salt stress conditions by means of growth substances, i.e., Ethrel, Indole Butyric acid (IBA) and Cycocel (CCC) was the objective of the present investigation.
MATERIALS AND METHODS
Isolation, purification and identification of the causal pathogens: Samples of soybean plant showing typical symptoms of root rot and stalk rot diseases were collected from the farm of Tag El-Ezz Agricultural Research Station, Dakahlia, Egypt during the summer season of 2007. The infected stem lower parts and roots were washed with tap water and cut into pieces, surface sterilized by immersing them in sodium hypochlorite (2%) for 2 min. The sterilized pieces rewashed several times with sterilized water, dried between two sterilized filter paper, placed on Potato Dextrose Agar (PDA) plates supplemented with streptomycin sulfate (100 μg mL-1) and incubated at 28°C for 5 days. The growing fungi were purified according to Hawker (1960). Identification of the isolated fungi was carried out based on taxonomic criteria as described by Ellis (1976) for Macrophomina phaseolina, Booth (1977) for Fusarium solani and F. oxysporum and Sneh et al. (1992) for Rhizoctonia solani.
Preparation of the fungal inoculum: The individual fungal inoculum of each fungus was prepared using sorghum: coarse sand: water (2:1:2 v/v) medium. The ingredients were mixed, bottled and autoclaved for 15 min at 1.5 air pressure. The sterilized medium was inoculated using agar discs obtained from the periphery of 6 days old colony of the isolated fungi. The inoculated media were incubated at 28°C for 15 days and were then used for soil infestation in the greenhouse experiment.
Greenhouse experiment
Pathogenicity test: Pathogenicity test of the isolated fungi was carried out under greenhouse. Each pathogen was added to the autoclaved clayey soil at a rate of 0.3% (w/w), then putted in sterilized pots (25 cm in diameter) filled up with 4 kg soil. The infested pots were watered daily for one week to enhance growth and distribution of the fungal inoculum. At the same time three pots containing autoclaved soil were prepared to serve as control. Seeds of soybean (Giza 22) were surface sterilized by immersing them in 0.1% sodium hypochlorite for two minutes then washed several times with sterilized water and sown at the rate of ten seeds pot-1, three replicates were used in each particular treatment. Disease assessment was recorded as pre and post-emergence damping-off as well as healthy survival plants after 20, 40 and 80 days from sowing, respectively.
Effect of soil-borne fungi on the anatomical structure of soybean seedlings: Specimens of soybean seedlings were chosen after 30 days from sowing for determination the anatomical changes which occurred by the pathogenic fungi. Small parts, 5 mm in length, were taken from the stem base (infected region). Specimens were killed and fixed in formalin: alcohol: acetic acid mixture (1:18:1 v/v), washed and dehydrated in alcohol series and embedded in paraffin wax (52-54°C m.p.). Sections at 12-15 μ thick were prepared by a rotary microtome, stained in crystal violet and erythrosine, cleared in xylol and mounted in Canada balsam (Gerlach, 1977). Sections were examined microscopically.
Effect of growth substances on the linear growth of pathogenic fungi: Ethrel (2-chloroethyl phosphonic acid) obtained from BDH chemicals LTD, U.K, indole butyric acid 99% (Alderich chemical company, England) and Cicocel 40% (Alderich chemical company, England) at 100 and 200 ppm as well as Kocide 101 77% w.p. and rizolex-T50 w.p. at 3 g L-1 were tested in vitro on the linear growth of the pathogenic fungi. Different concentrations were added to 10 mL of sterilized PDA before solidification and then poured in sterile petri-dishes. After solidification the plates were inoculated with fungal disc (5 mm) in the center of the plate and incubated at 27±1°C. Three plates for each particular treatment for each fungus were used as replicates; three plates were prepared to serve as control for each fungus. Linear growth was observed daily and diameter of fungal colonies were recorded when plates of any treatment were filled with the fungal growth.
Field experiments: Field experiments were carried out during 2008 and 2009 Summer seasons at Tag El-Ezz Research Station, Dakhalia Governorate, Egypt. On the base of soil salinity by measuring the Electrical conductivity (EC), the farm soil was divided into three blocks (1000, 2000 and 3000 ppm, approximately). Soybean seeds were soaked for 20 min in each tested growth substance (100 or 200 ppm) while, both fungicides (Kocide 101 or Rizolex T-50) were used as seed coating at the rate of 3 g kg-1 seed. The wetted seeds were air dried before sowing. Treated soybean seeds were sown in the first week of May of both seasons. A split plot design with three replicates was used in this experiment. The main plots were occupied by the salinity levels while sub-plots were occupied by the growth substances and fungicides. The area of each sub-plot was 3.5x3 m, sown with 375 seeds. The germination percentage and pre-emergence damping-off were determined after 20 days from sowing while post-emergence damping-off was determined at the 40th day from sowing.
