
S.J. Goussous
Department of Plant Production, Faculty of Agriculture, Jordan University of Science and Technology, P.O. Box 3030, Irbid 22110, Jordan
K.M. Hameed
Department of Plant Production, Faculty of Agriculture, Jordan University of Science and Technology, P.O. Box 3030, Irbid 22110, Jordan
I. Saadoun
Department of Biotechnology and Biological Engineering, Jordan University of Science and Technology, P.O. Box 3030, Irbid 22110, Jordan
Plant Pathology Journal
Year: 2009 | Volume: 8 | Issue: 3 | Page No.: 98-105







ABSTRACT
Isolation of microorganisms antagonistic to Orobanche was attempted using Orobanche plants and rhizosphere soil of Orobanche-infected crops in Jordan. Six fungi (Cephalosporium sp., Cylindrocladium sp., Epicoccum sp., Fusarium sp., F. oxysporum and F. solani) were isolated. Of these, isolates that belonged to the Fusarium genus were the most common (more than 80% of isolates). Pathogenicity tests of these isolates on O. cernua stems indicated that Cylindrocladium sp., Fusarium sp., F. oxysporum and F. solani were most effective. These organisms caused total necrosis of inoculated stem tissues. Epicoccum sp. caused moderate damage (60% severity); while Cephalosporium sp. was least effective causing only localized necrosis. Only Cylindrocladium and Fusarium sp. isolates infected Orobanche inflorescence, causing maceration of these tissues as well as total destruction of seeds. Several bacterial isolates, including Pseudomonas and Actinomyces sp., were also recovered from soil sampled from different agricultural fields. These bacteria were found to be pathogenic in varying degrees to Orobanche stems and inflorescence. A previously isolated Streptomyces sp., R9, was also assessed in this study for its ability to inhibit O. cernua seed germination. Results showed a germination rate of 1-1.4% and 2.9-3.8% for R9 crude and lyophilized culture filtrates, respectively. In comparison, seed germination rates were 33.8-42.2% and 6.2-9.1% for water and culture medium controls, respectively. This study indicates that a wide range of microorganisms could be employed as antagonists to Orobanche. Their use as potential bioherbicides to control Orobanche appears promising.
PDF Abstract XML References Citation
Received: March 02, 2008;
Accepted: June 06, 2008;
Published: June 20, 2008
How to cite this article
S.J. Goussous, K.M. Hameed and I. Saadoun, 2009. Isolation and Evaluation of Indigenous Fungal and Bacterial Isolates as Potential Bioagents Against Broomrape (Orobanche cernua) in Jordan. Plant Pathology Journal, 8: 98-105.
DOI: 10.3923/ppj.2009.98.105
URL: https://scialert.net/abstract/?doi=ppj.2009.98.105
DOI: 10.3923/ppj.2009.98.105
URL: https://scialert.net/abstract/?doi=ppj.2009.98.105
INTRODUCTION
The genus Orobanche, known in its common name as broomrape, contains more than 100 species of obligate root holoparasites that attack many dicotyledonous plants including almost all vegetables and grain legumes (Kuijt, 1969; Musselman, 1980; Parker and Riches, 1993; Press and Graves, 1995). It infests millions of hectares of arable land in the Mediterranean region as well as in West Asia and Eastern Europe (Sauerborn, 1992; Abu-Irmaileh, 1994; Parker, 1994). At maturity, individual Orobanche plants can produce thousands of tiny seeds (250x300 μm in diameter), which can remain viable in the soil 13 years or longer (Kuijt, 1969; Press and Graves, 1995). Germination of these seeds occurs when stimulated by root exudates from hosts. The emerging parts, radicles, elongate and attach through haustoria to the vascular system in the host roots. At this stage, the parasite starts to withdraw water and assimilates from the host plant. A tubercle is formed that grows into a flowering stem, which eventually emerges above ground and produces flowers yielding thousands of seeds.
