
B.J. Neya
Institut de l�Environnement et de Recherches Agricoles (INERA), 01 BP 476 Ouagadougou 01, Burkina Faso
J. Zabre
Institut de l�Environnement et de Recherches Agricoles (INERA), 01 BP 476 Ouagadougou 01, Burkina Faso
R.J. Millogo
UFR/SVT, Departement de Biologie et Ecologie Vegetales, Universite de Ouagadougou, 03 BP 7021 Ouagadougou 03, Burkina Faso
S. Guinko
UFR/SVT, Departement de Biologie et Ecologie Vegetales, Universite de Ouagadougou, 03 BP 7021 Ouagadougou 03, Burkina Faso
G. Konate
Institut de l�Environnement et de Recherches Agricoles (INERA), 01 BP 476 Ouagadougou 01, Burkina Faso
Plant Pathology Journal
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 75-84







ABSTRACT
The effect of contamination of cowpea seeds by Cowpea aphid borne mosaic virus (CABMV) on disease development was studied in three agro-ecological zones of Burkina Faso. Seed contamination is the only source of primary infection by CABMV in the field while aphid vectors are responsible for secondary infections. Two cowpea varieties with different seed contamination rate (0, 0.05, 0.25, 0.5, 1 and 5%) were used. The infected plants were recorded every week from the tenth Day After Sowing (DAS) and over seven weeks. In the same way, populations of aphids were evaluated in plots at 30 DAS. There was a difference between the average numbers of insects from one location to another. The initial contamination rate of cowpea seeds by the CABMV played a leading important role in epidemics development according to the areas. With the variety Local Gorom, the secondary infections were early and fast in Niangoloko zone. In Sapouy area, the secondary infections were lower and belated. In the centre zone the impacts were lower than those of Sapouy. In the case of variety Kvx61-1, the secondary infections were medium for all initial contamination rates even in years of great epidemics. For each year, any difference of impact rate had not got between the different seeds upset contamination levels Kvx61-1 behaved well in the three localities even the west where aphids appear precociously and densely.
PDF Abstract XML References Citation
How to cite this article
B.J. Neya, J. Zabre, R.J. Millogo, S. Guinko and G. Konate, 2008. Propagation of the CABMV from Infected Seeds in Three Zones of Burkina. Plant Pathology Journal, 7: 75-84.
DOI: 10.3923/ppj.2008.75.84
URL: https://scialert.net/abstract/?doi=ppj.2008.75.84
DOI: 10.3923/ppj.2008.75.84
URL: https://scialert.net/abstract/?doi=ppj.2008.75.84
INTRODUCTION
The world cowpea production was estimated at three millions tons. Up to 12.5 millions hectares are under cowpea cultivation, 64% of are located in Africa (Singh et al., 1997). An increased and sustained production has been recorded during the last five years in Malawi, Niger, Nigeria, Burkina Faso, Mali, Togo and Benin. In each part of the world and especially in Africa, cowpea production extension is emphasized.
Unfortunately, the increase of cowpea production is jeopardized by several biotic and abiotic constraints that limit or even annihilate cowpea yields. The point is the non availability of adapted varieties to the different zones, soil and climate combined action, harmful insects and weeds pressure, finally and especially diseases (Taiwo and Akinjogunla, 2006; Kareem and Taiwo, 2007). In the case of aphid-transmitted viruses, scabs or brown stains, susceptible cowpea varieties can suffer from yield losses between 50 and 100% in savannah zones (Reeves, 1983).
Very little monitored until a recent past on statistical grounds, the national production in Burkina Faso increased these last five years from 309,466 t in 1999 to around 456,000 ton in 2003 (DGPSA, 2004). Climatic conditions in the country is characterised by strong contrasts going from the North to the South, which imposes the use of adapted cowpea varieties and auspicious sowing dates to every zone. Burkina Faso is part of the Sahel region which stretches from Senegal to Sudan and is enclosed by 7° and 15° North latitude and 17° West and 24° East longitude. Three agro-ecological zones are distinguished (Anonymous, 1993):
| • | The Sahelian zone in the North with an annual rainfall of 400 to 600 mm |
| • | The Sudanian zone in the centre (600 to 900 mm of annual rainfall) |
| • | North Guinean zone (900 to 1200 mm) in the South West |
All agro-ecological zones are well suitable to cowpea cultivation because several varieties adapted to the different climatic conditions of the country have been developed by research institutions. However, most cowpea production is done in the centre and western part of the country. Although CABMV has been reported in all agro-ecological zones two decades ago (Somé, 1989), the effect of climatic conditions on the development of the cowpea mosaic disease has not been investigated.