Morphological and yield characters: Samples were taken at the 65th day from planting to estimate plant height (cm) and No. of branches and leaves plant-1. At harvesting, No. of pods plant-1, plant yield and weight of 100 seed were recorded.
Physiological characters: At the 65th day from sowing, the blade of the third leaf from tip (terminal leaflet) was taken to determine photosynthetic pigments (chlorophyll a, b and carotenoids) which extracted with methanol after adding traces of sodium carbonate (Robinson and Britz, 2000) and determined according to Mackinney (1941).
Total phenolics were determined after 65 days from sowing in fresh shoot using the folin-ciocalteau reagent according to Singleton and Roosi (1965).
Proline content was determined in shoot-dried material taken at the 65th day from sowing by the modified ninhydrine method (Troll and Lindsley, 1955) omitting phosphoric acid to avoid interference with concentrated sugars (Magne and Larher, 1992). Two grams of shoot dry weight were placed into tube containing 10 mL of distilled water. The tubes were kept in boiling water bath for 30 min, then cooled at room temperature. An aliquot from corresponding water extract was added to 2 mL ninhydrine reagent and the mixture was maintained in boiling water bath for 20 min, then cooled in an ice water bath. The product formed was extracted by 3 mL toluene by vigorous shaking. Absorption was measured spectrophotometically at 520 nm, using L-proline as standard.
Disease assessment: The disease severity of root-rot in mature stage was determined according to the scale suggested by Kravea (1960) as follows:
| 0 | = | Healthy plant |
| 1 | = | The lower part of stem is slight darkened |
| 2 | = | The region between the knots is heavily darkened |
| 3 | = | The lower part of the stem is heavily darkened while, the upper is whitened |
| 4 | = | Damping-off plant |
Stalk-rot disease severity was determined following the scale of Phillips (1971) in which:
| 0 | = | 0 cm (no discoloration under the first node) |
| 1 | = | 5 cm (discoloration through the first node) |
| 2 | = | 13 cm (discoloration through the second node) |
| 3 | = | 17 cm (discoloration through the third node) |
| 4 | = | 28 cm (discoloration through the fourth node) |
The severity of root rot and stalk rot diseases were calculated from the following formula:
Where:
| sd | = | Severity of disease. |
| a | = | No. of diseased plants having similar degree of infection |
| b | = | Degree of infection |
| Σ | = | Sum of (ab) |
| A | = | No. of examined plants |
| K | = | Highest degree of infection (in this case = 4) |
Sees quality: Soybean seeds were dried at 70°C for 48 h, grounded and analyzed for oil percentage (AOAC, 1970) and total nitrogen by semi-micro-Kjldahle (Pregl, 1945). The protein percentage was calculated by multiplying the N% by 6.25.
Statistical analysis: Data were analyzed with the statistical analysis software (CoStat, 2005). All multiple comparisons were first subjected to analysis of variance. Comparisons among means were made using Duncan's multiple range test at p<0.05 according to Gomez and Gomez (1984).
RESULTS
Isolation and pathogenicity test: Macrophomina phaseolina, Fusarium solani, F. oxysporum and Rhizoctonia solani; the causal pathogens of soybean root rot and stalk rot (brown stem rot) were isolated from infected plants. After identification trials, the four pathogens were further investigated for pathogenicity, the percentage of pre and post-emergence damping-off as well as healthy plants under greenhouse conditions are presented in Fig. 1. The tested fungi were pathogenic and caused pre- and post-emergence damping-off. This result was confirmed by the absence of emergence damping-off under non-infested soil (control). Rhizoctonia solani gave highest percentage of pre-emergence damping-off followed by M. phaseolina then F. solani. On the other hand, F. solani was recorded as the most aggressive pathogen in post-emergence damping-off followed by F. oxysporum then R. solani. The highest percentage of survival plants occurred under soil infested with F. oxysporum (64%) followed by F. solani (57.34%) then M. phaseloina (52%). In contrast, R. solani gave the highest percentage of diseased plants.
Anatomical structure of the infected soybean stem: The stem cross sections of healthy and infected plants by R. solani, M. phaseolina, F. solani and F. oxysporum showed obvious differences in anatomical structure (Fig. 2a-e). As depicted, the basal portion of the infected stem showed deformation in the anatomical structure occurred mainly in epidermis, cortex and pith compared with healthy plants.