Orobanche sp. is extremely difficult to control (Amsellem et al., 2001a). This is mainly due to their efficient mechanisms of seed production, dispersal and longevity. Furthermore, broomrapes are in intimate physiological relationship with their host plants, which makes it difficult to apply conventional weed control measures. Various Orobanche control approaches have been attempted including cultural and mechanical methods, delayed planting of host crops, soil fumigation, soil solarization, trap and catch-crops, herbicides, herbicide-resistant genetically engineered crops, tolerant and resistant cultivars (Foy and Jacobsohn, 1989; Parker and Riches, 1993; Dhanapal et al., 1996). However, most of these control measures proved to be ineffective or costly. For these reasons, broomrapes are considered a good target for biological control (Amsellem et al., 2001a; Sauerborn et al., 2007). In the past two decades, several microorganisms that are potentially useful for Orobanche biocontrol have been isolated and their pathogenicity evaluated. Among the several microorganisms reported were isolates of Fusarium oxysporum var. orthoceras which gave some control of O. cernua (Bedi and Donchev, 1991) and O. cumana on sunflowers (Thomas et al., 1998, 1999a). Also, isolates of F. oxysporum and F. solani gave excellent results against O. ramosa without any damage to the tomato host (Boari and Vurro, 2004). In addition, two very promising isolates of F. arthrosporioides and F. oxysporum were isolated in Israel from juvenile O. aegyptiaca plants and were found to be pathogenic to O. aegyptica, O. crenata and O. ramosa on most vegetable crops (Amsellem et al., 2001b). In Jordan, Hameed et al. (2001) isolated several pathogenic fungi from several Orobanche species and found Fusarium sp. to be the most common. Recently, Nemat Alla et al. (2008) were able to proceed in their work beyond the isolation and evaluation procedures to the development of effective bioherbicide formulations, which consisted of microconidia and chlamydospores of two isolates of F. oxysporum encapsulated in a wheat flour-kaolin matrix.
Considering the seriousness of broomrape problem in Jordan and the Mediterranean area, this study was initiated to explore the biological control of Orobanche sp. by using indigenous antagonistic microorganisms. The main objectives were to isolate and identify different fungi and bacteria pathogenic to Orobanche sp. from the Jordanian habitats and to test their potential as biological control agents for O. cernua under laboratory conditions.
MATERIAL AND METHODS
Collection of Orobanche plants: Healthy as well as diseased stems of O. ramosa and O. cernua plants were collected from Orobanche-infested tomato, eggplant and faba bean fields located in the Northern parts of Jordan during the period of March to August, 2007 (Table 1).
Healthy samples were collected for the purpose of seed extraction. To do this, mature Orobanche capsules were harvested and air-dried at room temperature for several days. Afterward, capsules were gently broken to allow seed release. Collected seeds were passed through a steel sieve (300 μm) and stored at 4°C until use. Diseased Orobanche samples were collected for isolation of potential pathogens as described below. These samples consisted of Orobanche plants showing disease symptoms such as wilting, dry or soft rot, complete blight of the stem and/or black floral parts, but were growing in association with healthy host plants that exhibited no apparent signs/symptoms of any sort of infection (Fig. 1).
Isolation and identification of potential pathogens: Orobanche stems showing wilted or discolored parts were used for potential pathogen isolation. This was performed according to common isolation procedures as described (Johnston and Booth, 1983; Sinclair and Dhingra, 1995). Briefly, 1-2 cm pieces of wilted or discolored stems were surface-sterilized with 10% sodium hypochlorite solution plus 0.1% Tween for 5 min and rinsed several times with Sterilized Distilled Water (SDW). Next, sterile stem segments were placed on moistened filter paper inside closed sterile plastic boxes (5x35x15 cm) and incubated at 25°C for 2-3 days to enhance symptoms and signs of possible pathogens.
