
Savas Korkmaz
Department of Plant Protection, Faculty of Agriculture, Canakkale Onsekiz Mart University, 17020, Canakkale, Turkey
Serkan Onder
Department of Plant Protection, Faculty of Agriculture, Canakkale Onsekiz Mart University, 17020, Canakkale, Turkey
Mustafa Gumus
Department of Plant Protection, Faculty of Agriculture, Ege University, 35100, Bornova, Izmir, Turkey
Plant Pathology Journal
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 85-91







ABSTRACT
In this study, biological properties and dsRNA patterns of nine Aegean region Turkish isolates of Citrus tristeza virus were investigated. In addition, CTV antigen concentration and effect of tissue sampling time taken from naturally affected Satsuma Owari mandarin plant grown in the field of Balikesir province were also determined. The results indicated that nine isolates graft inoculated to a set of indicator plants did not show any symptoms on sour orange, grapefruit and sweet orange plants or did not cause any stunting when compared with healthy control. However, all isolates showed vein clearing symptoms on Mexican lime. All nine strains showed full length of major dsRNA band of 13.3x106 Da MW on Madam Vinous plant. Moreover, they showed three full lengths of major or minor dsRNA bands with 2.0, 0.8 and 0.5x106 MW. All four different citrus varieties inoculated with KE-5 strain showed full length of major dsRNA. The additional two dsRNA of 0.8 and 0.5x106 MW were also detected as clear as full-length of dsRNA in four hosts, but recovery of these bands were less visible in sour orange. The highest ELISA value was obtained in March and April and then steadily decreased until September and then again gradually increased through December. ELISA value was the lowest in September. DsRNA recovery from a single isolate of CTV EK-2 grown in the field was good in March, April, May and June and it was poor in January, February, July, August, November and December. No dsRNA band was detected in September and October.
PDF Abstract XML References Citation
How to cite this article
Savas Korkmaz, Serkan Onder and Mustafa Gumus, 2008. Investigation of Biological Properties, Double-Stranded RNA Patterns and Antigen Concentration in Satsuma Owari Mandarins Infected with Citrus tristeza virus in Aegean Region of Turkey. Plant Pathology Journal, 7: 85-91.
DOI: 10.3923/ppj.2008.85.91
URL: https://scialert.net/abstract/?doi=ppj.2008.85.91
DOI: 10.3923/ppj.2008.85.91
URL: https://scialert.net/abstract/?doi=ppj.2008.85.91
INTRODUCTION
Citrus tristeza virus (CTV) is a member of the closterovirus with filamentous particle about 2000x11 nm in size which is transmitted infected budwood and spread various aphid species (Yokomi et al., 1994; Rocha-Pena et al., 1995). The virus was first observed in Orient and has been spread by man to the most citrus production regions. It has been epidemics in the western Hemisphere in Brazil and A rgentina in the 1930s (Lee and Rocha-Pena, 1992; Rocha-Pena et al., 1995). CTV and its vector Aphis gossipy Glover have been known in Turkey for many years but no cause any serious losses in the citrus plantation; however, it stands as a potential threat for citrus industry of Turkey because of mostly using sour orange as a rootstock (Cinar et al., 1993; Korkmaz, 2002).
CTV genome is composed of a single stranded, positive sense, RNA of 19296 nucleotides organized in 12 open reading frames which encodes at least 17 proteins (Karasev et al., 1995; Mawassi et al., 1996; Vives et al., 1999). The genomic RNA is encapsidadet by two proteins of 25 and 27 kDa, coating about 95 and 5% of the particle length, respectively (Febres et al., 1996). Additionally, infected plant extracts usually contain at least nine subgenomic RNAs that are 3’coterminal (Hilf et al., 1995) and the changeable number of defective RNAs that contain both genomic RNA termini but lack variable portions of the central region (Mawassi et al., 1995; Ayllon et al., 1999).
CTV has many different strains causing diversity of symptoms associated with CTV infection on different citrus hosts. Mild strains of CTV produce basically no noticeable symptoms in their hosts except mild vein clearing on Mexican lime. Severe strains produce a range of symptoms including seedling yellows, decline on sour orange rootstock, stem pitting of grapefruit and stem pitting of sweet orange (Garnsey et al., 1987; Roistacher and Moreno, 1991; Ballaster-Olmos et al., 1993).