The objective of this study is to assess and to compare the impact of the agro-ecological differences on cowpea mosaic disease development in Burkina Faso. This information is necessary for the development of adequate control measures against CABMV. Results obtained in this country may also be useful for other countries of the Sahel region, since they all shared similar agro-ecological conditions.
MATERIALS AND METHODS
Plant material and seed contamination rates: Two cowpea varieties were used throughout the experiments: Local Gorom representing the group of the susceptible varieties (Tignegre, 2000; Neya, 2002) with four levels of seed contamination rate (0, 0.05, 0.25 and 0.5%) and Kvx61-1 the one of the tolerant or resistant varieties (Tignegre, 2000; Neya, 2002) with four levels of seed contamination rate (0, 0.25, 1 and 5%).
All seed contamination levels were obtained by mixing contaminated seeds with virus-free seeds at appropriate proportions. Contaminated seeds were identified by means of the non-destructive Enzyme-linked immunosorbent assay (ELISA) as implemented by Konate and Neya (1996). Basically, the test was conducted as described by Clark and Adams (1977), especially with regard to buffer systems and incubation times. A fragment of 60 mg was taken from the distal part (opposite to the embryo side) of each seed and hand-ground with mortars and pestles in 150 μL extraction buffer. After decantation, the supernatant was used as antigen source. Absorbance readings (A405 nm) were recorded with a Metertech Σ960 automatic ELISA plate reader. Seed extracts were considered to be positive for CABMV detection if their A405 nm readings were higher than thrice those of healthy control seeds. The remaining embryo-side parts of the positive seeds of each cowpea variety were pooled and used as virus-contaminated seed batches.
Field experiments and statistical analyses: Cowpea mosaic disease development was investigated over three consecutive years (from 2002 to 2004) in the central, the west-central and the western parts of Burkina Faso. Specific locations in each part were Somnawaye (Centre), Sapouy (West-central) and Niangoloko (West). In each location, experiments were conducted on 200 m2-plots separated one from another by at least 50 m to minimize cross infections between plots. Treatments were composed of the different seed contamination rates and three replicates were done per treatment. In each plot, mineral fertiliser consisting of Nitrogen-Phosphate-Potassium (NPK 14-23-14) was applied at 100 kg ha-1. Cowpea seeds were sown in rows with distances of 0.80 m between rows and 0.40 m between seed holes on the same row.
Proportions of infected plants were recorded in each plot once a week from 10 Days After Sowing (DAS) to 59 DAS. Populations of aphid vectors were evaluated at 30 DAS using the six-digits (0 to 5) evaluation scale of Jackai and Singh (1988) where 0 = no infestation; 1 = 1-4 aphids per plant; 2 = 5-20 aphids per plant; 3 = 21-100 aphids per plant; 4 = 101-500 aphids per plant and 5 = more than 500 aphids per plant. All disease incidence and aphid counting data were analyzed by analysis of variance (ANOVA) (Stanton, 1996).
RESULTS
Counting of the total number of plants at germination: One week after sowing, the total number of the plants of each variety was been counted on all plots in the three zones. At the plots level, the number of plants varied between a minimum of 614 and a maximum of 737 (Table 1a, b).
