
M.A. Elwakil
Department of Plant Pathology, Faculty of Agriculture, Mansoura University, El-Mansoura, 35516, Egypt
I.M. El-Refai
Department of Botany, Faculty of Science, Tanta University, Tanta, Egypt
O.A. Awadallah
Department of Botany, Faculty of Science, Tanta University, Tanta, Egypt
M.A. El-Metwally
Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
M.S. Mohammed
Department of Botany, Faculty of Science, Tanta University, Tanta, Egypt
Plant Pathology Journal
Year: 2009 | Volume: 8 | Issue: 3 | Page No.: 90-97







ABSTRACT
This study was undertaken to study the seed-borne fungi of faba bean that attack the plants and reduce their yield in Egypt. The results provide a database for further study to control the pathogens. Twenty-six seed samples representing six faba bean cultivars collected from different parts of Egypt were used in this investigation. The blotter and deep-freezing methods were used. Surface- and non-surface-sterilized faba bean seeds were tested to detect and isolate the associated seed-borne fungi. The following 20 fungal species belonging to 13 genera were observed and identified: Aspergillus flavus (Link ex. Gray), Aspergillus niger (Van Tieghem), Aspergillus ochraceus (Wilhelm), Penicillium digitatum (Pers.:Fr.) Sacc., Penicillium italicum (Wehmer), Alternaria alternata (Fr.) Keissler, Botrytis faba (Sardina), Cephalosporium sp., Cladosporium cladosporioides (Frensen. de Vries), Epicoccum nigrum (Link), Fusarium oxysporum (Schlechlendahl), Fusarium semitectum (Berkeley and Ravenel), Fusarium solani (Mart.) Sacc., Fusarium verticillioides (moniliforme) (Sheld), Rhizoctonia solani (Kühn), Rhizopus stolonifer (Ehr. ex Fr.), Stemphylium globuliferum (Vestergr.) E.G. Simmons), Trichothecium roseum (Pers.) Link, Verticillium dahliae (Ehrenp) Vuill. The blotter method yielded a greater number of fungi than the deep-freezing method on both surface and non-surface sterilized seeds, but the deep-freezing method was better for slow-growing fungi. The pathogenicity test revealed that the most commonly isolated fungi from pre- and post-emergence damping-off and stunted seedlings were F. verticillioides, R. solani, Cephalosporium sp. and V. dahliae. These fungi significantly reduced the photosynthetic pigments in faba bean leaves. Fusarium verticillioides caused the greatest reduction in chlorophyll content (A, B and total chlorophyll). Fusarium oxysporum and V. dahliae significantly reduced carotenoid content. R. solani significantly reduced total phenols content when compared with the other tested fungi.
PDF Abstract XML References Citation
How to cite this article
M.A. Elwakil, I.M. El-Refai, O.A. Awadallah, M.A. El-Metwally and M.S. Mohammed, 2009. Seed-Borne Pathogens of Faba Bean in Egypt: Detection and Pathogencity. Plant Pathology Journal, 8: 90-97.
DOI: 10.3923/ppj.2009.90.97
URL: https://scialert.net/abstract/?doi=ppj.2009.90.97
DOI: 10.3923/ppj.2009.90.97
URL: https://scialert.net/abstract/?doi=ppj.2009.90.97
INTRODUCTION
Vicia faba, which has several common names (broad bean, fava bean, faba bean, horse bean, field bean, tic bean), is a species of bean (Fabaceae) native to North Africa and southwest Asia and is extensively cultivated elsewhere. Although, it is usually classified in the same genus Vicia as the vetches, some botanists place it in a separate monotypic genus as Faba sativa Moench. In much of the world, the name broad bean is used for the large-seeded cultivars grown for human food. Horse bean and field bean refer to cultivars with smaller, harder seeds (more like the wild species) used for animal feed, although their stronger flavour is preferred in some human food recipes, such as falafel. The term fava bean (from the Italian fava, meaning broad bean) is its most-used common name in the United States, with broad bean being the most-used common name in the UK.
In Egypt, faba beans are the most common fast food item in the Egyptian diet, eaten by rich and poor alike. Egyptians eat faba beans in various ways; the most popular way of preparing faba beans is taking cooked beans, mashing them and adding oil, lemon, salt and cumin. The prepared beans, called ful medames, are then eaten with bread.