Diagnosis of CTV strains using of indicator plants takes a long time because several different hosts are required and plant reaction and symptoms depend on several factors such as environmental conditions, plant varieties and virus strains. Serology is a widely used diagnostic method for CTV because these tests are simple, sensitive and inexpensive, however, when a plant is infected with more than one virus, apart from the target virus, other viruses may not be detected with serological methods. Otherwise, serological methods have not been able to identified differences between strains, including monoclonal antibodies with react several CTV isolates assayed worldwide (Valverde et al., 1990; Garnsey et al., 1993; Ochoa et al., 2000).
A method for isolation and analysis of double-stranded RNA (dsRNA) from virus infected plants tissues were developed by Morris and Dodds (1979). This method has been modified and is being used in several plant disease laboratories as a primary screening and diagnosis technique for viruses such as tobacco mosaic virus, cucumber mosaic virus and Citrus tristeza virus (Morris et al., 1983; Valverde et al., 1990; Moreno et al., 1993). DsRNAs which are produced in plants infected with RNA viruses are detected by using this technique. Healthy plants do not normally produce dsRNAs, however, some plants containing criptic viruses may yield similar size of dsRNAs to those related with single stranded RNA (ssRNA) viruses. This technique is not specific and can be used to distinguish not only different viruses but also strains of the same viruses as well as CTV strains (Dodds, 1986; Valverde et al., 1990; Moreno et al., 1993; Korkmaz, 2002).
DsRNA analysis provided the first direct evidence of separation of strains in CTV isolates. Subisolates obtained by inoculum propagation from a infected plant differed in their dsRNA patterns and also in their pathogenic behaviour (Moreno et al., 1991, 1993). Passage through different host species also produced modifications in their biological characteristics and/or dsRNA patterns (Moreno et al., 1993).
The aim of this study was to investigate biological properties and dsRNA patterns of nine isolates of Citrus tristeza virus collected from Satsuma plants Aegean region of Turkey. In addition, CTV antigen concentration and effect of tissue sampling time taken from naturally affected Satsuma Owari plant grown in the field of Balikesir province were also determined.
MATERIALS AND METHODS
CTV isolates and host plants: Nine CTV isolates, representative of citrus growing area of Aegean region of Turkey, were selected and determined double-stranded RNA (dsRNA) profiles and biological properties. CTV isolates, named KE-1, KE-2, KE-3, KE-4, KE-5, KE-6, KE-7, KE-8 and KE-9, were found in survey works in Aegean region of Turkey and kept at the Subtropical Fruit Research and Experimental Centre of Cukurova University. These isolates were maintained in Madam Vinous on Troyer citrange rootstock and kept in temperature controlled greenhouse conditions. Biological properties of these isolates was accomplished by graft inoculation with buds or leaf pieces from CTV infected donor plants onto a set of indicator plants as following; Citrus aurantifolia (Christm.) Swing. (Mexican lime), C. aurantium L. (sour orange), C. paradisi L. Rio red (grapefruit) and C. cinensis (L.) Obs., Madam Vinous (sweet orange). Inoculated plants were grown for 6-9 months in a temperature-controlled greenhouse at 18-26°C.
KE-5 CTV isolate was bud graft inoculated onto four seedlings of C. cinensis (L.) Obs., Madam Vinous (sweet orange), C. aurantium L. (sour orange), C. aurantifolia (Christm.) Swing. (Mexican lime), C. paradisi Rio red (grapefruit) plant to evaluate the host effect on dsRNA patterns. All inoculated plants were kept in the same greenhouse condition.
Titration of virus antigen: Bark tissues were collected from a 30 years old Satsuma Owari plant named EK-2 CTV isolate that found previous work at monthly intervals during one year period (2006). These samples were used for ELISA and dsRNA analysis. For ELISA test, peeled 0.5 g bark tissue was ground to a powder with mortar and pestle in 10 mL of phosphate-buffer saline, pH 7.4, containing 0.05% Tween 20, 2.0% polyvinylpyrrolidone (MW 40,000) and 0.2% bovine serum albumin. ELISA was carried out by a double antibody sandwich direct procedure as previously described (Garnsey and Cambra, 1991). The polyclonal coating antibody and IgG conjugated to alkaline-phosphates were provided from commercial company (Loewe-Germany) and were used according to the manufactory’s instruction. Substrate (p-nitrophenol phosphate) was added into ELISA plate and incubated for 30 min at room temperature. Absorbance at 405 nm was measured on a microtiter plate reader (Stat Fax-2100, Awareness Tech. Ins. (USA)).