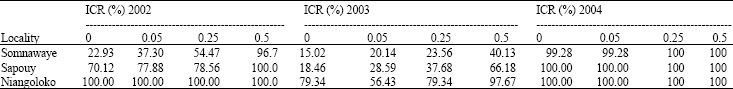
Assessment of aphid populations 30 days after sowing in the three locations: The number of aphids in the plots corresponding to each seeds initial contamination rate was apprised. The results were consigned in the Table 2a and b. The notes assigned in each of the two groups of tested cowpea varieties according to the entomologists scale varied between 2 and 4. For each year, the analysis of variance showed that there was no significant difference between the average number of insects per plot, per variety and per location. In 2002, for the Local Gorom (F = 1.58; p = 0.27) whereas for the variety Kvx61-1 (F = 0.33; p = 0.80). In 2003, these values were (F = 0.25 p = 0.86) for the Local Gorom and (F = 0.67; p = 0.59) for Kvx61-1. In 2004, for Local Gorom, F = 0.33; p = 0.80 while for Kvx61-1 (F = 0.25 and p = 0.86). Even though according to Newman-Keuls’ test, the analysis of variance didn't shown any significant difference of aphids infestation in the plots, but the note four was very often been assigned in the plots of the locality of Niangoloko confirming aphids steeper pressure in this zone.
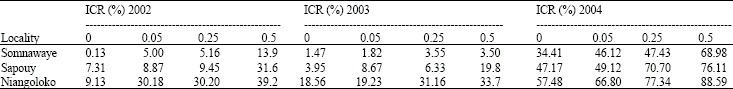
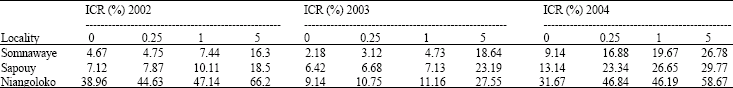
The effect of seeds contamination rate on the disease impact 31 and 59 DAS: In 2002, 31 DAS, the disease impact was as well superior in the locality of Niangoloko for each initial contamination rate with Local Gorom as with Kvx61-1 followed by Sapouy locality that had an intermediate impact.
| Table 1a: | Mean total number of plants per Initial Contamination Rate (ICR) and per zone with Local Gorom |
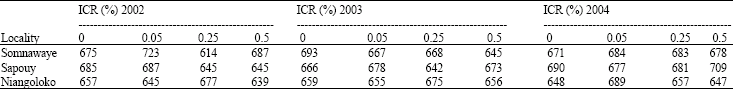 | |
| Table 1b: | Mean total number of plants per Initial Contamination Rate (ICR) and per zone with Kvx61-1 |
 | |
| ICR = Initial Contamination Rate | |
| Table 2a: | Aphid’s population assessment on the plots one month after sowing with Gorom local |
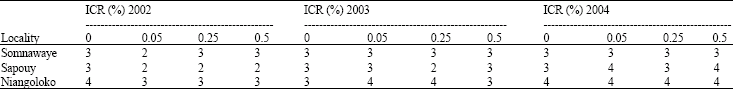 | |
| Table 2b: | Aphid’s population assessment on the plots one month after sowing with Kvx61-1 |
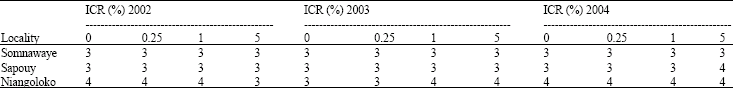 | |
| ICR = Initial Contamination Rate, Aphids number have bean appraised using the following scale, 0 = no infestation; 1 = 1-4 aphids per plant; 2 = 5-20 aphids per plant; 3 = 21-100 aphids per plant; 4 = 101-500 aphids per plant; 5 = more than 500 aphids per plant | |
The locality of Somnawaye had the weakest some impact both the variety and the initial contamination rate. With the variety Local Gorom, the analysis of variance didn't shown a significant difference between the various seeds initial contamination rates (F = 1.89; p = 0.21) (Table 3a), on the other hand, with the variety Kvx61-1, there is a significant difference between the contamination rates of 1 and 5% and the rates of zero and 0.25% (F = 4.81; p = 0.034), (Table 3b).
