
S.F. Hwang
Alberta Agriculture and Rural Development, Crop Diversification Centre North, Edmonton, AB T5Y 6H3, Canada
H.U. Ahmed
Alberta Agriculture and Rural Development, Crop Diversification Centre North, Edmonton, AB T5Y 6H3, Canada
B.D. Gossen
Agriculture and Agri-Food Canada, Saskatoon Research Centre, Saskatoon, SK S7N 0X2, Canada
H.R. Kutcher
Agriculture and Agri-Food Canada, Saskatoon Research Centre, Saskatoon, SK S7N 0X2, Canada
S.A. Brandt
Agriculture and Agri-Food Canada, Saskatoon Research Centre, Saskatoon, SK S7N 0X2, Canada
S.E. Strelkov
Department of Agricultural, Food and Nutritional Science, University of Alberta, Edmonton, AB T6G 2P5, Canada
K.F. Chang
Alberta Agriculture and Rural Development, Field Crop Development Centre, Lacombe, AB T4L 1W8, Canada
G.D. Turnbull
Alberta Agriculture and Rural Development, Crop Diversification Centre North, Edmonton, AB T5Y 6H3, Canada
Plant Pathology Journal
Year: 2009 | Volume: 8 | Issue: 3 | Page No.: 106-112







ABSTRACT
Impact of long-term crop rotations on populations of soilborne pathogens in the genera Fusarium, Pythium and Rhizoctonia and on canola seedling establishment and development was evaluated under controlled conditions. Soil samples were collected from two crop rotation experiments conducted at two sites in Saskatchewan, Canada. A part of the 2006-soil sample of each rotation was sterilized to compare canola seedling growth with and without soilborne pathogens. With 2007-soil, the sterilization treatment was replaced with a fungicide seed treatment (Apron Maxx) to assess the potential to reduce seedling diseases. Populations of Fusarium, Pythium and Rhizoctonia sp. were estimated in the soil of each rotation using dilution plating onto selective media for each fungus. Higher seedling emergence and increased growth of canola were obtained in the sterilized soil or due to seed treatment. Fusarium was the predominant genus followed by Pythium and Rhizoctonia sp. in the soil of both sites. This study suggests that diverse crop in the rotation may reduce the populations of Fusarium, Pythium and Rhizoctonia in the soil and may contribute to improve the overall growth of canola.
PDF Abstract XML References Citation
How to cite this article
S.F. Hwang, H.U. Ahmed, B.D. Gossen, H.R. Kutcher, S.A. Brandt, S.E. Strelkov, K.F. Chang and G.D. Turnbull, 2009. Effect of Crop Rotation on the Soil Pathogen Population Dynamics and Canola Seedling Establishment. Plant Pathology Journal, 8: 106-112.
DOI: 10.3923/ppj.2009.106.112
URL: https://scialert.net/abstract/?doi=ppj.2009.106.112
DOI: 10.3923/ppj.2009.106.112
URL: https://scialert.net/abstract/?doi=ppj.2009.106.112
INTRODUCTION
Canola (Brassica napus L. and Brassica rapa L.) is one of the major oilseed crops grown worldwide. The value of canola crops has increased recently, partly as a result of increased use of canola as biofuel (Lennox et al., 2007). The increased value of the crop has resulted in increased acreage and has exacerbated an existing trend towards shortened intervals between canola crops in crop rotations. Moreover, the availability of herbicide tolerant canola cultivars with strong blackleg resistance has facilitated this change. However, repeated cultivation of a susceptible crop can lead to increased populations of many pathogens. For example, blackleg disease incidence (Leptoshaeria maculans (Desm.) Ces. Et de Not.) was greater when canola was gown for four years consecutively (Guo et al., 2005). Among the numerous soil-borne pathogens Fusarium sp., Pythium sp. and Rhizoctonia sp. are persistent, recurrent problems causing canola seedling blight and damping off reducing crop stand establishment in the Canadian prairies. Disease incidence due to these pathogens was reported to be approximately 10% for several years at some locations in Saskatchewan (Pearse et al., 2004, 2005) and in central Alberta over 20% of fields surveyed in 2005 and 2006 suffered from poor seedling establishment (D. Orchard, pers. comm.). Seed treatment with fungicides is not always effective for the control of soilborne diseases and integrated, sustainable disease management options are desirable.