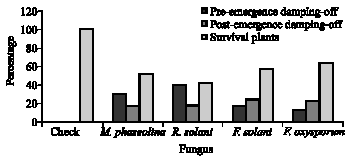 | |
| Fig. 1: | Pathogenicity test of pre- and post-emergence damping-off and survival soybean plants infected with isolated rot fungi under greenhouse conditions |
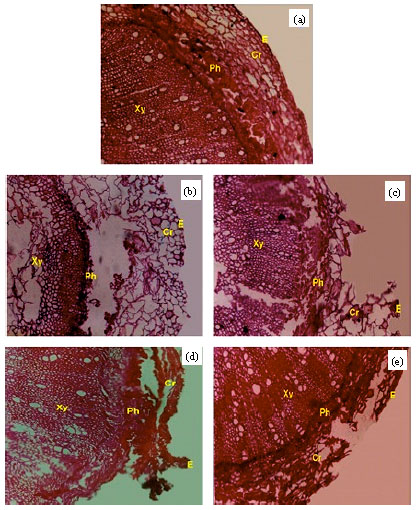 | |
| Fig. 2: | Cross sections of soybean stem basal portion in (a) healthy plant, (b) infected plant with R. solani, (c) infected plant with M. phaseolina, (d) infected plant with F. solani and (e) Infected plant with F. oxysporum. E: Epidermis; Cr: Cortex; Ph: Phloem; Xy: Xylem |
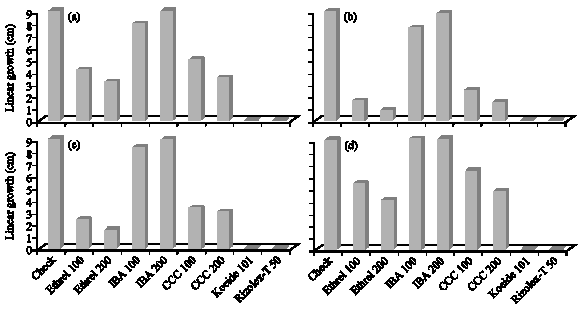 | |
| Fig. 3: | Effect of the tested growth substances and fungicides on the linear growth (cm) of the tested pathogenic fungi. (a) M. phaseolina, (b) R. Solani (c) F. solani and (d) F. oxysporum |
Complete disruption was observed in the epidermal cells and sever plasmolysis in the cortical cells with destruction of the outer cortical cells; R. solani was the most aggressive fungus. The disruption of cortical cells takes place in the deeper part followed by decay and degradation of primary cell wall. Some areas of cortex tissue showed a breakdown of cell wall components. The hyphae of fungal growth could be clearly seen in the inter- and intera-cellular epidermis, cortex and pith tissues. In severely infected stem by R. solani, the extensive damage in both cortex and pith tissues leading eventually to cell separation in some areas that leading to cell death. Generally, R. solani followed by M. phaseolina led to the injurious effects on basal stem structure more than F. solani and F. oxysporum.
Growth substances vis linear growth of the isolated pathogens: The effect of both levels of growth substances (Ethrel, IBA and CCC) and fungicides (Kocide 101 and Rizolex-T50) on the linear growth of the tested fungal pathogens are presented in Fig. 3a-d. Both fungicides completely inhibited the growth of all tested pathogenic fungi. On the other side, there is inversely relationship between increasing the concentration of growth substances and growth of the tested pathogenic fungi except IBA. The minimum growth of all pathogenic fungi occurred under the application of Ethrel (200 ppm) followed by CCC at 200 ppm then Ethrel 100 ppm as compared with the check. On contrast, IBA had no significant effect on fungal growth. There were differences in the sensitivity of the pathogens to the growth substances; R. solani being the most sensitive fungus, F. solani came next while, F. oxysporum came late.
Field evaluation of the tested growth substances
Germination percentage and damping-off: The effect of seed treatment with growth substances and fungicides on soybean germination and damping-off under salt stress conditions are presented in Table 1. Under natural infection, the germination of soybean seeds decreased significantly with increasing salinity level from 1000 to 3000 ppm during the two growing seasons. In contrast, pre- and post-emergence damping-off increased significantly with the salinity levels taking in consideration that post-emergence damping-off was less than pre-emergence damping-off.
Data also, show that all treatments at any dose used increased significantly germination percentage, at the same time decreased significantly damping-off. In this respect, rizolex-T was more effective followed by IBA at 200 ppm then kocide 101. Whereas, ethrel (200 ppm) did not cause any significant effect.
As for the interaction treatments data indicate that all growth substances counteracted the harmful effects of salinity on germination and both damping-off, IBA gave the highest effect followed by CCC, the high concentration (200 ppm) was more effective than the low concentration (100 ppm).