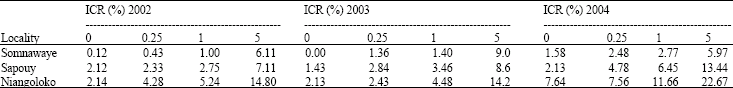
All as at 31 DAS, the disease impact rates at 59 days after sowing reached 100% with the variety Local Gorom either the upset contamination rate in Niangoloko. In Sapouy, the rate of sick plants also reached 100% in the plots where seeds were initially contaminated at 0.5% against 70 and 78% in the other plots. There again, there was no significant difference between the various initial rates but there is a significant difference between Niangoloko zone and the one of Somnawaye (F = 5.72; p = 0.025) (Table 4a). With the Kvx61-1 variety, the infection rates were lower in comparison with Local Gorom. For all initial contamination rates, the zone of Niangoloko had the higher impact (39 to 66%) followed by the one of Sapouy (7 to 18%). Low impacts have been recorded in Somnawaye (5 to 16%) from the lower initial contamination rate to the higher. The analysis of variance showed a significant difference between the zone of Niangoloko and the two others (F = 32.12; p = 0.001), (Table 4b).
For the year 2003, the different seeds initial contamination rates of Local Gorom gave at 31 DAS impact rates similar to the previous year. Niangoloko had the higher impact always followed by Sapouy and before Somnawaye. In each locality, the impacts of the mosaic virus changed in the increasing sense of the upset contamination rates. Therefore, a significant difference between the Niangoloko zone and the two others have been shown (F = 14.89; p = 0.001) (Table 3a). With the variety Kvx61-1 at this date, the number of sick plants was low in the three localities. The analysis of variance didn't shown a significant difference between the three zones (F = 0.46; p = 0.666). On the other hand, the impact corresponding to the contamination rate of 5% was significant different from those corresponding to the other rates (F = 15.78; p = 0.001) (Table 4b).
The impact rates of the disease with Gorom local 59 DAS were less elevated than those of the year before. In Niangoloko, this rate was respectively of 48.5% from virus-free seeds against 18.5 and 15% in Sapouy and in Somnawaye.
| Table 3a: | Disease impact 31 DAS from the different ICR with Local Gorom in the three localities |
 | |
| Table 3b: | Disease impact 31 DAS from the different ICR with Kvx61-1 in the three localities |
 | |
| ICR = Initial Contamination Rate | |
| Table 4a: | Disease impact 59 DAS from the different ICR with Local Gorom in the three localities |
 | |
| Table 4b: | Disease impact 59 DAS from the different ICR with Kvx61-1 in the three localities |
 | |
| ICR = Initial Contamination Rate | |
In each zone, the high impacts have been got with seeds with initial contamination rates of 5%. Niangoloko was in head with 97.67% followed by Sapouy with 66.18% and finally Somnawaye with 40.13% of impact rate. There wasn't a significant difference between the different contamination rates (F = 1.59; p = 0.267). On the other hand it has been between Niangoloko and the two other localities (F = 6.43; p = 0.018), (Table 4a).
At the same date, with the variety Kvx61-1, impacts were very low in the three zones and for all seeds initial contamination rates. The analysis of variance showed that there didn't have a significant difference between the three sites. But, the impact corresponding impact to the contamination rate of 5% was significantly different from those corresponding to the other rates (F = 14.05; p = 0.001) (Table 4b).
Finally in 2004, the disease impacts with Gorom local 31 DAS were upper than 50% in Niangoloko even with virus-free seeds. The analysis of variance didn't shown a meaningful difference as well between the three zones (F = 2.69; p = 0.121) as between the different contamination rates (F = 3.75; p = 0.060), (Table 3a). At this date, the impact rates were very low in the three zones with the variety Kvx61-1. With seeds with initial contamination of 5%, the impact was from 22.67% in Niangoloko, 13.44% in Sapouy and 6% in Somnawaye. No significant difference has been shown neither between the localities (F = 3.32; p = 0.083) nor between the different initial contamination rates (F = 2.35; p = 0.149), (Table 3b).
With Local Gorom, the impact rates reached 100% in all localities 59 DAS even in the plots where virus free seeds have been used. The analysis of variance showed that there is no significant difference between localities or between the different initial contamination rates, Table 4a. With the variety Kvx61-1, the impacts were at this date, lower than 50% for all initial contamination rates but the initial contamination rate of 5% in Niangoloko with an impact rate of 58.67%. There was no significant difference between the different initial contamination rates but in the other hand it had been between Niangoloko and the other localities (F = 8.68; p = 0.008), (Table 4b).