Long-term crop rotations have been used as a standard practise to reduce disease pressure in field crops. Krupinsky et al. (2002) reported that longer crop rotations contribute to the decline of pathogen populations. Crop rotation was found to break the associations between a host and a pathogen for effective control of both foliar and soil borne crop diseases like fusarium head blight (Gibberella zeae [Schwein.] Petch) (Dill-Macky and Jones, 2000), tan spot (Pyrenophora tritici-repentis (Died.) Drechs. (Bockus and Claassen, 1992) of wheat, common root rot (Cochliobolus sativus (Ito and Kuribayashi.) Drechs. ex Dast) of wheat (Bailey et al., 1992) and charcoal rot (Macrophomina phaseolina [Tassi.] Goid) of soybean (Wrather and Kending, 1998). A long term rotation (>5 years) was found to be effective in reducing the incidence of blackleg (Guo et al., 2005). Studies conducted over several years with different potato cropping systems indicated consistent reduction of soilborne disease caused by Rhizoctonia sp. when canola or rapeseed was grown prior to potato (Larkin and Griffin, 2006). Avoiding susceptible host crops in the rotation was found effective to reduce the impact of soilborne R. solani (Peel, 1998). The purpose of this study was to determine the impact of long-term crop rotations on soil pathogen populations (Fusarium, Pythium and Rhizoctonia sp.) and on the growth parameters of canola.
MATERIALS AND METHODS
Site, crop rotation and soil sampling: Long-term crop rotation experiments were established in 1998 at Scott and 1999 at Melfort, Saskatchewan, Canada under field conditions. The study locations represent the Moist Dark Brown soil zone at Scott and the Moist Black soil zone at Melfort (Kutcher and Brandt, 2006). Soil samples were collected from the plots of different crops in the rotation representing three replications of the trials from both the sites Melfort and Scott, Saskatchewan in 2006 and from Scott only in 2007 as the Melfort site was abandoned. The soil samples were obtained from the 8-year phase of the rotation at Melfort and the 9 and 10-year phase at Scott in 2006 and 2007, respectively. Two to 10 crop rotation cycles had been completed, depending on the number of crop(s) in a rotation, site and year. The crops included in the rotations were canola (Brassica napus L.), flax (Linum usitatissimum L.), pea (Pisum sativum L.) and wheat (Triticum aestivum L.). The rotations were continuous canola (C-C-C-C), continuous field pea (P-P-P-P), canola-wheat (C-W), pea-wheat (P-W), pea-canola-wheat (P-C-W), canola-wheat-pea-wheat (C-W-P-W) and canola-wheat-flax-wheat (C-W-F-W).
A soil probe was used to collect five soil samples from the top 15-20 cm for a total of approximately 2.0 kg of soil from each of the selected plots. A total of 21 soil samples were collected (3 replicationsx7 crop rotations). The samples were mixed thoroughly to make composite samples. To avoid cross-contamination the soil sampler was cleaned and decontaminated with a 70% ethanol spray, then dried after collecting samples from each plot. Plant debris was removed from the field soil samples using a sieve. A sub-sample (20 g) from each of these soil samples was air-dried for 3-4 weeks at room temperature and then finely ground using a mortar and pestle.
Determination of soil fungal population: Ten grams of ground composite soil samples were added to sterile 0.1% water agar medium to obtain a 10X dilution and was further diluted to 50X and 250X. Three selective growth media were used to isolate the soil fungi: MPVM agar (Mircetich, 1971), which is selective for Pythium sp.; PCNB medium (Nash and Snyder, 1962), which is selective for Fusarium sp. and Ko and Hora (1971) medium, which is selective for Rhizoctonia sp.
Using a 1000 μL pipette, 0.5 mL samples of the 10X, 50X and 250X dilutions were plated onto the respective selective media for Rhizoctonia, Pythium and Fusarium sp., using 4 or 5 Petri dishes (replicates) for each sample. The dishes were incubated at room temperature on a laboratory bench under natural light. Colony counts were performed after 3-4 days for Pythium sp. and after 5-7 days for Fusarium sp. and Rhizoctonia sp. For colony counting, dilutions of 50X, 100X and 250X were used for Pythium sp., Fusarium sp. and Rhizoctonia sp., respectively.
Greenhouse bioassay: The remainder of each of the soil samples was used for the greenhouse experiment. Ten seeds of canola cv. Westar were sown into eight replicate cups (~300 mL, 7 cm diameter), which had been perforated in the bottom for drainage. The experiments conducted with the 2006 soil samples were arranged in a split-plot design, with location as the main plot and the rotations in the sub-plots. The experiments were conducted in both sterilized and non-sterilized siol samples. With the 2007 soil samples the experiment was laid out following a randomized complete block design with eight replicate cups for each soil sample. Seed treated with Apron Maxx or non-treated seed was sown into non-sterilized soil. The experiments were conducted under greenhouse conditions with a 16 h photoperiod with 20°C day/16°C night temperatures.