 | |
| Fig. 1: | A broomrape (Orobanche ramosa) plant attached to its cucumber host plant collected from a field in Northern Jordan during this study (2007). Note disease symptoms manifested as extensive rotting of Orobanche shoots |
| Table 1: | List of locations, dates of collection, and species of Orobanche samples collected during this study (2007) |
 | |
Isolates that grew out from the diseased tissues were isolated on Potato Dextrose Agar (PDA) in 90 mm Petri dishes and incubated at 25°C for 3-4 days. Afterwards, these isolates were purified by a single spore/colony isolation technique and maintained on PDA slants. The recovered fungal isolates were identified to the genus based on cultural and morphological characteristics using the identification keys and observations described by Peerally (1991) and Dugan (2006). A number of isolates belonging to the genus Fusarium was identified further to the species level using taxonomic descriptions by Nelson et al. (1983), Barnett and Hunter (1998) and Summerell et al. (2003).
Isolation of potential Orobanche pathogens from rhizosphere soil randomly sampled from fields containing Orobanche-infected plants was also attempted. One gram of each soil sample was processed using the soil serial dilution method (Johnston and Boot, 1983). Aliquots (0.1 mL) from these dilutions were plated on PDA and Nutrient Agar (NA) media and incubated at 25°C. Suspected bacterial isolates growing in these plates were re-isolated and purified by single colony transfer.
Pathogenicity on Orobanche plants in vitro: The pathogenicity of isolates was tested onto stems of O. cernua. Healthy and fresh stems were cut into segments (15-16 cm long) and then surface sterilized as described above. For inoculation, stem segments were gently pricked with a sterile needle. Then, a single plug (4 mm in diameter) cut from actively growing cultures was carefully placed up-side down on the cut stem. The inoculated site was wrapped with Parafilm strips to retain moisture. In the control treatment, segments were inoculated with sterile agar plugs. Inoculated stems were individually placed in sterilized plastic boxes with moist filter paper and incubated at 25°C for 7 days. Six replicates were used per fungal isolate and the experiment was repeated twice. Disease incidence was determined by the presence or absence of infection. Disease severity was defined as the percentage of disease (necrosis, rotting)-affected area of each stem segments (assessment grades from 1 to 4, Table 3) and the arithmetic means for single treatments were calculated.
The pathogenicity of isolates was also assessed onto O. cernua inflorescence. Freshly detached inflorescence from healthy plants were surface disinfected in 10% sodium hypochlorite for 5 min and rinsed several times with SDW. Fungal suspension of each isolate was prepared by flooding the culture plate with 25 mL SDW and then scraping the surface with a sterile spatula to dislodge the fungal growth. The number of fungal propagules (mycelial fragments and/or spores) in the resulting suspension was adjusted 109 mL-1 using a hemacytometer. Bacterial inocula were prepared from overnight single-colony cultures in Nutrient Broth (NB). Bacteria were collected by centrifugation and then reconstituted in SDW to 108 cfu mL-1 using a spectrophotometer. Sterile inflorescence were inoculated by dipping in these suspensions for 15 min. Sterile distilled water was used in the control treatment. After inoculation, inflorescence were placed inside sterile plastic boxes and incubated as described above. Six inflorescence were used per treatment and the experiment was done twice. Symptom severity was visually assessed as percentages using a 1-2 scale (Table 3).
At the end of trials, reisolation of fungi and bacteria was conducted to fulfill Koch’s Postulates for suspected pathogens. This was done from lesions on two inoculated O. cernua stems or inflorescence of each treatment. The resulting cultures were subcultured and identified.
Data for disease severity were subjected to analysis of variance, according to a complete randomized experimental design. Significant means were tested by the Fisher's Least Significant Difference (LSD) at 0.05 levels of probability.