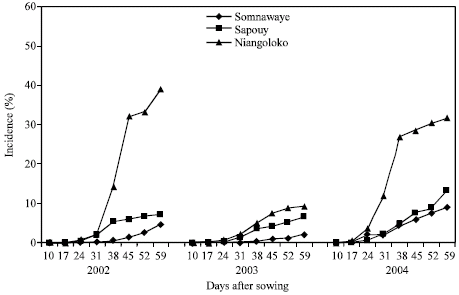
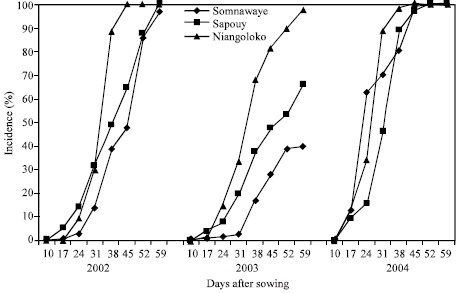
The kinetics of the CABMV in-field spreading from a virus free seeds (0%) from 2002 to 2004 with the two varieties in the three zones: The evolution of the number of sick plants in the plots sowed with virus-free seeds during the three years differs from one year to the next with however some likeness. With the variety Local Gorom representing the group of the varieties with high power of virus transmission by seeds, the obtained curves in Niangoloko zone are all of sigmoid pace. It indicates a very fast propagation speed permitting to reach in 2002 and in 2004 infection rates of 100% between 17 and 38 DAS.
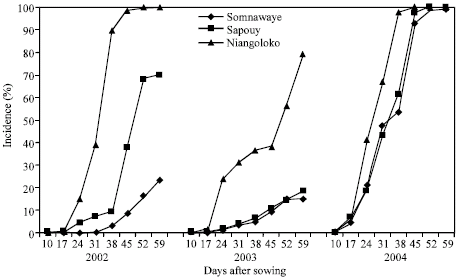 | |
| Fig. 1: | Kinetic of the CABMV propagation from an initial contamination rate of 0% with Gorom local in the three localities in 2002, 2003 and 2004 |
 | |
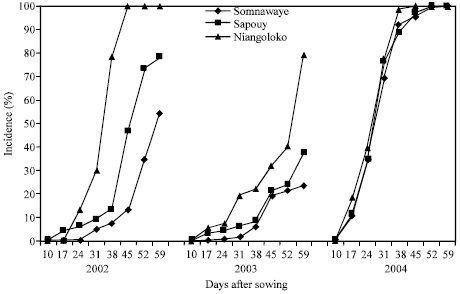
| Fig. 2: | Kinetic of the CABMV propagation from an initial contamination rate of 0% with Kvx61-1 in the three localities in 2002, 2003 and 2004 |
The disease impact remained lower than 80% in 2003, year of low epidemic (Fig. 1). With the variety Kvx61-1, the secondary infections began practically 31 DAS giving thus a low impact rate in 2002 (39%) and in 2004 (32%). In 2003, the spreading curve kinetics showed that the epidemic development speed was very low giving an impact rate of 9% at the end of the infected plants counting (Fig. 2).
In Sapouy, with the variety Local Gorom, the obtained spreading kinetics curves showed a low plants infection speed in 2002 until 38 DAS. After this date, the propagation speed increased to reach an impact rate of 70% at 59 DAS. In 2003, the disease propagation speed was very low. The impact rate was lower than 20% at the end of the experience. On the other hand in 2004, the propagation curve kinetics gave a sigmoid pace indicating a fast speed of the epidemic development. Indeed, started 17 DAS, the secondary infections reached an impact rate of 100% 45 DAS (Fig. 1). With the variety Kvx61-1, the propagation speed was very low in 2002, in 2003 and in 2004 with an impact of 13% in spite of that was a year of virus high epidemic (Fig. 2).