Seedling emergence was assessed 2 weeks after seeding. Seedling height was measured at 3 weeks, at which time the seedlings were removed from the soil. The roots were washed under running tap water and root vigour and shoot dry weight recorded. Root vigour was evaluated on a 1-4 scale (where: 1 = Poor; 2 = Fair; 3 = Fairly good and 4 = Good).
Data analysis: Data from the soil samples collected from Melfort and Scott in 2006 were combined prior to Analysis of Variance (ANOVA) after exploratory data analysis revealed homogeneity of the variance between sites. The combined data were analyzed using the General Linear Model (GLM) procedure of SAS statistical software (SAS Institute Inc, 2005), with site, replication and crop rotations as independent variables and plant growth parameters (seedling emergence, root vigour and shoot weight) as the response variables. As a result of a significant site effect and sitexrotation interactions for some plant growth parameters, data from 2006 were analyzed separately for each site. When the 2006 and 2007data from Scott were combined and analyzed, there was significant year effect and yearxrotation interaction. In addition, single degree of freedom contrasts were performed to assess the effect of crop rotations. Separate ANOVA and linear contrasts were performed for fungal populations. These pre-planned contrasts were chosen to compare the impact of intensive production of canola and field pea on soil mycoflora and canola growth parameters. The contrasts compared the 4 year rotations (C-W-F-W and C-W-P-W) vs. continuous canola (C-C-C-C), the 4 year rotations vs. continuous pea (P-P-P-P); 4-year rotations vs. rotations of 3 and 2 year (P-C-W, C-W and P-W); the 3 year rotation (P-C-W) vs. 2-year rotations (C-W and P-W); the 3 year rotation (P-C-W) vs. continuous canola; P-C-W vs. continuous pea; and continuous canola vs. continuous pea. The means comparisons for site effects, soil sterilization effects and seed treatment effects were performed using Least Significance Difference (LSD) test at p≤0.05 significant levels.
RESULTS
Statistical analysis: The analysis of variance indicated a significant site effect (p≤0.01) with respect to seedling emergence and root vigour. In 2006, seedling emergence and root vigour were higher in the soil from Melfort than from Scott (Table 1). The effect of soil treatment (sterilized and non-sterilized, 2006) or seed treatment (treated with Apron Maxx or non-treated, 2007) was significant (p≤0.01) for most of the growth parameters. As expected, seedling emergence was higher in sterilized versus non-sterilized soil from both sites (Table 1). Sterilization also affected shoot dry weight but not root vigour in 2006. In the soil from Scott in 2007, seed treatment with Apron Maxx increased seedling emergence, shoot dry weight and root vigour compared to the non-treated seed (Table 1).
Populations of Pythium sp. and Rhizoctonia sp. were higher (p≤0.01) in soil from Melfort than from Scott and there was no difference in the incidence of Fusarium sp. (Table 1). The effect of crop rotation was significant (p≤0.01) on populations of Fusarium and Pythium sp., but not Rhizoctonia.
The effect of crop rotations was significant (p≤0.01) for all plant growth parameters in both the Melfort and Scott soils in 2006. In soil collected in 2007 from Scott, the rotation effect was significant (p≤0.01) for shoot dry weight and seedling height.
Seedling emergence: For soil collected in 2006, seedling emergence was higher in the three-crop rotation (P-C-W) compared to the two-crop rotations (C-W, P-W) in sterilized soil (Table 2). The three-year rotation also had higher emergence counts than the continuous canola (C-C-C-C) or pea (P-P-P-P) rotations in sterilized soil from Melfort, although the differences were not significant. Compared to continuous pea, the continuous canola had higher seedling emergence (Table 2).
