
H.R. Kavousi
Department of Biotechnology and Plant Breeding, College of Agriculture, Ferdowsi University of Mashhad, Mashhad, Iran
H. Marashi
Department of Biotechnology and Plant Breeding, College of Agriculture, Ferdowsi University of Mashhad, Mashhad, Iran
J. Mozafari
National Plant Gene-Bank, Seed and Plant Improvement Institute, Mahdasht Road, Karaj, Iran
A.R. Bagheri
Department of Biotechnology and Plant Breeding, College of Agriculture, Ferdowsi University of Mashhad, Mashhad, Iran
Plant Pathology Journal
Year: 2009 | Volume: 8 | Issue: 3 | Page No.: 127-132







ABSTRACT
The fungal disease, ascochyta blight, caused by Ascochyta rabiei is a major yield limiting factor of chickpea (Cicer arietinum L.) around the world. Expression analysis of genes induced in general defense response can provide clues to elucidate major defense mechanisms against pathogen infection in chickpea plants. The role of key phenylpropanoid pathway enzymes response to Ascochyta rabiei in chickpea was studied under greenhouse conditions using a reverse transcription and semi-quantitative polymerase chain reaction (SQ-PCR). Transcript accumulation of four genes encoding phenylalanine ammonia-lyase (PAL), chalcon synthase (CHS), isoflavone reductase (IFR) and Flavanone 3-Hydroxylase (F3H) induced in response to race 3 of A. rabiei was compared in resistant and susceptible genotypes. Results obtained in this study showed that in resistant genotype all 4 phenylpropanoid pathway genes: PAL, CHS, IFR and F3H were rapidly up regulated 6 h after inoculation with race 3 of A. rabiei. However, transcripts of PAL and IFR genes were rapidly accumulated in both resistant and susceptible cultivars. Therefore, induction of key enzymes of phenylpropanoid pathway appeared to be an important defense mechanism of chickpea plants against A. rabiei.
PDF Abstract XML References Citation
How to cite this article
H.R. Kavousi, H. Marashi, J. Mozafari and A.R. Bagheri, 2009. Expression of Phenylpropanoid Pathway Genes in Chickpea Defense Against Race 3 of Ascochyta rabiei. Plant Pathology Journal, 8: 127-132.
DOI: 10.3923/ppj.2009.127.132
URL: https://scialert.net/abstract/?doi=ppj.2009.127.132
DOI: 10.3923/ppj.2009.127.132
URL: https://scialert.net/abstract/?doi=ppj.2009.127.132
INTRODUCTION
Chickpea (Cicer arietinum L.) is the third most important pulse crop in the world, but a major factor limiting its world production is a destructive fungal disease known as ascochyta blight caused by Ascochyta rabiei (Pass.) L. (Coram and Pang, 2007). Breeding high yielding cultivars with enhanced resistance is considered the most effective means of controlling the disease. Intensive genetic analyses successfully identified major loci determining fungal resistance in chickpea. Similar studies also revealed the multigenic nature of the resistance as well as Quantitative Trait Loci (QTL) associated with resistance to ascochyta blight in chickpea (Cho and Muehlbauer, 2004). In addition to this information revealing the mode of action of the resistance genes, understanding the molecular mechanism of the resistance against A. rabiei is equally important.
Plants respond to pathogen infection by activating certain responses that have been implicated as mechanisms of disease resistance. These responses include the hypersensitive reaction, the de novo synthesis of phytoalexins, proteinase inhibitors, hydrolytic enzymes and increased production and deposition of lignin into the plant cell wall (Lawton and Lamb, 1987). A ubiquitous feature of plant response to pathogen challenge is apparently the enhanced activation of phenylpropanoid pathway, suggesting general defensive role for phenylpropanoid compounds (Moniz de Sa et al., 1992). Phenylpropanoid metabolism comprises a complex series of branching biochemical reactions which lead to the production of important phenolic compounds involved in defense responses of a host plant (Weisshaar and Jenkins, 1998). Phenolics that occur constitutively and function as preformed inhibitors are generally referred to as phytoanticipins and those that are produced in response to infection by pathogen called phytoalexins and constitute an active defense response. Phytoalexins are low molecular weight substances, essentially absent from uninfected plant tissues, but accumulating upon infection by pathogenic organisms (Kuc, 1995). In resistant plant, phenolic based defense responses are characterized by the early and rapid accumulation of phenolics at the infection site resulting in the effective isolation of the pathogen (Cherif et al., 2007). The de novo synthesis and differential accumulation of antimicrobial phytoalexins in incompatible and compatible plant-pathogen interactions play crucial roles in the specificity of host’s resistance (Barz et al., 1990).