Effect of R9 isolate on seed germination: Streptomyces isolate, R9, was shown to cause reduced seed germination of common broad-leaved and grass weeds (Saadoun et al., 2007). In this study, R9 was evaluated for its ability to inhibit O. cernua seed germination under laboratory conditions. This was done according to the agar germination bioassay described by Serghini et al. (2001) with slight modification. Fresh and healthy O. cernua seeds were surface-sterilized for 5 min in 10% sodium hypochlorite solution. Floating seeds were discarded and the remaining seeds were air-dried for 2 h under a laminar flow. For preconditioning, groups of 200-300 sterile seeds were sprinkled onto 9 mm moistened filter paper in Petri-dishes and incubated in a refrigerator (8°C) for 2 weeks. Subsequently, preconditioned seeds were uniformly spread onto water agar plates. To each plate, 3 mL of a germination stimulant solution was aseptically applied to induce germination. Two types of germination stimulants were used: (1) the synthetic stimulant, strigol, 'GR 24' (bis-lactone (XXIII)) dissolved in SDW at the rate of 0.2 μg L-1 (Johnson et al., 1976), and (2) root exudates obtained from lentil plants. The latter was prepared from 2-week-old lentil seedlings incubated with roots bathing in SDW at the rate of 25 seedlings per 500 mL water for 48 h in darkness. After addition of stimulants, seeds were left to settle for several hours. Next, seeds were treated with R9 culture filtrates in the crude form or as lyophilized powder reconstituted in SDW (Bataineh, 2004). To obtain R9 crude culture filtrate, the bacteria were grown in 250 mL Glucose Peptone Molasses (GPM) in shaken broth culture for 7 days at 28°C. The filtrate was collected by filtration and then cold sterilized by passage through 2 μM Millipore filters. In the prepared agar seed lawns, wells (10 mm in diameter) were cut in the center using a sterile cork borer. About 250 μL of R9 extracts either in the form of crude culture filtrates or as lyophilized powder reconstituted in SDW were transferred into the wells. Sterile distilled water and GPM broth were used as controls. Seed germination was evaluated after incubating the plates in a humid incubator at 25°C for 3 days. Using a binocular microscope, number of germinated seeds was counted from 100 seeds of each plate. Data were expressed as percentage of germination. Each treatment had four replicates and the experiment was repeated twice. Variances among experiments were homogenous, and the results from one experiment are presented.
RESULTS
Isolation and identification of potential pathogens: Thirty three fungal isolates were recovered from diseased Orobanche samples. Of these, 24 isolates were suspected to be pathogenic to Orobanche plants. Based on similarity in microscopic and macroscopic features, these selected isolates were divided into six groups. A representative isolate of each group was given a code and used for further identification and pathogenicity tests. The identity of the representative isolates were found to be: Cephalosporium sp., Cylindrocladium sp., Epicoccum sp., Fusarium sp., F. oxysporum and F. solani (Table 2). Of the selected isolates, Fusarium sp. (9 isolates) and F. oxysporum (8 isolates) were most frequently isolated. Three F. solani isolates were recovered, whereas, the rest of fungal genera were only isolated once or twice. The origin of all these fungi was O. cernua plants, except for one Fusarium sp. isolate, which was isolated from a diseased O. ramosa plant infecting faba bean.
Three bacterial isolates were recovered during this survey. Two of these isolates, BI4 and BI7, were recovered from diseased O. cernua plants, while the third, BIA, from the rhizosphere of Orobanche-infected plants (Table 2). These three isolates still require further testing to be identified. The rest of bacterial isolates (P9, P10, P14, P31, P35, P38, P40, P42, MR6, GM1X, Ma3 and R9) were obtained from native soils of different crop fields (Table 2).
| Table 2: | Identity and source of microorganisms isolated in this study (2007) |
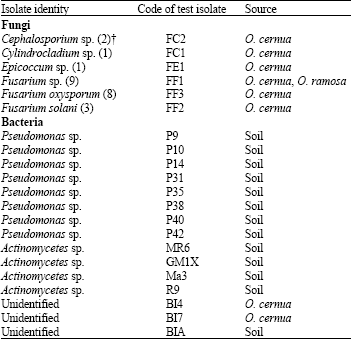 | |
| †: Numbers in parentheses indicate frequency of fungal isolates | |
These isolates were characterized in other studies to be Pseudomonas sp. (Benzina, 2004) and Actinomycetes sp. (Tahtamouni et al., 2006).