In soil from Scott in 2006, no significant differences in seedling emergence were detected in any of the contrasts for sterilized soil (Table 3). In non-sterilized soil, seedling emergence was higher in soil seeded to continuous canola compared to the four-crop rotation.
| Table 1: | Effects of site, soil and seed treatments on canola emergence, root vigour and shoot weight and effects of site on populations of Fusarium, Pythium and Rhizoctonia |
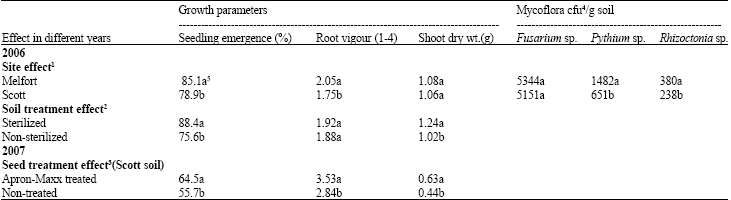 | |
| 1Growth parameter data for the site effect are the means of 8 replicates x7 different crop rotation soils x2 soil treatments (sterilized and non-sterilized). 2Growth parameter data for the soil treatment effect are the means of 8 replicates x7 different crop rotation soils x2 sites. 3Growth parameter data for the seed treatment effect are the means of 8 replicate cups x7 different crop rotation soils x2 sites. Cfu4: Colony forming unit. 5Values followed by the same letter are not significantly different at p≤0.05 by LSD test | |
| Table 2: | Linear contrasts of the effect of soils from different crop rotations, collected from Melfort, Sastatchewan in 2006, on emergence, root vigour and shoot weight of canola |
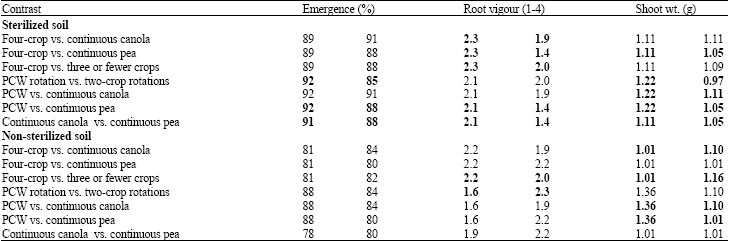 | |
| Bold values are significantly different at p≤0.05 based on linear contrasts | |
| Table 3: | Linear contrasts of the effect of soils from different crop rotations, collected from Scott, Saskatchewan in 2006 on emergence, root vigour and shoot weight of canola |
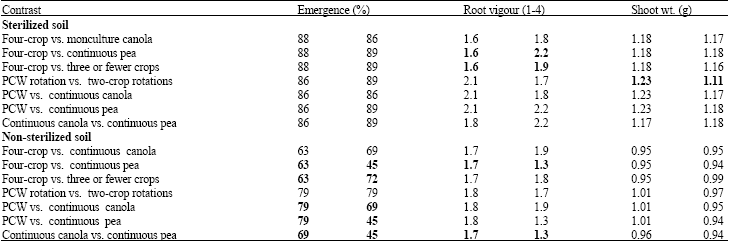 | |
| Bold values are significantly different at p≤0.05 based on linear contrasts | |
However, canola emergence was lower in soil that had been sown to continuous pea than in soil from the four-crop rotation. Continuous canola resulted in higher emergence than continuous pea. Soils from rotations of three or fewer crops had higher emergence than the four-crop rotation. The three-crop (P-C-W) rotation had higher emergence than continuous canola or pea.
In soil from Scott in 2007, seedling emergence from non-treated seed was higher in the continuous pea and three or fewer crop rotations compared to the four-crop rotation (Table 4). Emergence from seed treated with Apron Maxx was significantly higher in soil taken from continuous canola or pea compared to the three-crop rotations (Table 4).
Root vigour: In sterilized soil, root vigour was higher in Melfort soil collected from the four-crop rotations than from the other rotations (Table 2). Similarly, root vigour was higher in soils obtained from three-crop rotations compared to two-crop rotations or continuous cropping. Root vigour was also higher in the soil from continuous canola than from continuous pea. In contrast, root vigour in sterilized soil from Scott was lower in the W-W-W-W rotation compared to other rotations (Table 3). In soil from Scott in 2007, root vigor for fungicide-treated seed was significantly higher in continuous canola or pea, or in the two-year rotations compared to P-C-W (Table 4).
Shoot weight: Shoot weight was significantly higher in the four-crop rotation compared to continuous cropping of pea in sterilized soil from Melfort collected in 2007. Shoot weight was also significantly higher in the soil from three-crop rotations compared to other rotations. Soil collected from continuously cropped canola had higher shoot weight compared to continuously cropped pea.