The best-characterized phenylpropanoid derived phytoalexins are the pterocarpans, isoflavans and isoflavanones of legumes including chickpea, bean, alfalfa, pea and soybean (Dixon et al., 2002). Pterocarpans constitute a group of natural isoflavonoids, which are typically involved as phytoalexins in the defense against pathogens of leguminous plants. Medicarpin and maackiain are two main phytoalexins in chickpea. In contrast to many other isoflavonoids, pterocarpans are mostly not constitutively expressed but induced by biotic and abiotic stresses (Dixon et al., 2002). In this study we were interested in the role of induced defense responses in chickpea against A. rabiei and examined the expression patterns of genes encoding four key enzymes representing various part of A. rabiei induced phenylpropanoid pathways. Transcripts of these genes including one enzyme of the central pathway, Phenylalanine Ammonia-Lyase (PAL), three enzymes of flavonoid/isoflavonoid pathway including chalcone synthase (CHS), Isoflavone reductase (IFR) and Flavanone 3-Hydroxylase (F3H) were compared in resistant and susceptible chickpea genotypes over a range of post-inoculation time points.
MATERIALS AND METHODS
Plant material and fungal isolates: This research was started in October of 2008 in Department of Biotechnology and Plant Breeding, College of Agriculture, Ferdowsi University of Mashhad and was terminated in March of 2009. Seeds of the resistant (Kc 218848) and susceptible (Kc 217854) chickpea genotypes obtained from National Plant Gene Bank of Iran were surface sterilized with 70% ethanol for 30 sec and washed three times with distilled water for 2 min and soaked in water overnight. After germination in 2 days, five seedlings were transplanted in pots with 20 cm diameter in a mixture (2:1 ration) of sterilized soil and sand. Plants were grown in the greenhouse at 20±2°C with natural day light. Two weeks old plants were inoculated with pycnidiospore suspension of 2x105 spore mL-1 harvested from 14 days old culture of race 3 of A. rabiei on Chickpea Seed Meal Agar (CSMA) medium. Plants were inoculated by spraying approximately 5 mL of the spore suspension per plant with a plastic sprayer until run-off. Control plants were sprayed with sterile distilled water. Infected and uninfected plants were placed in plastic boxes for 3 days at high humidity (>90%). Relative humidity was gradually decreased to approximately 75% by opening the boxes from lower edges under the greenhouse conditions mentioned earlier.
Total RNA extraction and cDNA synthesis: Total RNA was extracted from infected and uninfected plants at 0, 3, 6, 12, 24, 48, 72 and 168 h after inoculation with RNXTM plus kit (Cinagen Inc., Iran) according to the method recommended by the manufacturer. In order to minimize the variation in gene expression among individual plants in each line, leaf and stem tissues collected from 4 to 5 individuals were homogenized together for RNA isolation. Extracted total RNAs (30 μg per reaction) were treated with DNaseI to remove DNA contaminants. Equal amount of the DNA-free RNAs (5 μg per reaction) for each time-course was used for cDNA synthesis. One microgram oligo dT18 primer was added to RNA in a microtube and incubated for 5 min at 70°C followed by immediate cooling on ice. Tris buffer (10 mM), dNTP (1 mM), RNAse inhibitor were then added and incubated at 37°C for 5 min. After adding 200 Units of M-MuLV enzyme to the cocktail, the reaction mixture was incubated for 120 min at 42°C. Then it was incubated at 70°C for 10 min in order to inactivate the enzyme. The cDNAs were stored in 20°C until use.