Faba bean is an excellent source of protein (20-25%), calcium (0.15%), phosphorus (0.50%), lysine (1.5%) and methionine-cystine (0.5%), dry weight. It is also an excellent source of complex carbohydrates, dietary fiber, choline, lecithin, minerals and secondary metabolites (phenolics and levo-dihydroxy-phenylalanine (L-DOPA), which is the precursor of the neurotransmitter dopamine and naturally found in seedlings, green pods and beans) (Rabey et al., 1992).
In Egypt, faba bean yields slightly increased from 3192.80 kg ha-1 in 1997 to 3315.79 kg ha-1 in 2006. Its cultivated area gradually decreased from 149,160 ha in 1997 to 95,000 ha in 2006 (FAO, 2008).
Many faba bean pathogens were found to be seed-borne and seed transmitted. Therefore, several methods have been developed for testing seeds for associated microorganisms, which were discussed in various reviews (De Tempe, 1961, 1963, 1964; Agarwal, 1976; Neergard, 1977; Agarwal and Sinclair, 1987; Gaur and Dev, 1988).
All stages of faba bean growth were subjected to numerous injuries and stresses that interfere with their growth and development. There are many seed-borne fungi, while a number of fungi are serious pathogens on flowers and maturing seeds. These pathogens reduce the yield of seed both qualitatively and quantitatively. Other fungi, including saprophytes and very weak parasites, may lower the quality of seeds. The most common seed-borne fungi listed on faba bean are: Ascochyta fabae, which causes leaf and pod spot; Botrytis cinerea, the cause of grey mould; Botrytis fabae, the cause of chocolate spot; Fusarium sp., the cause of foot rot and wilts; and Rhizoctonia solani, the cause of damping-off of seedlings. Seed abortion, shrunken seeds, reduction in seed size, seed rot, seed necrosis, seed discoloration, reduction in germination capacity and physiological alterations in seed are the symptoms caused by these pathogens (Neergard, 1979).
Irwin (1987) declared that recent advances in the detection of seed-borne fungal pathogens have come largely through improvements in existing methods, in particular agar plating and incubation tests. These methods do not require sophisticated and expensive laboratory equipment and are thus widely applicable for use in developing countries.
Abdel-Hafez (1988) isolated Aspergillus, Penicillium, Rhizopus, Mucor and Fusarium from faba bean seeds. Of the 69 sp. and 4 vars. of pathogens isolated, he found that the most frequent species were A. niger, A. flavus, A. nidulans, A. terreus, A. flavus var. columnaris, M. chrysogenum, P. citrinum, P. funiculosum, R. stolonifer, M. hiemalis and F. moniliforme (Gibberella fujikuroi).
Dubey and Patel (2000) reported that Alternaria alternata, the cause of blight of broad bean (Vicia faba), remained viable in diseased plant debris or seeds for over one year. They showed that infected plant debris, infested soil or infected seeds were the primary pathogen sources for new infections.
Rauf (2000) isolated 24 fungi belonging to different genera from legume seeds in Pakistan using the blotter paper method. Alternaria alternata, Ascochyta sp., Colletotrichum sp., Fusarium sp. and Macrophomina phaseolina were the most frequent isolated fungi, all of which are known as common pathogens in these legumes.
This study was undertaken to analyze the seed-borne fungi of faba bean that attack the plants and reduce their yield in Egypt. The results provide a database for further study to control the pathogens.
MATERIALS AND METHODS
Collection of samples: Twenty-six seed samples of six susceptible faba bean cultivars (Giza 2, Sakha 1, 2, 3, Masr 1, Nubaria 1) were collected from the Agriculture station of Sakha and from commercial markets in different parts of Egypt, including Gharbia, Dakahlia, Kafr-Elshekh, Damitta and Sharkia Governorates, during the years 2004 and 2005.
Detection of seed borne fungi of faba bean by Seed Health Testing (SHT): Detection of seed-borne mycoflora of faba bean seeds was carried out according to the procedures published by the International Seed Testing Association (ISTA) (1996). Two hundred seeds of each faba bean sample were tested using the standard blotter and deep freezing methods.
Each sample was divided into two groups, non-surface sterilized seeds and seeds surface sterilized in 1% sodium hypochlorite solution for 5 min followed by 3 successive rinses in tap water. The excess water was removed by placing the treated seeds between sterilized tissue paper until dry.