In the locality of Somnawaye, with Local Gorom, the disease propagation speed was very low in 2002 and in 2003 giving respectively impact rates of 23 and 15%. On the other hand, this speed was high and had generated an impact rate of 100% at 52 DAS in 2004 (Fig. 1).
 | |
| Fig. 3: | Kinetic of the CABMV propagation from an initial contamination rate of 0. 25% with Gorom local in the three localities in 2002, 2003 and 2004 |
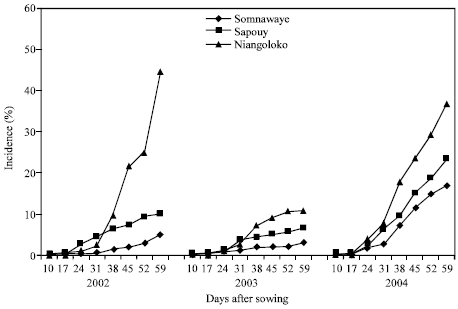 | |
| Fig. 4: | Kinetic of the CABMV propagation from an initial contamination rate of 0. 25% Kvx61-1 local in the three localities in 2002, 2003 and 2004 |
With the variety of low virus transmission power by seeds, the obtained curves showed that the disease propagation speed was even lower over the three years compared to those of Sapouy (Fig. 2).
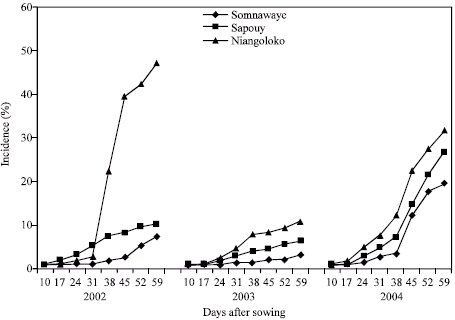
The kinetics of the CABMV propagation in-field from an initial contamination rate of 0.25% from 2002 to 2004 with the two varieties: From an initial contamination rate of 0.25%, the evolution of sick plants number in the plots during the three years differs not only from one year to the next but also from one variety to the other.
The obtained curves got in Niangoloko zone with the variety Local Gorom representing the group of the varieties with high powers of virus transmission by seeds, are all of sigmoid pace excepted the one of 2003. Started to 17 DAS, secondary infections had a very fast propagation speed allowing reaching in 2002 and in 2004 infection rates of 100% between 17 and 38 DAS. The disease impact remained lower than 50% in 2003, year of low epidemic (Fig. 3). With the variety Kvx61-1, secondary infections timidly began 31 DAS. This situation gave thus a mean impact rate of 45% in 2002 and in 2004. The propagation kinetics curve showed that in 2003 the epidemic development speed was very low with an impact rate of 11% (Fig. 4).
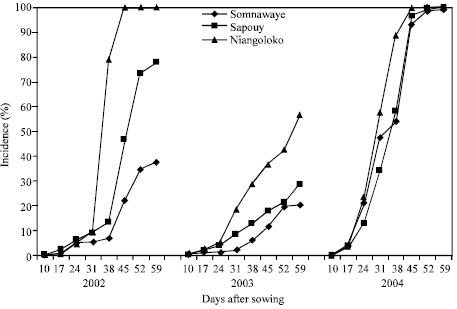 | |
| Fig. 5: | Kinetic of the CABMV propagation from an initial contamination rate of 0.05% with Gorom local in the three localities in 2002, 2003 and 2004 |
 | |
| Fig. 6: | Kinetic of the CABMV propagation from an initial contamination rate of 0.5% with Gorom local in the three localities in 2002, 2003 and 2004 |
With the variety Local Gorom in Sapouy, the propagation kinetics curves got showed a delayed infection starting until 36 DAS in 2002 and 2003. The propagation speed increased after this date to reach an impact rate of 78% in 2002 against 38% in 2003 59 DAS. The propagation kinetics curves gave on the other hand in 2004 a sigmoid pace indicating a fast speed of the epidemic development. Indeed, started 10 DAS, the secondary infections reached an impact rate of 100% 38 DAS (Fig. 3). With the variety Kvx61-1, the propagation speed was lower than 10% in 2002 and in 2003 and even in 2004 with an impact of 23.34% in spite of the fact that it is one year of virus high epidemic associated to a basis contamination rate of 0.25% (Fig. 4).
In the locality of Somnawaye, with Local Gorom, the disease of propagation speed was medium in 2002 and low in 2003 giving respectively impact rates of 55 and 24%. On the other hand, this speed was high and generated an impact rate of 100% 52 DAS (Fig. 3). With the variety Kvx61-1 with low power of virus transmission by seeds, the obtained curves showed that the disease propagation speed was even lower through the three years compared to those of Sapouy (Fig. 4).