Pathogenicity on Orobanche plants: All tested fungal isolates developed symptoms on inoculated O. cernua stem segments, but to varying degrees (Table 3). Of these, test isolates of Cylindrocladium sp., Fusarium sp., F. oxysporum and F. solani were the most effective producing 80-92% severity ratings. Symptoms produced by these isolates consisted of marked necrosis, soft rot and loss of consistency of almost all stem tissue 7 days after inoculation (Fig. 2a). Isolate FE1 (Epicoccum sp.) produced moderate symptoms (60% severity) manifested by necrosis extending to about 2 cm beyond inoculation site, whereas FC1 (Cephalosporium sp.) was the least effective causing only localized damage (30% severity). Of the tested fungal isolates, only Cylindrocladium and Fusarium sp. affected O. cernua inflorescence (Table 3). These two isolates were highly effective in this respect causing complete maceration of inflorescence as well as the destruction of seeds.
Pseudomonas isolates, P10, P31 and P35 were highly pathogenic to O. cernua leading to almost total destruction of stem segments (90-100% damage) (Table 3, Fig. 2b). These three bacterial isolates were equally effective on O. cernua inflorescence causing their total necrosis (Fig. 2c). Isolates P9, P40 and P42, on the other hand, led to significantly lower severity ratings of about 50% on stems but with no apparent effect on inflorescence. Orobanche stems and inflorescence inoculated with the remaining two Pseudomonas isolates, P14 and P38, were undistinguishable from control (Table 3).
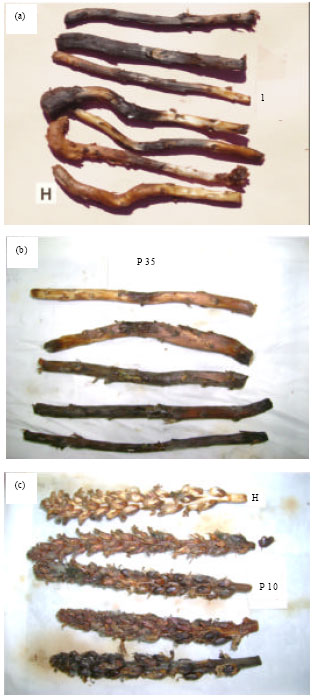 | |
| Fig. 2: | Pathogencity tests of microorganisms isolated in this study on Orobanche cernua stems and inflorescence. (a) and (b): Orobanche stem segments inoculated with Pseudomonas isolate P35 and the fungus Cylindrocladium sp., respectively; (c): Orobanche inflorescence inoculated with Pseudomonas isolate P10. Note the extent of disease reaction on the inoculated tissue compared to the healthy tissue (control, labeled H in (a) and (c) |
| Table 3: | Disease incidence and severity ratings of Orobanche cernua tissues (stems and inflorescence) inoculated with various fungi/bacteria isolated in this study (2007) |
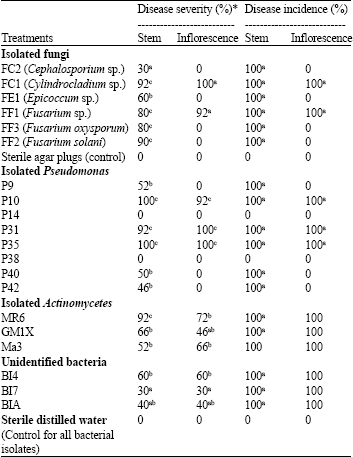 | |
| *Disease severity defined as the extent of necrosis (or rotting) of inoculated stem segments or inflorescence. Ratings on stem segments are based on a scale of 0-4. 0: No symptoms; 1: Symptoms localized to inoculation site (5% of stem tissue affected); 2: Infection surrounded stem at inoculation site (30% affected); 3: Tissue maceration extending 2-3 cm below and above inoculation site except for xylem tissue (60-70% of stem segment affected); 4: Loss of consistency of all stem tissue 7 days (100% of stem segment affected ). On inflorescence, assessment scale was as follows: 0: Inflorescence tissue unaffected, bearing healthy seeds; 1: Dark discoloration of surface tissues (50% of tissues affected); 2: Inflorescence are black with maceration of internal tissues, seeds destroyed (100% affected). -Values are means of six replicates; For each of the two group of organisms (fungi, bacteria), means within a column followed by the same letter are not significantly (p<0.05) different according to Fischer’s LSD test | |