Standard Blotter Method (SBM): Replicates of ten seeds were plated in 9 cm diameter Petri dishes containing three layers of blotters (filter paper) soaked with sterilized tap water. The plates were incubated at 25±2°C for 7 days under 12 h alternating cycles of cool white fluorescent light and darkness.
Deep Freezing Blotter Method (DFB): In this method, seeds plated as described above were incubated at 25±2°C for 24 h, then transferred to -20°C for 24 h. This was followed by 7 days incubation at 25±2°C for 12 h under alternating cycles of cool white fluorescent light and darkness.
Identification of the fungi isolated from seeds: Samples of faba bean seeds tested using the standard blotter method and deep freezing blotter method were examined under a stereoscopic binocular microscope (6-50X) for the presence of seed-borne fungi. A compound microscope was used to confirm the identifications by examining the morphology of conidia and conidiophores.
With the aid of the stereoscopic microscope, hyphal tips from the fungal colonies on seeds were picked up by using the tips of sterilized stretched capillary tubes and transferred to plates containing Water Agar (WA) medium (Dhingra and Sinclari, 1995). Pure cultures of the fungi were obtained and all isolates were maintained on Potato Carrot Agar (PCA) medium for further studies. The pure isolates were identified by comparison with the description sheets of the Commonwealth Mycological Institute, Kew, Surrey, England (CMI) and the Danish Government Institute of Seed Pathology (DGISP) (Raper and Fennel, 1965; Ellis, 1971; Chidambaram et al., 1973; Moubasher, 1977; Booth, 1985; Burrges et al., 1988; Singh et al., 1991).
PATHOGENICITY TESTING OF SEED-BORNE FUNGI OF FABA BEAN UNDER GREENHOUSE CONDITIONS
Six predominant fungal pathogens (Cephalosporium sp., Fusarium oxysporum, F. solani, F. verticillioides, Rhizoctonia solani and Verticillium dahliae) were selected from the seed health testing studies and evaluated for their pathogenicity on faba bean seeds and seedlings.
Inoculum from each of the above cultures was colonized separately on media consisting of crushed broad bean seed coats and peanut pod shells (1:1).
Five hundred milliliter glass bottles, each containing 200 g of this media moistened with water, were autoclaved and the media in each bottle inoculated with a 6 mm mycelial disk taken from a 7 day old colony grown on PDA. The glass bottles were then incubated at 25±2°C for 14 days.
Sandy clay soil 1:2 (w/w) was chemically sterilized using a 5% formaldehyde solution and transferred into 15 cm diameter pots, each containing 3 kg soil. Pots were inoculated with the selected fungi growing in the seed coat/pod media separately at the rate of 5% (w/w) and kept in the greenhouse for one week before sowing the faba bean seeds. Pots containing non-inoculated soil were used as controls. Three replicates were used per treatment.
Pathogen-free seeds raised from the previously tested lot of cultivar Giza 2 were surface sterilized and planted (5 seeds/pot) in both inoculated and non-inoculated soil. All pots were maintained in the greenhouse under natural conditions during the winter season and watered as needed. Fifteen days after sowing, the disease ratios were determined by recording the number of non-emerged seeds (pre-emergence damping-off), while post-emergence damping off and surviving plants were recorded 30 days after sowing. The equations described by Khalifa (1987) were followed:
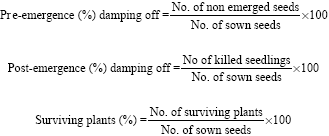 |
Re-isolations of the tested fungi were made from diseased seeds and seedlings manifesting symptoms.