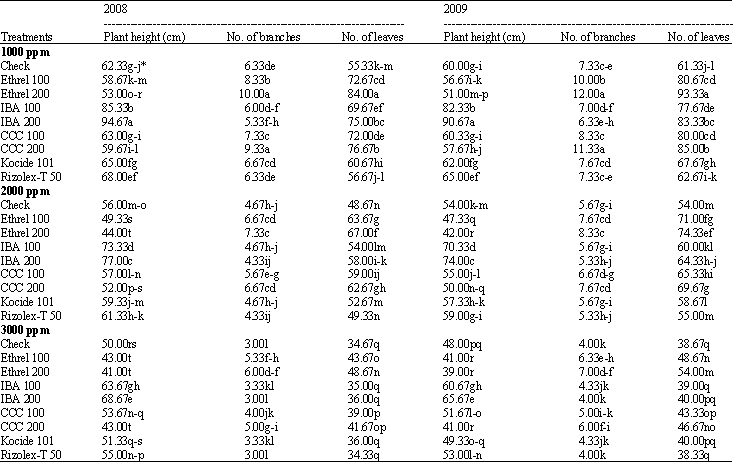
Morphological characters: The effect of growth substances on soybean plant height, No. of branches and No. of leaves under salt stress conditions during the two seasons of 2008 and 2009 are presented in Table 2.
| Table 1: | Effect of the tested growth substances and fungicides on germination percentage and damping-off disease of soybean plants grown under different salt stress conditions |
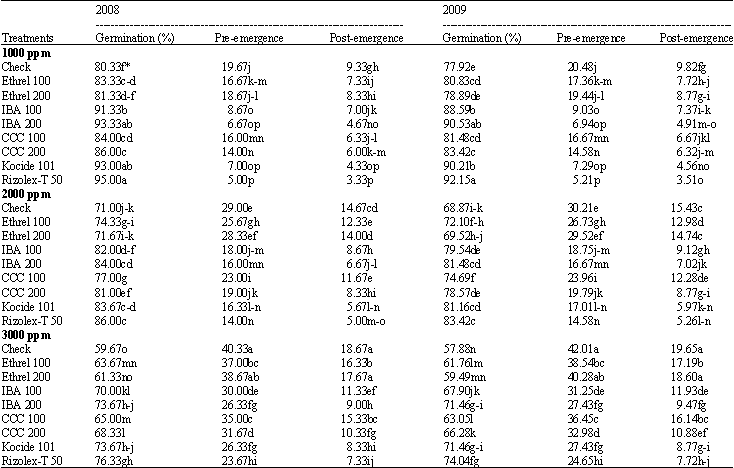 | |
| *Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range test at p<0.05 | |
It can be easily noticed the negative correlation between the salinity and the tested morphological characters. The greatest reduction in morphological characters occurred at 3000 ppm. Data also, indicate that both levels of IBA (100 and 200 ppm) at any level of salinity increased significantly plant height, IBA at 200 ppm was more effective. On contrast, an additive effects of salinity on decreasing plant height was recorded due to both levels of ethrel (100 and 200 ppm) and CCC (200 ppm) during the two seasons. While, CCC at 100 ppm and kocide 101 had no significant effect in this respect.
Any level of either ethrel or CCC increased significantly No. of branches plant-1 under salt stress. Ethrel (200 ppm) followed by (CCC 200 pmm) at 1000 ppm salt stress were the most effective. Meanwhile, other treatments had no significant effect on this parameter. All growth substances excreted positive significant effect on No. of leaves/plant at the different levels of salt stress.
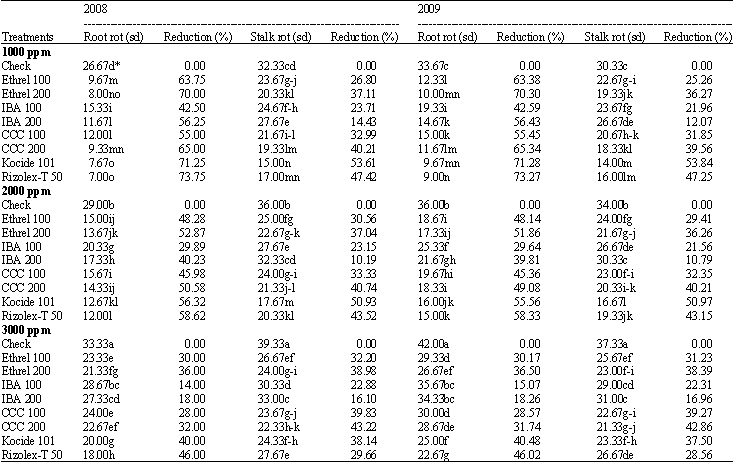
Disease incidence: Concerning the effects of salt stress on root rot and stalk rot disease, results of Table 3 show that the severity of root and stalk rots disease increased significantly with increasing salt stress from 1000 to 3000 ppm, the maximum increment occurred under the highest level (3000 ppm).
All tested materials clearly reduced the severity of root and stem rots compared to check during the two growing seasons. The maximum reduction in root and stalk rots were recorded with both fungicides. Ethrel (200 ppm) was the best growth substance followed by CCC (200 ppm) then ethrel (100 ppm). Taking in consideration that no significant differences between fungicides and high level of CCC or ethrel (200 ppm). The presence of growth substances at any level of salt stress decreased significantly the severity of root and stalk rots as compared with check. Ethrel (200 ppm) among growth substances gave the highest reduction in root-rot disease severity followed by CCC (200 ppm).