All three Actinomycetes isolates had varying degrees of symptom severity on O. cernua stems and inflorescence (Table 3). Isolate MR6 produced the highest disease rating on stems (92%) but with only a moderate effect on inflorescence (72%). The other two Actinomycetes isolates, GM1X and Ma3, had moderate ratings (52-66%) on stem parts but were mild to moderate (46 and 66%, respectively) on inflorescence. The unidentified bacterial isolates, BI4, BI7 and BIA, caused mild to moderate damage ranging between 30 to 60% on both plant parts tested (Table 3). At the end of trials, all tested isolates were successfully re-isolated from the selected plant parts.
| Table 4: | Effect of culture filtrates (CF) of Streptomyces isolate, R9, on percent germination of Orobanche cernua seeds using two germination stimulants (GR-24 and lentil root exudates) |
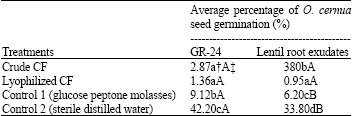 | |
| †Means within a column followed by the same small letter are not significantly (p<0.05) different according to Fischer’s LSD test. ‡Means within a row followed by the same capital letter are not significantly (p<0.05) different according to Fischer’s LSD test | |
Effect of R9 on seed germination: R9 isolate produced significant inhibition of O. cernua seed germination indicated by a germination rate of 0.95-1.36 and 2.87-3.80% for crude and lyophilized culture filtrates, respectively (Table 4). In comparison, seed germination rates for the water and GMP controls were significantly higher being 33.80-42.20 and 6.20-9.12%, respectively. As to the effect of stimulant type on seed germination, no significant differences existed between the two treatments (R9 crude and lyophilized filtrates) in this regard. In contrast, this effect was apparent between the SDW and GPM control treatments as germination rate was significantly higher (47 and 25%, respectively) when the synthetic stimulant GR-24 was used.
DISCUSSION
The agricultural habitats of Jordan have been heavily cultivated since biblical times. As a result, these habitats are expected to be very rich in microorganisms that interact with the parasitic plant Orobanche. Present study introduces several of such isolates from diseased Orobanche specimens and analyzes the rhizosphere soils of Orobanche-infected plants in Northern Jordan.
Results of this survey indicated that the most common (more than 80% of selected fungal samples) as well as the most effective of all isolates recovered from diseased Orobanche plants belonged to the genus Fusarium (Table 3). Such findings are not unexpected since almost all species that were previously isolated and proposed for biological control of Orobanche sp. belong to this genus (Thomas et al., 1999b; Amsellem et al., 2001a, b; Boari and Vurro, 2004; Sauerborn et al., 2007). In Jordan, Hameed et al. (2001) also reported the common occurrence of Fusarium sp. on all Orobanche species investigated. Soilborne pathogens have a number of practical advantages in the control of root parasites such as Orobanche (Sauerborn et al., 2007). For one, this parasite spends most of its life cycle below ground and thus produces most damage to host plants before emergence. Therefore, Fusarium provides a good candidate as an Orobanche pathogen since it can grow and survive through the soil for a long time.