Extraction and purification of dsRNA: A modified procedure based on that described by Valverde et al. (1990) was used. Barks of young shoots were used for the analysis, however, when bark tissues were not sufficiently enough, midribs of the leave were used. Young shoots of plants from inoculated with same isolate of CTV were taken, leaves and thorns were cut off and barks were peeled, immediately handled or frozen with liquid nitrogen and stored -20°C for later use. The same procedure was also used for naturally infected field grown EK-2 isolate of CTV.
Double-stranded RNA in STE (0.1 M NaCl, 0.05 M Tris-HCl, 1.0 mM EDTA, pH 6.8), were purified from phenol/chloroform/isoamylalcohol (25:24:1)-treated, buffered extracts from 2 to 6 g of bark tissue by two cycles of fractionation on columns of Whatman CF-11 cellulose in the presence of 16% ethanol. DsRNAs eluted from cellulose in ethanol free STE were concentrated by precipitation with two volumes of cold 95% ethanol and 0.1 volume of 3.0 M sodium acetate, pH 5.5 and resuspended in 30 μL TE (10 mM tris, 1.0 mM EDTA) buffer.
Electrophoresis was performed in 1% agarose or 5% polyacrylamide gel in electrophoresis buffer (40 mM tris, 20 mM sodium acetate, 1 mM EDTA, pH 7.8) for either 3 h at 70 volt (agarose gel) or 5 h at 100 volt (polyacrylamide gel). Gels were stained in 20 ng mL-1 ethidium bromides, visualized on an UV transilluminator and photographed with Polaroid type 667 black and white films. In addition silver staining was, if required, carried out on the same gel (Iglio, 1983). Molecular weights of CTV dsRNA were estimated by the graphical method of Bozarth and Harley (1976), in comparison with previously used standards (Dodds and Bar-Joseph, 1983; Valverde et al., 1986).
RESULTS
Nine isolates graft inoculated to a set of indicator plants did not show any symptoms on sour orange, grapefruit and sweet orange plants or did not cause any stunting when compared with healthy control. However, all isolates showed vein clearing symptoms on Mexican lime (Table 1). In addition, four isolates (KE-1, KE-2 and KE-8) induced severe leaf cupping and chlorosis symptoms and three isolates (KE-3, KE-4 and KE-7) showed chlorosis symptoms. On the other hand, non of the CTV isolates evaluated in this study caused stem pitting on Mexican lime within the period of 6-9 months after graft inoculation.
DsRNA patterns of CVT isolates: The dsRNA profiles of these isolates showed one major virus specific dsRNA band of 13.3x106 Da MW which is corresponded to the full length of replicative form of CTV genomic ssRNA (MW = 5.4-6.5x106) (Dodds and Bar-Joseph, 1983).
| Table 1: | Biological indexing results for nine CTV isolates used for dsRNA analysis |
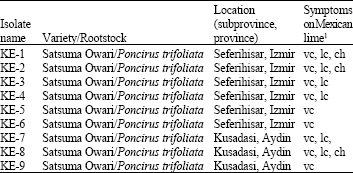 | |
| 1vc: vein clearing, ch: chlorosis, lc: leaf cupping | |
However, healthy Madam Vinous sweet orange plant did not show any dsRNA band onto gel. Agarose or polyacrylamide gel electrophoresis of dsRNA aliquots equivalent to one-tenth of extracted dsRNA normally resulted in detectable dsRNA band from 2 to 6 g bark tissue. Another three major dsRNAs having of 0.5, 0.8 and 2.0x106 MW were detected for all nine isolates. Additionally, most of isolates also showed some detectable major dsRNA bands having MW of greater than 2.0x106 Da. DsRNA analysis was several times done for nine CTV isolate and dsRNAs were detected throughout the year in the Madam Vinous sweet orange plants kept in a temperature controlled greenhouse (Fig. 1).
Influence of host on dsRNA pattern: One major virus specific dsRNA band of 13.3x106 Da MW was clearly obtained for all four citrus species. The additional two dsRNA of 2.0 and 0.8x106 MW were also detected as clear as full-length of dsRNA in four hosts, but recovery of these bands was less in sour orange. The 0.5x106 MW of dsRNA was detected in three hosts but sour orange did not have this band. Therefore, the results indicate that Madam Vinous and Mexican lime were the best hosts for dsRNA analysis. Rio-red grapefruit was good and sour orange was poor (Fig. 2).