Physiological characters
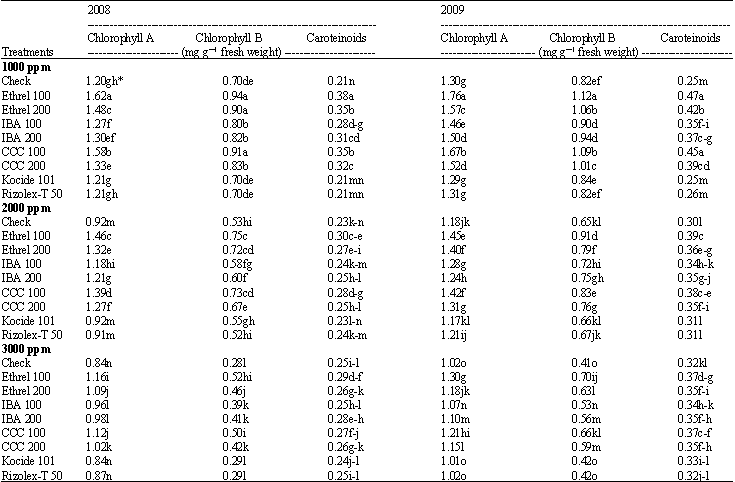
Photosynthetic pigments: Results in Table 4 show significant decrease in chlorophyll a and b with the increasing of salt stress level from 1000 to 3000 ppm, while, an induction response of salinity was recorded in carotenoids content. Concerning the effects of growth substances data show that all photosynthetic pigments (chlorophyll a, b and carotenoids) increased by using both levels of growth substances.
| Table 2: | Effect of some growth substances and fungicides on soybean morphological characters grown under different salt stress conditions |
 | |
| *Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range test at p<0.05 | |
| Table 3: | Effect of the tested growth substances and fungicides on disease incidence of soybean root rot and stalk rot under different salt stress conditions |
 | |
| *Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range test at p<0.05 | |
| Table 4: | Effect of the tested growth substances and fungicides on the photosynthetic pigments contents of soybean plant grown under different salt stress conditions |
 | |
| *Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range test at p<0.05 | |
The highly increase occurred under low level of ethrel followed by low level of CCC. It is worthy to mention that the low level of either ethrel or CCC was more effective than the high level. On the other hand, all photosynthetic pigments were not affected significantly under both of fungicides used.
The interaction between salinity level and growth substances significantly increased chlorophyll a and b as well as carotenoids content in both seasons. Generally, growth substances alleviated the injuries effects of salinity on chlorophyll content compared with fungicides.
Total phenols and proline content: Total phenols and proline content in soybean shoots increased significantly with increasing salinity levels from 1000 to 3000 ppm (Table 5), the high level of ethrel was more effective in this respect followed by the high level of CCC then IBA. However, both fungicides had not any significant effect on phenol and proline content in soybean shoots. Cycocel (200 ppm) followed by IBA (200 ppm) then ethrel (200 ppm) gave the highest values of total phenols and proline content under low salinity level, whereas, ethrel (200 ppm) followed by CCC (200 ppm) were the most effective under moderate and high salinity levels.
Yield and its components: The No. of pods plant-1, plant yield (g) and weight of 100 seeds decreased significantly with increasing salinity level (Table 6). On contrast, all concentrations of growth substances increased significantly these parameters compared with the check. The highest values of No. of pods plant-1 and plant yield occurred under application of ethrel (200 ppm) followed by CCC at 200 ppm while, no significant effect was recorded by using both fungicides. In both seasons, IBA at 200 ppm led to the maximum increase in weight of 100 seeds followed by ethrel at 200 ppm then IBA at 100 ppm. The application of growth substances counteracted the harmful effects of soil salinity on No. of pods plant-1, plant yield and weight of 100 seeds. Ethrel was the most effective treatment followed by CCC then IBA.
Seed quality: It is clear from Table 7 that oil and protein percentage in soybean seeds decreased significantly with increasing salinity level. The great reduction occurred under the high salinity level (3000 ppm). On the other hand, all concentrations of growth substances increased significantly soybean seed oil and protein percentage.