In non-sterilized soil, shoot weight was higher in the three-crop rotation than in soil from 4 or 2 year rotations or from continuous pea cropping (Table 2). In sterilized soil from Scott in 2006, shoot weight was higher in the three-crop rotation compared to the two-crop rotations (Table 3).
| Table 4: | Linear contrasts of the effect of soils from different crop rotations, collected from Scott, Saskatchewan in 2007 on emergence, root vigour and shoot weight of canola |
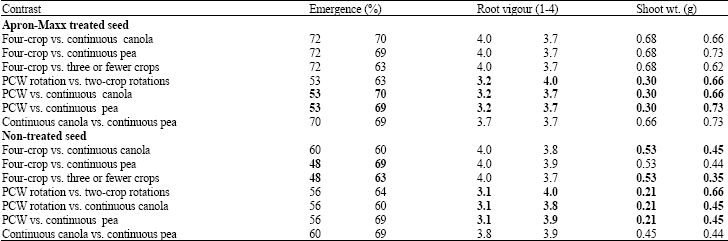 | |
| Bold values are significantly different at p≤0.05 based on linear contrasts | |
| Table 5: | Linear contrasts of the effect of crop rotation on soil mycoflora in soil samples collected from long-term crop rotation experiments at Melfort and Scott, Saskatchewan in 2006 and 2007 |
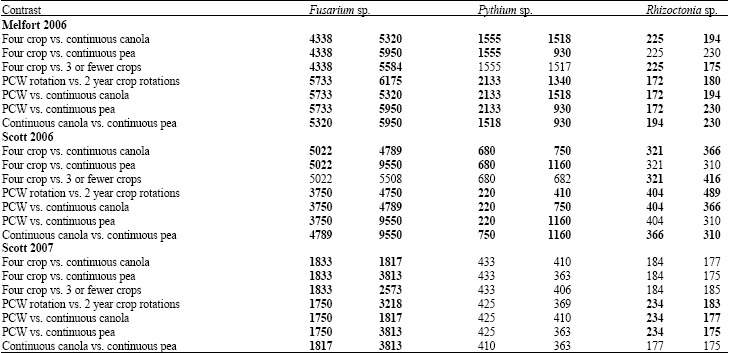 | |
| Bold values are significantly different at p≤0.05 based on linear contrasts | |
Shoot weight of fungicide-treated seed in soil from Scott in 2007 was significantly higher in the two-crop rotations or the continuous cropping of canola or pea compared to the soil from the three-crop rotation (Table 4). When non-treated seed was used, the four-crop rotations had a higher shoot weight than the other rotations. Similar to results from the fungicide-treated seed, shoot weight was greater in the two-crop rotations and the continuous crops than in the 3 year rotation (Table 4).
Fungal populations: In 2006, Fusarium sp. was the most commonly isolated pathogens, followed by Pythium sp. and Rhizoctonia sp. in soil from both Melfort and Scott (Table 5). There were more fungal isolates collected in 2006 than in 2007 from the Scott soil and lower populations of each of the pathogenic fungi in the four-crop rotations compared to the other rotations with a few exceptions in case of the soil from Scott in 2006 (Table 5).
The population of Fusarium sp. was consistently higher in soil from continuous pea plots compared to continuous canola. Populations of Pythium sp. were lower in soil from the four-crop and three-crop rotations than in soil from the continuous crops at Scott in 2006. The trends were reversed at Melfort in 2006 for the four-crop rotations compared to the continuous crops, but similar results were observed for the three-crop rotations compared to the continuous crops or 2 year rotations. No significant differences among rotations in populations of Pythium sp. were detected at Scott in 2007.
DISCUSSION
In this study, seedling emergence and other growth parameters were higher in sterilized versus non-treated soil and in fungicide-treated versus non-treated seed, indicating that the pathogens in the soil samples reduced canola emergence and growth. Sterilization influences a variety of soil properties and makes some nutrients more available than others. The growth of canola in sterilized soil may have been affected by these alterations, but the large differences in emergence and shoot weight of canola, associated with the fungicide seed treatment in soil from Scott in 2007, confirm that the most important factor across all sites was the impact of soil-borne pathogens.
As expected, the most frequently isolated microorganisms with the potential for pathogenicity were Fusarium, Pythium and Rhizoctonia sp., although there were other fungi in the plates. These genera are known to be associated with seedling blight of canola. Pythium sp. was more frequently isolated at Melfort than Scott, while Rhizoctonia sp. was more common at Scott. This observation is consistent with the literature pertaining to the environmental requirements of each genus of these fungi (Bailey and Lazarovits, 2003; Manian and Manibhushanrao, 1990; Rajan and Singh, 1973). Pythium sp. are more prolific under moist conditions, while Rhizoctonia can survive in drier environments (Willetts, 1971). Melfort usually receives more precipitation than Scott (~425 mm annually compared to ~350 mm), is slightly cooler and the soil has a higher percentage of organic matter. However, the soil is more acidic at Scott than at Melfort (Kutcher and Brandt, 2006), which likely makes it more conducive for Pythium than for Rhizoctonia. The long-term pattern of soil moisture was likely the dominant factor that determined which genera of fungi were predominant at each site. There was no difference between sites for the populations of Fusarium sp.