Estimation of photosynthetic pigments content: The spectrophotometric method recommended by Metzner et al. (1965) was used in this investigation. A definite weight of fresh leaves was homogenized immediately after harvesting in 5 mL cold aqueous acetone (85%), kept overnight in a refrigerator and then centrifuged. The supernatant, which contained pigments, was diluted with cold aqueous acetone to an appropriate volume for spectrophotometric measurements. The extract was measured against a blank of pure 85% acetone solution at three wavelengths: 452, 644 and 663 nm. The concentrations of chlorophyll A (Chl. A), chlorophyll B (Chl. B) and carotenoids (Carot.) were determined using the following equations:
Chl. a = 10.3 E663-0.918 E644 Chl. b = 19.7 E644-3.87 E663 Carot. = 4.2 E452-(0.0264 Chl. a + 0.426 Chl. b) |
Estimation of total phenols content: Total phenols were determined using the Folin-Ciocalteau reagent (Singleton and Rossi, 1965). For preparation of the calibration curve, 1 mL aliquots of 0.02, 0.04, 0.06, 0.08 and 1.00 mg mL-1 ethanolic catechol solutions were combined with 0.5 mL Folin-Ciocalteau reagent. After 3 min, 2 mL (200 g L-1) sodium carbonate was added and the contents were mixed thoroughly. The final color change was measured colorimetrically at 650 nm using a SPEKOL11 (Carl Zeiss, Jena, Germany) and the calibration curve was drawn.
Samples (2 g) of fresh leaves were homogenized in 80% aqueous ethanol at room temperature and centrifuged in a cold centrifuge at 10,000 rpm for 15 min and the supernatant was saved. The residues were re-extracted twice with 80% ethanol and the supernatants were pooled, put in evaporating dishes and evaporated to dryness at room temperature. The residues in the evaporating dishes were dissolved in 5 mL of distilled water. One hundred microlitres of this extract was diluted to 3 mL with water and mixed with the same amount of reagent as described above. The color was developed and absorbance measured after 60 min. The results were expressed as mg catechol/100 g fresh weight material. All determinations were replicated three times. Total content of phenolic compounds in plant ethanolic extracts was calculated as catechol equivalents by the following equation:
where, T is Total content of phenolic compounds, in mg of catechol/100 g of fresh weight material, c is the concentration of catechol established from the calibration curve, in mg mL-1, V is the volume of extract in mL and m is the weight of pure plant ethanolic extract in g.
Statistical analysis: Data were subjected to analysis of variance using SAS version 6.11 by Duncan’s multiple range tests (Duncan, 1995).
RESULTS
Fungi recovered from faba bean seeds
Seed Health Testing (SHT): Blotter and deep-freezing methods recommended by ISTA (1996) were used. Non-surface-sterilized seeds and seeds surface-sterilized in 5% sodium hypochlorite solution were examined to detect seed borne fungi of faba bean. Twenty-six samples of seeds were selected for use in the present investigation: 200 seeds from each sample were tested.
Twenty fungal species belonging to 13 genera (Table 1) were isolated and identified from the collected seed samples using seed health testing methods.
Table 1 shows the three species of the genus Aspergillus found on the seeds, as described by Raper and Fennell (1965). These species were Aspergillus flavus, A. niger and A. ochraceous.
The isolates of Penicillium found were P. digitatum and P. italicum, as described by Raper and Tom (1949).
The table also shows 15 fungal species other than the above fungi. They were identified as belonging to 11 different genera. The genus Fusarium included five species: F. oxysporum, F. semitectum, F. solani and F. verticillioides. The other genera isolated included only one species each.
| Table 1: | List of the identified fungi isolated from faba bean seeds by seed health testing methods by ISTA, 1996 |
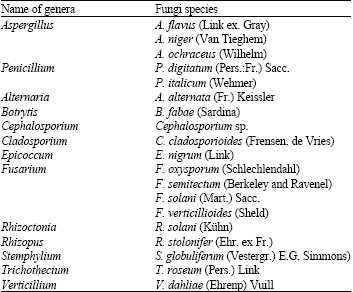 | |
Frequency of isolation of the seed-borne fungi of faba bean: The frequency of isolation of each fungus was recorded in terms of percentage and range of infection. The tables show that the blotter method yielded a greater number of fungi than the deep-freezing one, with both surface-sterilized and non-surface-sterilized faba bean seeds.
Fungi isolated from non-surface-sterilized seeds
Standard Blotter Method (SBM): Table 2 shows that the standard moist blotter method (SBM) enhanced the growth of saprophytic fungi.
The isolated fungal genera are ranked as follows according to the total mean percentage: Penicillium (37.09%), Aspergillus (18.42%), Cladosporium (9.29%), Alternaria (2.18%), Rhizopus, (2.06%), Rhizoctonia (0.98%), Fusarium (0.56%), Botrytis (0.33%), Stemphylium (0.29%), Trichothecium (0.15%), Epicoccum (0.12%) and Cephalosporium (0.08%).