| Table 5: | Effect of the tested growth substances and fungicides on the total phenol and proline content of soybean plant grown under different salt stress conditions |
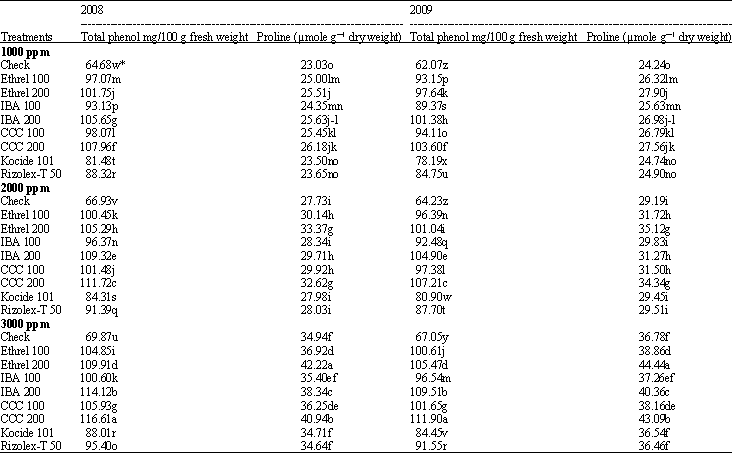 | |
| *Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range test at p<0.05 | |
| Table 6: | Effect of the tested growth substances and fungicides on yield components at harvest stage of soybean grown under different salt stress conditions |
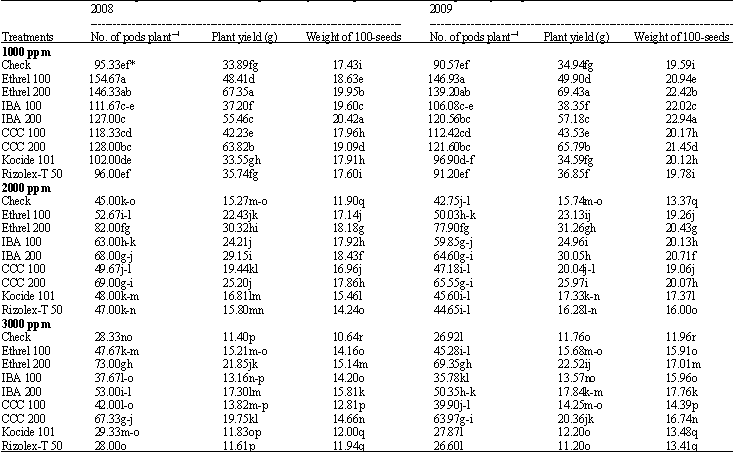 | |
| *Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range test at p<0.05 | |
| Table 7: | Effect of the tested growth substances and fungicides on the protein and oil content of soybean plant grown under different salt stress conditions |
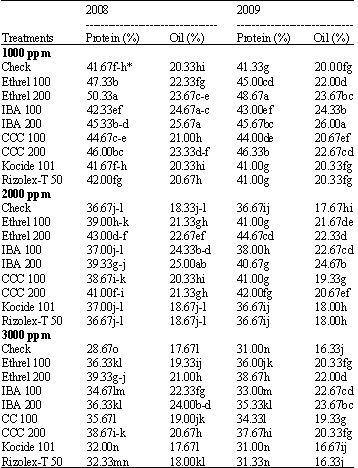 | |
| *Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range test at p<0.05 | |
The high level of ethrel followed by high CCC level recorded maximum protein content. Irrespective to the salinity level, the high seed oil content occurred under the application of IBA. On the other side, fungicides had no effect on seed oil and protein % in both seasons. Generally, the growth substances used counteracted the harmful effect of salinity on soybean seed oil and protein content.
DISCUSSION
The pathogenicity tests proved the pathogenic nature of the isolated fungi as causative agents of damping off. Rhizoctonia solani was the most aggressive as pre-emergence damping-off pathogen while, F. solani caused the highest rate of post-emergence damping-off. These results confirm the aggressive pathogenicity of the isolated fungi. The high rate of damping-off by such fungi may be occurred due to seed rot (Hussain et al., 1989) and inhibition of seed germination by fusaric acid, especially by Fusarium species (Matsui et al., 1988). Moreover, Gally et al. (1998) stated that Fusarium cause rotten lesions on seed cotyledons and plumule soft rot followed by pre-emergence damping-off. In addition, Oyekan and Naik (1987) found that M. phaseolina causes damping-off of seedlings. Also, the infection with these fungi leads to breakdown of the root system, in turn reducing the absorption surface and uptake of essential nutrients and water (Porter et al., 1990). In the present investigation, all tested pathogenic fungi cause heavy damage in seedlings anatomy structure (Fig. 2) which in turn causes post-emergence damping-off.