Semi-quantitative PCR: Relative Semi-quantitative PCR was performed to determine RNA expression levels of the four phenylpropanoid pathway genes (PAL, CHS, IFR and F3H) and a constitutively expressed chickpea Ubiquitin gene (as an internal control) using specific primers (Table 1). Each PCR was performed in a total volume of 25 μL containing 10 mM Tris-HCl pH 8.3, 50 mM KCl, 2 mM MgCl2, 200 μm each of the four dNTPs, 2 μL cDNA and 1 U Taq DNA polymerase (Fermentas, USA). The reaction cycling condition consisted of a 2 min initial denaturation at 94°C followed by 28 cycles of reaction with denaturation for 30 sec at 94°C, annealing for 30 sec at 55°C and extension for 1 min at 72°C and finally elongation at 72°C for 5 min. Agarose gel (1.5%) electrophoresis was used to observe semi-quantitative PCR products. All experiments were repeated at least 2 times to verify the results (Bustin, 2002).
Quantification of amplified products was performed by Total-Lab software. The intensities of the DNA bands amplified from genes of interest were normalized relative to that of ubiquitin gene by dividing the former to the latter (Fig. 1).
| Table 1: | Gene-specific primers to amplify the phenylpropanoid pathway related genes in chickpea |
 | |
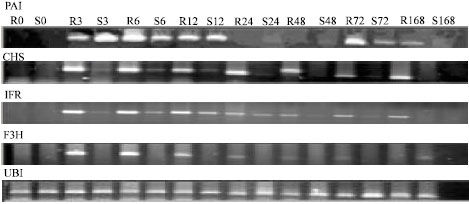 | |
| Fig. 1: | Differential gene expression of phenylpropanoid pathway genes and Ubiquitin in chickpea genotypes after Inoculation with A. rabiei race 3 |
RESULTS AND DISCUSSION
Environmental stimuli such as pathogen attack may rapidly change plant gene expression. Accumulated transcript levels are good indicators of de novo gene expression for synthesis of many compounds including plant defensive arsenal like enzymes leading to biosynthesis of phytoalexins (Ni et al., 1996). The rapidity of phytoalexin accumulation is associated with the type and intensity of plant response to the pathogen (Barz et al., 1990; Cramer et al., 1985; Kuc, 1995). The general phenylpropanoid pathway starting from phenylalanine to coumaroyl-CoA is initiated by the enzyme Phenylalanine Ammonia-Lyase (PAL). Phenylpropanoid metabolism branches to give rise to numerous compounds, many of which are specific to particular plant species. An important branch leads to the production of flavonoids and isoflavonoids. The first committed step in these pathways is catalyzed by chalcone synthase (CHS) (Weisshaar and Jenkins, 1998).
Inoculation of chickpea plants with Ascochyta rabiei, race 3, resulted in rapid up regulation of PAL transcript in both resistant and susceptible genotypes, while no expression (transcript) of PAL was detected in control plants over time-course of sampling. Exposure of chickpea genotypes to the A. rabiei infection resulted in 2-fold increase in PAL expression intensity in susceptible cultivar compared to resistant genotype. Six hours after infection, expression of PAL elevated in resistant cultivar and at this time its intensity was 2-fold above that in susceptible genotype. Twelve hours post inoculation, the expression level of PAL was approximately similar in both resistant and susceptible cultivars. 24 and 48 h after infection, expression of gene down regulated, however gene expression in susceptible genotype was higher than resistance one. Seventy two hours after infection, again expression of PAL dramatically up-regulated and resulted in 3-fold increase in resistant genotype than susceptible one. Really, expression pattern of PAL in resistant genotype was biphasic and showed two peaks in 6 h and 72 h after infection. Afterwards, PAL gene expression down-regulated in resistance cultivar. However, the intensity of gene expression still was higher in resistance cultivar with compared to susceptible one (Fig. 2a).
These results indicated that the production of PAL, leading to phenylpropanoid biosynthesis in chickpea, is an early defense response against Ascochyta rabiei, race 3.
The first enzyme of phenylpropanoid biosynthesis, PAL, has been studied extensively in many host-pathogen combinations.