Diverse crop rotations are known to reduce populations of soil-borne pathogens, but continuous production of any single crop results in increased populations of pathogens and insect pests specific to that crop (Christen and Sieling, 1995). In our study, seedling emergence and growth generally increased with increased diversity of the crop rotation, while populations of soil-borne pathogens declined with increased diversity of rotation. Several of the growth parameters examined under greenhouse conditions showed substantially different trends across sites and years. For example, emergence in soil from continuous canola was sometimes greater than in the four-crop rotation or continuous pea. This may be an artifact of the experiment, resulting from a greater seed bank of volunteer canola in the continuous canola relative to other rotations. However, soil sterilization should have killed any volunteers. One factor that likely contributed to these deviations from that expected in the growth parameters and soil mycoflora is the weed population. In continuous cropping, the weed population is often high and difficult to control and can deplete soil nutrients as well as affect the composition of soil mycoflora (Abdalla and Omer, 1981). Other factors, such as the residual effect of seed and foliar pesticides applied to the various crops, may also account for a portion of this variability. Differences in soil characteristics and weather parameters between sites may have also influenced the results.
This study reinforces the previous findings and concludes that the spectrum of soil-borne pathogens responds to crop rotation and environment. In turn, canola seedling growth is affected by which soil-borne pathogens are present and by their relative abundance. Continuous cropping, particularly of pea, resulted in higher populations of Pythium and Fusarium and in most cases, lower seedling emergence and growth. Populations of Rhizoctonia did not show consistent trends among the rotation types. Further research should be conducted on identification of specific pathogens in soil samples and on the year-to-year dynamics of various soil pathogen populations as various crop rotations progress.
ACKNOWLEDGMENTS
We are grateful to L. Sproule and D. Gerein at Scott and C. Kirkham and D. Cross at Melfort for collecting the soil samples. We thank the Saskatchewan Canola Development Commission, who through the Canola Council of Canada’s Canola Agronomic Research Program, provided partial funding for this project.
REFERENCES
- Abdalla, M.H. and F. Omer, 1981. Studies on the rhizosphere mycoflora of Euphorbia granulata Forsk. and E. heterophylla L. Mycologia, 73: 9-15.
CrossRefDirect Link - Bailey, K.L. and G. Lazarovits, 2003. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res., 72: 169-180.
Direct Link - Christen, O. and K. Sieling, 1995. Effect of different preceding crops and crop rotations on yield of winter oil-seed rape (Brassica napus L.). J. Agron. Crop Sci., 174: 265-271.
CrossRef - Dill-Macky, R. and R.K. Jones, 2000. The effect of crop residues and tillage on Fusarium head blight of wheat. Plant Dis., 84: 71-76.
CrossRef - Guo, X.W., W.G.D. Fernando and M. Entz, 2005. Effects of crop rotation and tillage on blackleg disease of canola. Can. J. Plant Pathol., 27: 53-57.
Direct Link - Ko, W. and F.K. Hora, 1971. Selective medium for the quantitative determination of Rhizoctonia solani in soil. Phytopathology, 61: 707-710.
Direct Link - Krupinsky, J.M., K.L. Bailey, M.P. McMullen, B.D. Gossen and T.K. Turkington, 2002. Managing plant disease risk in diversified cropping systems. Agron. J., 94: 198-209.
CrossRefDirect Link - Willetts, H.J., 1971. The survival of fungal sclerotia under adverse environmental conditions. Biol. Rev., 46: 387-407.
CrossRef - Wrather, J.A. and S.R. Kending, 1998. Tillage effects on Macrophomina phaseolina population density and soybean yield. Plant Dis., 82: 247-250.
CrossRef - Larkin, R.P. and T.S. Griffins, 2007. Control of soilborne potato diseases using Brassica green manures. Crop Prot., 26: 1067-1077.
CrossRef
Umar Sani Abdullahi Reply
I'll try to had your noticing always, because i need to benefits your ideas. Thanks