The harmful effects of salinity on germination, growth, yield and seed quality of soybean may be explained as; the salinity increased osmotic potential of the solution around the seeds and decreased water uptake by the seeds (Wang et al., 2001). Also, the depressing effect of chloride on enzymes activity such as hydrolytic enzymes e.g., protease and lipase (Younis et al., 1987; Munns, 2002) which is usually, followed by decrease in seed germination. Moreover, El-Nabarawy (1994) and El-Saht (1994) stated that the major effect of salinity on seed germination might be due to hormone imbalance in the seeds. The decrease in growth due to salinity may be attributed to the increase in respiration rate resulting from higher energy requirement (Schwarz and Gale, 1981), ionic imbalance (Abo-Hamid, 1994; Ashraf and Foolad, 2007; Sharifi et al., 2007) and decrease in meristematic activity and cell enlargement (Khadr et al., 1994). Kord and Khalil (1995) added that salinity reduced hormone delivery from root to leaves throughout the seedling, which inhibits plant growth. Van Hoorn et al. (2001) noted a decrease in rhizobium growth at increasing salinities and decrease of the No.of nodules. Moreover, salinity causes a disturbance of the nitrogen uptake (Jeuffroy and Sebillotte, 1997). In this study salinity decreased chlorophyll content in soybean leaves (Table 4) in turn, decrease carbohydrate content which followed by decrease oil and protein content in the seeds (Table 7). These results are in agreement with Smith et al. (1989) and Yang et al. (1990).
The possible mechanisms by which growth substances overcome the depressive effects of salinity are; enhancing water absorption by germinated seeds (Maske et al., 1997), cytokinins synthesis that affect plant-water balance (Mac Robbie, 1981), mineral composition (Abo-Hamid, 1994), physiological and hormonal levels within the plants (El-Saht, 1994), decreasing root resistance to water flow and Cl¯ uptake (El-Banna, 1985). Etherl decreased chloride uptake and this in turn may increase plant salt tolerance (Kasele et al.,1995). In this investigation, the effects of growth substances on alleviation the harmful effects of salinity may be due to the increase in total chlorophyll content (Table 4), phenol and proline (Table 5) which used as indicators in the osmoregulation of plant. The increase in branches number by etherl application may be due to the liberation of ethylene that considered as natural growth regulators which causes inhibition of terminal bud growth and stimulation of lateral shoot development (Helaly et al., 1984). Cycocel also increased No. of branches due to the imbalance in the endogenous hormone levels i.e., gibberellins, cytokinins and auxins, it also, lead to decrease in auxins level (Heather and Jones, 1983).
Indole butyric acid and CCC were the most effective in reducing soybean infections with damping-off. This is might be due to the inhibitory effect of growth substances on fungal growth (Fig. 3), sporulation and sclerotial formations (Khalifa, 2003). These results are in agreement with Marei (2000), Khalil (2002), Chowdhury (2003) and Metwally et al. (2006) on peanut root rot diseases. The results also indicate that etherl followed by CCC then IBA decreased significantly soybean infection with root rot and stalk rot disease. This may be due to the positive effect of these substances on chlorophyll, total phenol and proline content (Table 4 and 5). Phenolics are well known as antifungal, antibacterial and antiviral compounds occurring in plants. Moreover, proline is used as a storage compound for energy consequently, reduced carbon needs. Phenolics and proline are used as indicator for any stress conditions that occur to plants. The first step of the defense mechanism in plants involves a rapid accumulation of phenols at the infection site, which restricts or slows the growth of the pathogen (Matern and Kneusal, 1988; Gogoi et al., 2001). Moreover, Chowdhury (2003) stated that treated plants with growth regulators showed an increase in total phenol, calcium content and the activity of chatechol oxidase, which protect plants against pathogen stress. In this investigation, there was a positive relationship between chlorophyll and phenol content from one side and oil and protein percentage in soybean seeds from other side. This is in agreement with Smith et al. (1989), they reported that there is a relationship between photosynthesis and oil and protein content, hence induced sucrose translocated from leaf to the seeds, which metabolized to precursors for protein and oil. Therefore, any factor lead to increase in chlorophyll content will be expected to stimulate yield as well as oil and protein content in seeds.
There is no doubt that chlorophyll content is a good parameter reflecting the health condition of any plant. Photosynthesis or carbon assimilation consists in the synthesis of certain carbohydrates from CO2 and H2O by green cells in the presence of light. Carbohydrates are used as a source of energy. All living organisms require energy not only for growth and reproduction but also for maintenance of life itself. The light energy used is taken up by chlorophyll which present in chloroplast.
It could be concluded that the application of etherl, IBA and CCC at 200 ppm as seed soaking is recommended for reducing root and stalk rots in soybean plants as well as improving growth, yield and seed quality under salt stress conditions.
REFERENCES
- Abo-Hamid, S.A., 1994. Salinity-calcium interaction on growth analysis and ion content and proline levels at various growth stages of sorghum plants. J. Agric. Sci., Mansoura Univ., 19: 1701-1715.
Direct Link - Akande, S.R., O.F. Owolade and J.A. Ayanwole, 2007. Field evaluation of soybean varieties at Ilorin in the southern guinea savanna ecology of Nigeria. Afr. J. Agric. Res., 2: 356-359.
Direct Link - Aoki, A., A. Kanegami, M. Mihara, T. Kojima, M. Shiraiwa and H. Takahara, 2005. Molecular cloning and characterization of a novel soybean gene encoding a leucine-zipper-like protein induced to salt stress. Gene, 15: 135-145.