PAL is the first enzyme of phenylpropanoid metabolism and its activity is often linked with high production of phenols, phytoalexins and lignins which are factors contributing to the resistance of the plant cell. Therefore, high PAL activity is considered as an indicator of resistance in many host-parasite combinations (Zhang et al., 1997). Pre-treatment of chickpea seedlings with Rhizobium isolates before inoculation with Fusarium oxysporum f. sp. ciceris (Foc) race 0 enhanced the accumulation of the phenylalanine ammonia-lyase (PAL), chalcone synthase (CHS) and isoflavone reductase (IFR) genes involved in phenylpropanoids metabolism (Arfaoui et al., 2007).
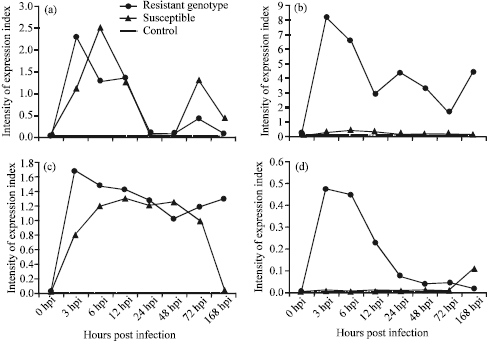 | |
| Fig. 2: | Gene expression pattern of 4 phenylpropanoid pathway genes in aschochyta blight infected chickpea. (a) PAL, (b) CHS, (c) IFR and (d) F3H |
The enzyme was rapidly synthesized de novo and accumulated in elicitor stimulated bean cultivars (Cramer et al., 1985) and in fungal treated cell cultures of carrot (Kurosaki et al., 1986). Increased PAL activity was observed in soybean hypocotyls and leaves in response to glucan elicitor isolated from the fungus Phytophtora megasperma (Kimpel and Kosuge, 1985). PAL activity was observed in Brassica napus at 12 h after inoculation with Leptosphaeria maculans (Chakraborty et al., 1993).
Chalcone synthase was differentially expressed in resistant and susceptible genotypes. A dramatic increase was observed in CHS gene expression 3 h post inoculation followed by gradual decrease of transcript level until 24 h post inoculation. However, an unexpected increase of transcript level was observed in 24 and 168 h post inoculation time points. In susceptible cultivar, the CHS transcript did not significantly increase after inoculation compared to resistant genotype. Also in control plants there was no detectable expression of CHS transcripts over time curse of sampling (Fig. 2b).
In legumes both PAL and CHS enzymes are encoded by multi gene families and most, if not all, of these genes are induced in response to pathogen attack. It is therefore possible that very rapid and massive up regulation of these transcripts immediately after inoculation with A. rabiei could be due to activation of one or a sub set of these genes and late up regulation of those may be caused by late activation of other members of the gene family (Ni et al., 1996).
In cell-suspension cultures of two chickpea cultivars, the maximum of the elicitor-induced CHS mRNA activity was determined 4 h after onset of induction in the culture of both cultivars, although in cells of resistant cultivar this mRNA activity was induced at a level two-fold higher than in cells of the susceptible (Daniel and Barz, 1990).
Lawton and Lamb (1987) showed that induction of PAL and CHS genes occurred in wounded and infected hypocotyls during race-cultivar-specific interaction of bean and fungus Colletotrichum lindemuthranum, the causal agent of anthracnose (Lawton and Lamb, 1987).
Isoflavone reductase (IFR) is an enzyme that first cloned from legumes and specifically involved in the branch of isoflavonoid biosynthesis pathway leading to isoflavane and petrocarpan phytoalexins in chickpea (Dixon et al., 2002). In resistance genotype, accumulation of IFR transcripts increased 3 h after inoculation and remained elevated up to 6 h after elicitation, while in control plants no IFR transcript was detected over time-course of sampling. Afterward, expression of IFR gradually decreased, although the change in transcription rate of IFR gene in resistant genotype was biphasic and again up-regulated 72 h after infection (Fig. 2c).
In susceptible cultivar, the maximum gene expression of IFR delayed by 12 h after inoculation and then gradually decreased. However, 48 h after infection gene expression intensity increased. At this time IFR gene expression in susceptible genotype was even higher than resistant one (Fig. 2c).