Titration of virus antigen and dsRNA analysis of field sample: The highest ELISA value was obtained in March and April and then steadily decreased until September and then again gradually increased through December. ELISA value was the lowest in September even absorbance value fell below cutoff point of 2.5 times of the healthy control. These results show that spring is the best time to diagnosis of CTV in the field conditions (Table 2).
DsRNA recovery was good in March, April, May and June in naturally infected isolate KE-2. A major full-length of dsRNA band 13.3x106 MW and other some major or minor dsRNA bands (2.0 and 0.8x106 MW) were detected as clear as the same CTV isolate which was greenhouse inoculated.
| Table 2: | Detection of CTV from bark samples collected from a naturally infected Satsuma tree at monthly intervals for period of 1 year (2006) by ELISA and dsRNA |
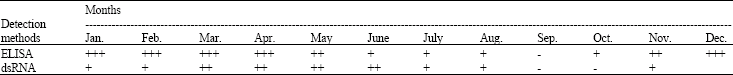 | |
| ++++: Very good, ++: Good, +: Poor, -: No reaction | |
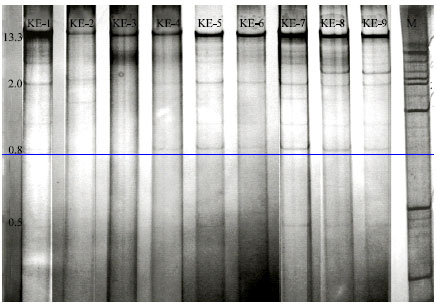 | |
| Fig. 1: | Polyacrylamide gel electrophoresis of dsRNA from greenhouse grown Madam Vinous sweet orange plants with infected nine Citrus tristeza virus isolates. Numbers on the left indicate molecular weights (x106 Da), as determined from the standards. M: Molecular weight standard Tobacco mosaic virus and Cucumber mosaic virus |
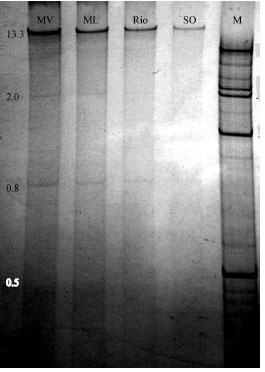 | |
| Fig. 2: | Host-induced variations in dsRNA recovery and dsRNA pattern of Citrus tristeza virus isolate of KE-5. Numbers on the right indicate molecular weights (x106 Da), as determined from the standards. Electrophoretic profiles: MV (Madam Vinous sweet orange); ML (Mexican lime); Rio (Rio Red grapefruit) and SO (sour orange) |
DsRNA pattern was poor in January, February, July, August, November and December. No dsRNA band was detected in September and October even in polyacrylamide gel (data not shown). For evaluation of dsRNA results, KE-2 isolate of CTV that maintained in Madam Vinous sweet orange plant in greenhouse was compared with the field sample as a control.
DISCUSSION
Nine Agean region of Turkish isolates of CTV showed only vein clearing symptoms in Mexican lime and none of those isolates showed either seedling yellows or stem pitting symptoms in inoculated plants. These results indicate that nine Aegean region Turkish isolates of CTV are mild strain did not cause stunting or stem pitting symptoms in field conditions in Turkey.
When the biological properties of three Turkish and one Cyprus isolates of CTV were investigated in a previous study it was found that Turkish isolates showed symptoms on only Mexican lime suggesting that they were all mild isolates of CTV (Korkmaz, 2002). This study shows that biological properties of Aegean region of Turkish CTV isolates has not changed and severe isolates are still absent at least in Aegean citrus regions of Turkey. On the other hand, the same study demonstrated that one CTV isolate from Guzelyurt (Cyprus) showed symptoms on grapefruit, Madam Vinous and Mexican lime (Korkmaz, 2002). Kyriakou et al. (1993) found several CTV infected grapefruit trees exhibited evident decline symptoms in the field in Cyprus revealing that severe isolates of CTV are present in Cyprus. Considering the close proximity of Cyprus to Turkey as well as the use of sour orange as the primary rootstock and the high abundance of the vector Aphis gossypii in Turkey suggest that CTV always remains as a threat to the Turkish citrus industry (Cinar et al. 1993). Therefore, periodic monitoring and biological indexing of CTV isolates in Turkey is necessary.