PubMed - Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Chowdhury, A.K., 2003. Control of Sclerotium blight of groundnut by plant growth substances. Crop Res., 25: 355-359.
Direct Link - Galley, A. and R.A. Fletcher, 1997. Relative efficiency of paclobutrazol, propiconazol and tetraconazole as a stress protectants in wheat seedlings. Plant Growth Regul., 21: 169-175.
CrossRefDirect Link - Gogoi, R., D.V. Singh and K.D. Srivastava, 2001. Phenols as a biochemical basis of resistance in wheat against karnal bunt. Plant Pathol., 50: 470-476.
CrossRefDirect Link - Jeuffroy, M.H. and M. Sebillate, 1997. The end of flowering in pea influence of plant nitrogen nutrition. Eur. J. Agron., 6: 15-24.
Direct Link - Kasele, I.N., J.F. Shanahan and D.C. Nielsen, 1995. Impact of growth retardants on corn leaf morphology and gas exchange traits. Crop Sci., 35: 190-199.
Direct Link - Macrobbie, E.A.C., 1981. Effects of ABA in 'isolated' guard cells of Commelina communis L. J. Exp. Bot., 32: 563-572.
CrossRefDirect Link - Mackinney, G., 1941. Absorbtion of light by cholorophyll solution. J. Biol. Chem., 140: 315-322.
Direct Link - Magne, C. and F. Larher, 1992. High sugar content of extracts interferes with colorimetric determination of amino acids and free proline. Anal. Biochem., 200: 115-118.
CrossRefPubMedDirect Link - Matern, U. and R.E. Kneusel, 1988. Phenolic compounds in plant disease resistance. Phytoparasitica, 16: 153-170.
CrossRefDirect Link - Matsui, Y., M. Murayama, S. Nishi and A. Ihnuma, 1988. Soybean blight caused by Fusarium oxysporum var redolens and production of fusaric acid by the fungus. J. College Dairying Jap. Nat. Sci., 12: 403-412.
Direct Link - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Muthukumarasam, M. and R. Panneerselvam, 1997. A melioration of Nacl stress by triadimefon in peanut seedlings. Plant Growth Regul., 22: 157-162.
CrossRefDirect Link - Nassiuma, D. and W. Wasike, 2002. Stability assessment of soybean varieties in Kenya. Afr. Crop Sci. J., 10: 139-144.
CrossRefDirect Link - Robinson, J.M. and S.J. Britz, 2000. Tolerance of a field grown soybean cultivar to elevated ozone level is concurrent with higher leaflet ascorbic acid level, higher ascorbate-dehydroascorbate redox status and long term photosynthetic productivity. Photosynth. Res., 64: 77-87.
CrossRefDirect Link - Schwarz, M. and J. Gale, 1981. Maintenance respiration and carbon balance. J. Exp. Bot., 32: 933-941.
Direct Link - Sharifi, M., M. Ghorbanli and H. Ebrahimzadeh, 2007. Improved growth of salinity-stressed soybean after inoculation with salt pre-treated mycorrhizal fungi. J. Plant Physiol., 164: 1144-1151.
CrossRefPubMedDirect Link - Singleton, V.L. and J.A. Rossi, 1965. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic., 16: 144-158.
CrossRefDirect Link - Smith, A.J., R.W. Rinne and R.D. Seif, 1989. Phosphoenolpyruvate carboxylase and pyruvate kinase involvement in protein and oil biosynthesis during soybean seed development. Crop Sci., 29: 349-353.
Direct Link - Troll, W. and J. Lindsley, 1955. A photometric method for the determination of proline. J. Biol. Chem., 215: 655-660.
Direct Link - Van Hoorn, J.W., N. Katerji, A. Hamdy and M. Mastrorilli, 2001. Effect of salinity on yield and nitrogen uptake of four grain legumes and on biological nitrogen contribution from the soil. Agric. Water Manage., 51: 87-98.
CrossRefDirect Link - Wang, D., M.C. Shannon and C.M. Grieve, 2001. Salinity reduces radiation absorption and use efficiency in soybean. Field Crop Res., 69: 267-277.
CrossRef - Yang, Y.W., R.J. Newton and F.R. Miller, 1990. Salinity tolerance in sorghum 1. whole plant response to sodium chloride in S. bicolor and S. halepense. Crop Sci., 30: 775-780.
Direct Link - Younis, M.E., M.N.A. Hasaneen and M.M. Nemet-Alla, 1987. Plant growth, metabolism and adaptation in relation to stress conditions IV. Effects of salinity on certain factors associated with the germination of three different seeds high in fats. Ann. Bot., 60: 337-344.
CrossRefDirect Link - Phillips, D.V., 1971. Influence of air temperature on brown stem rot of soybean. Phytopathology, 61: 1205-1208.
CrossRefDirect Link