Tiemann et al. (1991) also showed that IFR enzyme is induced to a pronounced higher degree in a resistant chickpea cultivar than in susceptible ones upon treatment with A.rabiei elicitor (Tiemann et al., 1991). Rapidly increased higher levels of PAL, CHS and IFR enzymes were observed with increased levels of their transcription in elicited cell suspension cultures of Medicago sativa (Ni et al., 1996).
Flavanone 3-Hydroxylase (F3H) enzyme which plays a key role at the point of branch diverging either to isoflavonoid or flavonoid metabolism leading to accumulation of phytoalexin or anthocyanin respectively. These two biosynthesis pathways mediated by isoflavone synthase (IFS) and F3H compete for flavanones as common substrate (Cho and Muehlbauer, 2005). F3H was up regulated very rapidly 3 h post inoculation with A. rabiei, race 3, followed by gradual decrease after 12 h. Contrary to the expression pattern of F3H in the resistant genotype, F3H was not detected in susceptible genotype until seven days after inoculation (Fig. 2d). Based on these results, induction of F3H at a late infection time did not confer resistance in chickpea while higher expression of F3H immediately after inoculation with fungus appeared to be a critical factor for blight resistance.
In plant defense responses, the extent and time of expression of defense related genes are two vital factors in discrimination of resistant from susceptible genotype.
Upon treatment of chicpea with A. rabiei elicitor, a pronounced induction of these enzymes was also observed in the resistant cell lines than in the susceptible ones (Gunia et al., 1991). Results obtained in this study provides molecular evidence confirming earlier reports indicating effects of flavonoids on resistance to fungi in crop plants particularly, in chickpea for resistance to A. rabiei (Cho and Muehlbauer, 2005).
Armero et al. (1993) and Cachinero et al. (2002) emphasized that key enzymes that involved in phenylpropanoid and isoflavonoid pathways play a crucial role in biological control of chickpea diseases caused by antagonistic microorganisms.
The present data suggest that phenylpropanoid metabolism is enhanced in resistant chickpea variety (Kc 218848) immediately after inoculation with race 3 of A. rabiei which finally leads to the accumulation of constitutive phenols and phytoalexins. The phytoalexin accumulated in resistant plant cells are in enough amounts to inhibit the extension of the invasive mycelia and play an important role in the first step of infection in chickpea plants. It suggested that accumulation of isoflavones and phytoalexins may be inhibited or degraded more rapidly in susceptible genotypes of chickpea than resistant ones. In susceptible plants, products of defense mechanism might be broken down by the fungus (Nasir et al., 2000). Higher accumulation of transcripts of intermediate enzymes involved in production of phytoalexins in resistant genotype resulted in the faster defense response in resistant variety than in susceptible one.
REFERENCES
- Arfaoui, A., A. El-Hadrami, Y. Mabrouk, B. Sifi and A. Boudabous et al., 2007. Treatment of chickpea with Rhizobium isolates enhances the expression of phenylpropanoid defense-related genes in response to infection by Fusarium oxysporum f. sp. ciceris. Plant Physiolo. Biochem., 45: 470-479.
CrossRef - Armero, J., F. Cabello, J.M. Cachinero, R. Lopez-Valbuena. J. Jorrin, R.M. Jimenez-Diaz and M. Tena, 1993. Defense Reaction Associated to Host-Nonspecific and Host-Specific Interactions in the Chickpea (Cicer arietinum L.)-Fusarium oxysporum Pathosystem. In: Mechanism of Plant Defense Response, Fritig, B. and M. Legrand (Eds.). Kluwer Academic Publishers, Dordrecht, Boston, London, pp: 316-319.
- Barz, W., W. Bless, G. Borger-Papendorf, W. Gunia, U. Mackenbrock, D. Meier, Ch. Otto and E. Super, 1990. Phytoalexins as Part of Induced Defense Reaction in Plants: Their Elicitation, Function and Metabolism. In: Bioactive Compounds from Plants, John Wiley and Sons (Eds.). A Wiley Interscience Publication, New York, pp: 140-155.