Of the reported Fusarium sp. considered for biological control of broomrapes, F. oxysporum was the predominant species (Amsellem et al., 2001a, b; Boari and Vurro, 2004; Thomas, 1998, 1999a; Sauerborn et al., 2007; Nemat Alla et al., 2008). Few reports have characterized this fungus further to its special form orobanche (Saremi and Okhovvat, 2008). The specificity of this fungal form to infect only Orobanche sp. is an important attribute when considering a pathogen for biological control, since an Orobanche-specific pathogen may provide better environmental safety as a bioherbicide. At this stage and due to lack of modern technologies in our laboratory, identification of our selected F. oxysporum isolate, FF3, was not carried below the species level (Lievens et al., 2008). However, our survey in different locations showed this fungus to cause severe disease symptoms on O. cernua plants that were attached to healthy tomato or eggplant hosts. Furthermore, preliminary pathogenicity tests of FF3 on these two crops indicated negative results (data not shown). Based on these observations, we can conclude to a great extent of certainty that FF3 isolate belongs to the special form, orobanche. Nonetheless, studies are in progress to determine the exact identity of this isolate as well as other isolates that belong to the Fusarium genus.
Other fungi isolated in this study were Cephalosporium sp., Cylindrocladium sp. and Epicoccum sp. To our knowledge, this is the first report of natural association of Cephalosporium and Cylindrocladium with Orobanche plants. Epicoccum sp., on the other hand, was previously reported once in Nepal (Thomas et al., 1999b). Of these fungi, the soilborne Cylindrocladium sp. produced a total necrosis on both O. cernua stems and inflorescence to an extent comparable to Fusarium sp. isolates. Thus, this fungal isolate could also be useful for biocontrol of O. cernua providing that the host specificity of this fungus is addressed.
Three indigenous Pseudomonas isolates (P10, P31 and P35) and one Actinomycetes isolate (MR6) were highly aggressive on stem segments and inflorescence of O. cernua (Table 3). The three Pseudomonas isolates were earlier reported to promote growth in several crops (Benzina, 2004). Therefore, these isolates could represent an excellent option in Orobanche biocontrol. In addition, as these bacterial isolates (plus other previously mentioned fungal isolates) affect inflorescence, therefore, an alternative potential use of these isolates would probably be the prevention of O. cernua seed production. Postemergence spray application might be an option for delivery, provided that the right formulation can be found.
This study indicates the wide range of microorganisms that could be employed as antagonists of Orobanche and presents evidence supporting the importance of bacteria in Orobanche biological control research. Further, host-range tests of these isolates, including important crop plants, are needed to be carried out. Currently, studies are in progress to evaluate the ability of these organisms in the greenhouse and under field conditions.
ACKNOWLEDGMENTS
This research was supported by Grant No. 122-2007 from Abdul Hameed Shoman Fund for Supporting Scientific Research, Abdul Hameed Shoman Foundation, Amman, Jordan. The authors express their sincere appreciation to the Abdul Hameed Shoman Foundation for their generous financial support of this research through their Fund for Supporting Scientific Research. Continued gratitude is also expressed for the Deanship of Research at the Jordan University of Science and Technology for their legal and logistic coverage of the project.
REFERENCES
- Amsellem, Z., S. Barghouthi, B. Cohen, Y. Goldwasser and J. Gressel et al., 2001. Recent advances in the biocontrol of Orobanche (broomrape) species. BioControl, 46: 211-228.
CrossRefDirect Link - Boari, A. and M. Vurro, 2004. Evaluation of Fusarium spp. and other fungi as biological control of broomrape (Orobanche ramose). Biol. Control, 30: 212-219.
Direct Link - Hameed, K.M., I.M. Saadoun and Z. Al-Shyab, 2001. Potential biological control of Orobanche by fungi isolated from diseased specimens in Jordan. Plant Pathol. J., 17: 189-195.
Direct Link - Musselman, L.J., 1980. The biology of Striga, Orobanche and other root-parasitic weeds. Ann. Rev. Phytopathol., 18: 463-489.