According to the occurrence percentage, the isolated genera ranked as follows: Aspergillus, Penicillium, Cladosporium, Alternaria, Rhizopus, Fusarium, Stemphylium, Cephalosporium, Trichothecium, Botrytis, Epicoccum and Rhizoctonia.
The standard moist blotter method (SBM) with non-surface-sterilized seeds showed an increase in the growth of fast growing saprophytes, viz., A. flavus, A. niger, Rhizopus stolonifer and Penicillium digitatum and a decrease in the growth of the slow growing seed-borne fungi, viz., F. solani, F. oxysporum, Cephalosporium sp. and Verticillium sp. This method was suitable for detecting Rhizoctonia solani.
| Table 2: | Incidence of seed-borne fungi in 26 non- surface sterilized seeds samples of faba bean |
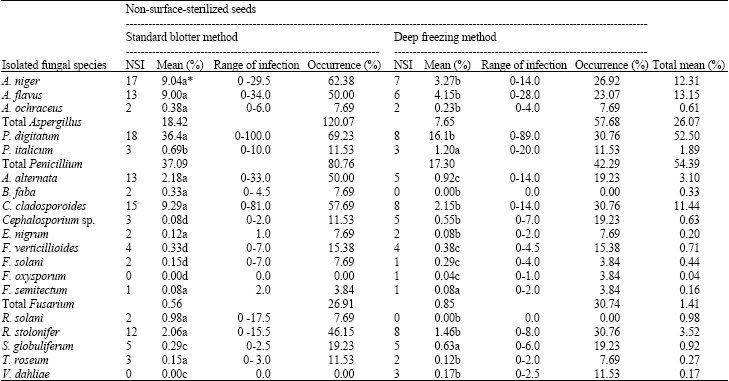 | |
| NSI: No. of seed lots infected, Mean (%) = Mean infection of the tested samplesxtotal No. of samples examined = 26, Occurrence (%) = NSI/total No. of samples 100*, Means within a column followed by the same letter(s) are not significantly different according to Duncan’s multiple range tests | |
Deep Freezing Method (DFM): Table 2 shows the fungi isolated from faba bean seeds using the deep freezing blotter method (DFM) and their percentages: Penicillium (17.30%), Aspergillus (7.65%), Cladosporium (2.15%), Rhizoctonia (1.46%), Rhizopus (1.46%), Alternaria (0.92%), Fusarium (0.85%), Stemphylium (0.63%), Cephalosporium (0.55%), Verticillium (0.17%), Trichothecium (0.12%) and Epicoccum (0.08%).
According to the occurrence percentage, the isolated genera were ranked as follows: Aspergillus, Penicillium, Cladosporium, Rhizopus, Fusarium, Alternaria, Cephalosporium, Stemphylium, Verticillium, Epicoccum and Trichothecium.
The deep freezing blotter method (DFM) with non-surface-sterilized seeds showed a decrease in the growth of fast growing saprophytes, including A. flavus, A. niger, Rhizopus stolonifer and Penicillium digitatum and an increase in the slow growing seed-borne fungi, viz., F. solani, F. oxysporum, Cephalosporium sp. and Verticillium sp. This method was not suitable for detecting Rhizoctonia solani.
Fungi isolated from surface sterilized seeds
Standard Blotter Method (SBM): Table 3 shows that the standard moist blotter method (SBM) inhibited fast-growing saprophytes more with surface-sterilized seeds than with non-surface-sterilized seeds.
Based on total mean percentage, the isolated fungal genera ranked as follows: Fusarium (4.59%), Aspergillus (2.79%), Penicillium (2.61%), Cladosporium (0.88%), Alternaria (0.30%), Rhizopus (0.15%), Stemphylium (0.15%) and Cephalosporium (0.14).
Based on percentage occurrence, the isolated genera ranked as follows: Fusarium, Aspergillus, Penicillium, Cladosporium, Alternaria, Rhizopus, Stemphylium and Cephalosporium.
The standard moist blotter method (SBM) with surface-sterilized seeds showed an increase in the growth of fast-growing saprophytes, viz.: A. flavus, A. niger and Penicillium digitatum, but less than with non-surface-sterilized seeds. It showed a decrease in the growth of the slow-growing seed-borne fungi, viz; F. solani, F. oxysporum, Cephalosporium sp., F. verticillioides and Verticillium sp., but higher than with non-surface-sterilized seeds.