- Bustin, S.A., 2002. Invited review: Quanitification of mRNA using real-time reverese transcription PCR (RT-PCR): Trends and problems. J. Mol. Endocrinol., 29: 23-39.
Direct Link - Cachinero, J.M., A. Hervas, R. M. Jimenez-Diaz and M. Tena, 2002. Plant defense reactions against Fusarium wilt in chickpea induced by incompatible race 0 of Fusarium oxysporum f. sp. ciceris and nonhost isolates of F. oxysporum. Plant Pathol., 51: 765-776.
Direct Link - Cho, S. and F.J. Muehlbauer, 2004. Genetic effect of differentially regulated response genes on resistance to necrotrophic fungal pathogens in chickpea (Cicer arietinum L.). Physiol. Mol. Plant Pathol., 64: 57-66.
CrossRef - Cho, S., W. Chen and F.J. Muehlbauer, 2005. Constitutive expression of the flavanone 3-hydroxylase gene related to pathotype-specific ascochyta blight resistance in Cicer arietinum L. Phys. Molecular Pathol., 67: 100-107.
CrossRef - Coram, T.E. and Pang, E.C.K, 2007. Transcriptional profiling of chickpea genes differentially regulated by salycilic acid, methyl jasmonate and aminocyclopropan carboxylic acid to reveal pathways of defense-related gene regulation. Functional Plant Bio., 34: 52-64.
Direct Link - Cramer, C.L., J.N. Bell, T.B. Ryder, J.A. Bailey and W. Schuch et al., 1985. Co-ordinated synthesis of phytoalexin biosyntheyic enzymes in biologically-stressed cells of bean (Phaseolus Vulgaris L.). The Embo J., 4: 285-289.
PubMed - Dixon, R.A., L. Achnine, P. Kota, C.J. Liu, M.S.S. Reddy and L.J. Lang, 2002. The phenylpropanoid pathway and the plant defence: A genomic perspective. Mol. Plant Pathol., 3: 371-390.
CrossRefDirect Link - Gunia, W., W. Hinderer, U. Wittkampf and W. Barz, 1991. Elicitor induction of cytochorome P-450 monooxygenase in cell suspension cultures of chickpea (Cicer arietinum L.) and their involvement in pterocarpan phytoalexin biosynthesis. Z. Naturfosch, 46: 58-66.
Direct Link - Kuc, J., 1995. Phytoalexins, stress metabolism and disease resistance in plants. Annu. Rev. Phytopathol., 33: 275-297.
CrossRefDirect Link - Kurosaki, F., N. Tashiro and A. Nishi, 1986. Induction of chitinase and phenylalanine ammonia-lyase in cultured carrot cells treated with fungal mycelial walls. Plant Cell Physiol., 27: 1587-1591.
Direct Link - Lawton, M.A. and C.J. Lamb, 1987. Transcriptional activation of plant defense genes by fungal elicitor, wounding and infection. Mol. Cell. Biol., 1: 335-341.
PubMed - Ni, W., T. Fahrendorf., G.M. Balance, C.J. Lamb and R.A. Dixon, 1996. Stress responses in alfalfa (Medicago sativa L.). XX. Transcriptional activation of phenylpropanoid pathway genes in elicitor-induced cell suspension cultures. Plant Molecular Bio., 30: 427-438.
PubMedDirect Link - Tiemann, K., D. Inze., M. van Montagu and W. Barz, 1991. Pterocarpan phytoalexin biosynthesis in elicitor-challenged chickpea (Cicer arietinum L.) cell cultures. Eur. J. Biochem., 200: 751-757.
CrossRef - Weisshaar, B. and G.I. Jenkins, 1998. Phenylpropanoid biosynthesis and its regulation. Curr. Opin. Plant Biol., 1: 251-257.
Direct Link - Zhang, I., M.P. Robbines, I.I.W. Carver and R.J. Zeyen, 1997. Induction of phenylpropanoid gene transcript in oat attached by Erysiphe graminis at 20 ◦C and 10◦C. Physiol. Mole. Plant Pathol., 51: 15-33.
CrossRefDirect Link