CrossRef - Saadoun, I., R. Rawashdeh, T. Dayeh, Q. Ababneh and A. Mahasneh, 2007. Isolation, characterization and screening for fiber hydrolytic enzymes-producing streptomycetes of Jordanian forest soils. Biotechnology, 6: 120-128.
CrossRefDirect Link - Tahtamouni, M.E.W., K.M. Hameed and M. Saadoun, 2006. Biological control of Sclerotinia sclerotiorum using indigenous chitinolytic Actinomycetes in Jordan. Plant Pathol. J., 22: 107-114.
Direct Link - Amsellem, Z., Y. Kleifeld, Z. Kerenyi, L. Hornok, Y. Goldwasser and J. Gressel, 2001. Isolation, identification and activity of mycoherbicidal pathogens from juvenile broomrape plants. Biol. Control, 21: 274-284.
CrossRefDirect Link - Barnett, H.L. and B.B. Hunter, 1998. Illustrated Genera of Imperfect Fungi. 4th Edn., APS Press, Saint Paul, MN., ISBN-13: 978-0-89054-192-0, pp: 150-259.
Direct Link - Dhanapal, G.N., P.C. Struik, M. Udayakumer and P.C.J.M. Timmermans, 1996. Management of broomrape (Orobanche spp.): A review. J. Agron. Crop Sci., 176: 335-359.
Direct Link - Dugan, F.M. and F.M. Dugan, 2006. The Identification of Fungi: An Illustrated Introduction with Keys, Glossary and Guide to Literature. American Phytopathological Society, Saint Paul, MN, USA., ISBN: 9780890543368, Pages: 176.
Direct Link - Lievens, B., M. Rep and B.P.H.J. Thomma, 2008. Recent developments in the molecular discrimination of formae speciales of Fusarium oxysporum. Pest Manage. Sci., 64: 781-788.
CrossRefPubMedDirect Link - Nelson, P.E., T.A. Toussoun and W.F.O. Marasas, 1983. Fusarium Species: An Illustrated Manual for Identification. 1st Edn., Pennsylvania State University Press, University Park, University Park, PA., USA., ISBN-13: 978-0271003498, Pages: 226.
Direct Link - Nemat Alla, M.M., Y.M. Shabana, M.M. Serag, N.M. Hassan and M.M. El-Hawary, 2008. Granular formulation of Fusarium oxysporum for biological control of faba bean and tomato Orobanche. Pest Manage. Sci., 64: 1237-1249.
PubMedDirect Link - Peerally, A., 1991. The classification and phytopathology of Cylindrocladium species. Mycotaxon, 40: 323-366.
Direct Link - Saremi, H. and S.M. Okhovvat, 2008. Biological control of Orobanche aegyptiaca by Fusarium oxysporum f. sp. orobanche in northwest Iran. Commun. Agric Appl. Biol. Sci., 73: 931-938.
PubMedDirect Link - Sauerborn, J., D. Muller-Stover and J. Hershenhorn, 2007. The role of biological control in managing parasitic weeds. Crop Prot., 26: 246-254.
CrossRefDirect Link - Serghini, K., A.P. de Luque, M. Castejon-Munoz, L. Garcia-Torres and J.V. Jorrin, 2001. Sunflower (Helianthus annuus L.) response to broomrape (Orobanche cernua Loefl.) parasitism: Induced synthesis and excretion of 7-hydroxylated simple coumarins. J. Exp. Bot., 52: 2227-2234.
Direct Link - Summerell, B.A., B. Salleh and J.F. Leslie, 2003. A utilitarian approach to Fusarium identification. Am. Phytopathol. Soc. Plant Dis., 87: 117-128.
CrossRefDirect Link - Thomas, H., J. Sauerborn, D. Muller-Stover and J. Kroschel, 1999. Fungi of Orobanche aegyptiaca in Nepal with potential as biological control agents. Biocontrol Sci. Technol., 9: 379-381.
CrossRef