Deep Freezing Method (DFM): Table 3 shows that the deep freezing blotter method (DFM), when used with surface-sterilized seeds, inhibits the fast-growing saprophytic fungi and increases the growth of slow-growing seed-borne fungi more than when used with non-surface-sterilized seeds.
Based on total mean percentage, the isolated fungal genera ranked as follows: Fusarium (8.24%), Verticillium (2.31%), Penicillium (2.00%), Aspergillus (1.68%), Alternaria (1.38%), Cephalosporium (0.77%), Stemphylium (0.46%), Cladosporium (0.23%), Rhizopus (0.15%) and Trichothecium (0.08%).
| Table 3: | Incidence of seed-borne fungi in 26 surface-sterilized seeds samples of faba bean |
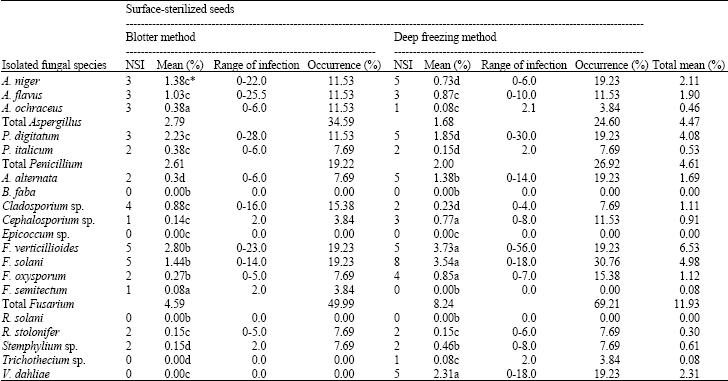 | |
| NSI: No. of seed lots infected, Mean (%) = Mean infection of the tested samplesxtotal No. of sample examined = 26, Occurrence (%) = NSI/total No. of samplesx100*, Means within a column followed by the same letter(s) are not significantly different according to Duncan’s multiple range tests | |
Based on percentage occurrence, the isolated genera ranked as follows: Fusarium, Penicillium, Aspergillus, Alternaria, Verticillium, Cephalosporium, Cladosporium, Rhizopus, Stemphylium and Trichothecium.
The deep freezing blotter method (DFM), when used with surface-sterilized seeds, showed a significant decrease in the growth of fast-growing saprophytes, including A. flavus, A. niger, Rhizopus stolonifer and Penicillium digitatum and a significant increase in the slow-growing seed-borne fungi, viz.: F. solani, F. oxysporum, F. verticillioides, Cephalosporium sp. and Verticillium sp.
PATHOGENICITY TESTING OF SOME PATHOGENIC SEED-BORNE FUNGI OF FABA BEAN
Results of the pathogenicity tests of Cephalosporium sp., F. solani, F. oxysporum, F. verticillioides, R. solani and V. dahliae are shown in Table 4. Disease severity readings of the pathogens on seeds and seedlings were made 15 and 30 days after planting for pre- emergence damping off, post-emergence damping off and stunted seedlings.
Data in Table 4 shows that F. verticillioides had the highest percentage of pre-emergence damping off among all isolated fungi. The percentage of infection was 53.33% on seed compared with the control treatment, which had 0% infection.
| Table 4: | Pathogenicity testing of some seed-borne pathogenic fungi of faba bean under greenhouse conditions |
 | |
| Control = non-infested soil, *Means within a column followed by the same letter(s) are not significantly different according to Duncan’s multiple range tests | |
R. solani caused the highest percentage of post-emergence damping off among all the pathogens. The recorded percentage was 33.33% on seedlings.
Cephalosporium sp. and V. dahliae caused the highest percentage of stunted seedlings among all isolated fungi. The recorded percentage was 33.33% on seedlings, while the controls had 6.67% stunted seedlings.