DsRNAs were detected from plants infected with all of nine isolates of CTV including full-length replicative form and several major or minor additional bands. Detectable dsRNA bands were obtained from one-tenth of extracted sample from 2 to 6 g CTV infected bark tissue. Dodds et al. (1987) reported successful detection of dsRNA band was purified from 1 to 2 g tissue. Present results is in agreement with those results, however, Moreno et al. (1990) suggested a minimum of 5 g tissue was necessary to detect the complete dsRNA pattern of some Spanish isolates. Therefore, amount of tissue used for analyses shows a variation depending upon isolate and environmental conditions.
A minor dsRNA band of 0.5x106 MV was detected in all of CTV isolates assayed. Results from previous studies suggest that this dsRNA pattern of CTV isolates was associated with severe strains causing the seedling yellows or stem pitting reaction on grapefruit (Dodds et al., 1987), but Moreno et al. (1990) observed this band by several moderate or mild strain Spanish isolates that no cause stem pitting on grapefruit. Albertini et al. (1988) also observed this band in their CTV strain K that does not cause symptoms even on Mexican lime plant. Results of these both investigations showed similarity to our findings.
Choice of a satisfactory host is important to readily determinate dsRNA patterns. Sweet orange and Mexican lime was found to be the best hosts. Grapefruit was also good host but, sour orange was the poor host. Moreno et al. (1990) also indicated sour orange was the poorest host for dsRNA.
Virus titer from individual sweet orange plant grown in the field was the highest in March and April and the lowest in September. This was an expected results that previously reported by various researches (Dodds et al., 1987; Garnsey et al., 1987; Mathews et al., 1997; Korkmaz, 2002). DsRNA analysis was not as good as ELISA results in the field conditions. Even, in polyacrylamide gel, dsRNA was not detected in September and October. In our conditions, it can be expressed that dsRNA analysis is not able to use as a diagnosis method for CTV in lately summer and early autumn season from field samples as Valverde et al. (1990) also reported, sometimes, field collected samples yield low amount of dsRNA and greenhouse inoculations are needed to improve dsRNA yield.
As a conclusion, this study gives some clues for the further investigation such as dsRNA profiles, host effect dsRNA pattern and diagnosis of CTV from field grown trees.
ACKNOWLEDGMENT
This study was supported by a Grant No. 104O129 from the Scientific and Technological Research Council of Turkey (TUBITAK).
REFERENCES
- Albertini, D., R. Vogel, C. Bove and J.M. Bove, 1988. Transmission and preliminary characterization of Citrus tristeza virus strain K. Proceedings of the 10th Conference of the International Organization of Citrus Virologists, November 17-21, 1986, Riverside, CA., pp: 17-21.
Direct Link - Hilf, M.E., A.V. Karasev, H.R. Pappu, D.J. Gumpf, C.L. Niblett and S.M. Garnsey, 1995. Characterization of Citrus tristeza virus subgenomic RNAs in infected tissue. Virology, 208: 576-582.
CrossRefDirect Link - Karasev, A.V., V.P. Boyko, S. Gowda, O.V. Nikolaeva and M.E. Hilf et al., 1995. Complete sequence of the Citrus tristeza virus RNA genome. Virology, 208: 511-520.
CrossRefDirect Link - Korkmaz, S., 2002. Investigation of biological properties, double-stranded RNA patterns and antigen concentration in citrus species infected with citrus tristeza virus, Phytoparasitica, 30: 420-428.
CrossRefDirect Link - Mathews, D.M., K. Riley and J.A. Dodds, 1997. Comparison of detection methods for Citrus tristeza virusin field trees during months on nonoptimal titer. Plant Dis., 81: 525-529.
Direct Link - Mawass, M., A.V. Karasev, E. Mietkiewska, R. Gafny, R.F. Lee, W.O. Dawson and M. Bar-Joseph, 1995. Defective RNA molecules associated with Citrus tristeza virus. Virology, 208: 383-387.
CrossRefDirect Link - Mawassi, M., E. Mietkiewska, R. Gofman, G. Yang and M. Bar-Joseph, 1996. Unusual sequence relationships between two isolates of Citrus tristeza. J. Gen. Vir., 77: 2359-2364.
CrossRefDirect Link - Rocha-Pena, M.A., R.F. Lee, R. Lastra, C.L. Niblett, F.M. Ochoa-Corona, S.M. Garnsey and R.K. Yokomi, 1995. Citrus tristeza virus and its aphid vector Toxoptera citmicida: Threats to citrus production in the Caribbean and central and North America. Plant Dis., 79: 437-445.
CrossRefDirect Link