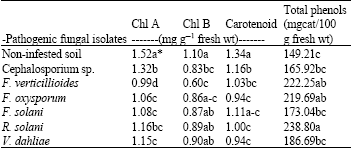
Effect of the isolated pathogenic fungi on the concentration of photosynthetic pigments and total phenols content in faba bean seeds grown in pathogen-infested soil under greenhouse conditions: Data in Table 5 shows that chlorophyll A content of infected plants was significantly affected by the stress of F. verticillioides, which was the most aggressive isolated fungus.
| Table 5: | Effect of pathogenic fungal isolates on photosynthetic pigments and total phenols content of faba bean plants under greenhouse conditions |
 | |
| Control: Non-infested soil, *Means within a column followed by the same letter(s) are not significantly different according to Duncan’s multiple range tests | |
It lowered the content of chlorophyll A to 1.06 mg g-1 fresh wt., while the control treatment had 1.52 mg g-1 fresh wt.
Chlorophyll B content of infected plants was also affected significantly by F. verticillioides, decreasing to 0.60 mg g-1 fwt while the control had 1.10 mg g-1 fwt.
Both F. oxysporum and V. dahliae reduced the carotenoid content of infected plants. They lowered it to 0.94 mg g-1 fresh wt. compared with the control, which had 1.19 mg g-1 fresh wt.
Table 5 also shows that the total phenols content of infected plants was significantly increased due to the effects of the tested pathogens. R. solani caused the greatest increase. It resulted in 238.80 mg catechol/100 g fwt compared with the control treatment, which had 149.21 mg catechol/100 g fresh wt.
DISCUSSION
Faba bean in Egypt is susceptible to a number of fungal diseases which decrease production and lower the quality of seeds. Due to the lack of information regarding seed-borne diseases of this crop in Egypt, the researchers studied the seed-borne fungi of faba bean which affect production and seed quality. The blotter seed health as well as deep freezing methods were used (ISTA, 1996).
Use of the standard blotter method results in rapid saprophyte growth, which often impairs the detection of parasitic fungi. Use of the deep-freezing method results in the growth of saprophytic bacteria and yeasts, which inhibit spore germination of some important seed-borne fungi (Neergard, 1979).
Isolation trials revealed Aspergillus flavus, Aspergillus niger, Aspergillus ochraceus, Penicillium digitatum, Penicillium italicum, Alternaria alternata, Botrytis faba, Cephalosporium sp., Cladosporium cladosporioides, Epicoccum nigrum, Fusarium oxysporum, Fusarium semitectum, Fusarium solani, Fusarium verticillioides (moniliforme), Rhizoctonia solani, Rhizopus stolonifer, Stemphylium globuliferum, Trichothecium roseum and Verticillium dahliae. These results are in agreement with the findings of other researchers (Abdel-Hafez, 1988; Dubey and Patel, 2000; Rauf, 2000).
Pathogenicity tests showed that Cephalosporium sp., F. solani, F. oxysporum, F. verticillioides, R. solani and V. dahliae were the most pathogenic fungi isolated from faba beans which showed seed decay, pre-emergence damping off and post-emergence damping-off. The concentrations of chlorophyll A, chlorophyll B and carotenoid in plants infected with the above-mentioned pathogens showed significant reductions, while the total phenols in diseased plants increased.
These results indicate that there is a correlation between the disease incidences due to the above fungi and the concentration of total phenols, as well as chlorophyll A, chlorophyll B and carotenoid, in plant tissue. The use of pathogen-free seed is a must to produce quality faba bean seeds.
ACKNOWLEDGMENTS
The authors thank Dr. Conrad J. Krass, Primary State Plant Pathologist, California, Department of Food and Agriculture, Sacramento, CA, USA (retired) for critical review of the manuscript.
REFERENCES
- Metzner, H., H. Rau and H. Senger, 1965. Untersuchungen zur synchronisierbarkeit einzelner pigmentmangel-mutanten von Chlorella. Planta, 65: 186-194.
CrossRefDirect Link - Neergard, P., 1977. Seed Pathology: Volumes 1 and 2. Red Globe Press London, New Wharf Road, London, Pages: 1164.
CrossRefDirect Link - Rabey, J.M., H. Shabtai, E. Graff and A.D. Korczyn, 1992. Improvement of Parkinsonian features correlate with high plasma levodopa values after broad bean (Vicia faba) consumption. J. Neurol Neurosurg Psychiat, 55: 725-727.
PubMedDirect Link - Singleton, V.L. and J.A. Rossi, 1965. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic., 16: 144-158.
CrossRefDirect Link