The kinetics of the CABMV propagation in-field from an initial contamination rate of 0.05 and 0.5% of 2002 to 2004 with Local Gorom: The obtained curves in the zone of Niangoloko with the variety Local Gorom from the initial contamination rates of 0.05 and 0.5% are all of sigmoid paces and similar, excepted the one of 2003 with 0.05% of initial contamination rate.
 | |
| Fig. 7: | Kinetic of the CABMV propagation from an initial contamination rate of 1% with Kvx61-1 in the three localities in 2002, 2003 and 2004 |
The secondary infections had a very fast propagation speed and started between 10 and 17 DAS, they reached then infection rates of 100% 45 DAS. The disease impact remained lower than 100% in 2003 in the plots that received contaminated seeds at 0.05 and 0.5%. In spite of the fact that the second contamination rate is ten times superior to the first. The impacts rates were more or less the same (Fig. 5, 6).
The obtained kinetics of propagation curves of Sapouy locality with these two seeds contamination levels was different according to the basis contamination level. The impact rates from seeds contaminated at 0.05% have been raised in 2002 (78%), very low in 2003 (29%) and very elevated and precocious in 2004, 100% at 45 DAS (Fig. 5). With seeds contaminated at 0.5%, the disease propagation speed has been raised in 2002 and reached the impact rate of 100% 59 DAS against 66% in 2003. It has been very height and very precocious in 2004 and reached the maximum impact rate 45 DAS (Fig. 6).
In the locality of Somnawaye, from seeds contaminated at 0.05% with Local Gorom, the disease propagation speed was low in 2002, 37% 59 DAS. It was even lower in 2003 giving an impact rate of 20% at the same date. On the other hand, this speed was high and precocious in 2004 and generated an impact rate of 100% 45 DAS (Fig. 5). With an initial contamination rate of 0.5%, impacts of 100% have been obtained between 52 and 59 DAS in 2002 and in 2004 following a fast epidemic development contrary to the year 2003 where an impact of 40% has been obtained 59 DAS (Fig. 6).
The kinetics of the CABMV propagation in- field from an initial contamination rate of 1 and 5% from 2002 to 2004 with Kvx61-1: With the variety Kvx61-1 and from infected seeds at 1% by virus, the secondary infections timidly began 31 DAS in 2002 in Niangoloko. Then, the propagation speed became medium and gave an impact rate of 48% 59 DAS. This propagation speed and this impact rate were similar to those obtained in 2004. The curve of propagation kinetics had a shy progression in 2003 and the epidemic development speed was very low with an impact rate of 11% (Fig. 7).
With seeds contaminated at 5%, the features of epidemics development were identical to those gotten with infected seeds at 1%. Therefore, a high increase of impact rates has been observed (Fig. 8).
In Sapouy zone, the propagation speed was low and the impact rates were of 10% in 2002 and 7% in 2003 and even in 2004 with an impact of 27% in spite of the fact that it is a year of the high epidemic associated to a contamination rate 1% (Fig. 7).
From an initial rate of 5%, the impacts were of 19% in 2002, 23% in 2003 and 30% in 2004 (Fig. 8).
In Somnawaye, with Kvx61-1 seeds contaminated at 1%, the obtained curves showed that the disease propagation speed was lower during the three years compared to those of Sapouy and Niangoloko. The impact rates progressed between 5 and 20% (Fig. 7). With a basis rate of 5%, the disease propagation speed had the previous features (Fig. 8).
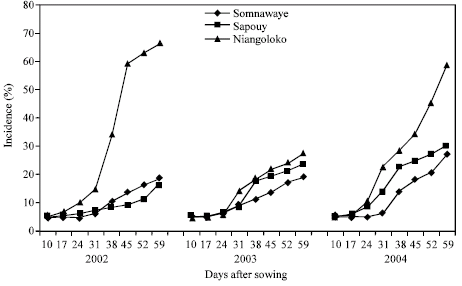 | |
| Fig. 8: | Kinetic of the CABMV propagation from an initial contamination rate of 5% with Kvx61-1 in the three localities in 2002, 2003 and 2004 |
DISCUSSION
To the term of this study, it came out again aphids infestation importance varies according to the seasons and is encouraged by drought pockets. These results confirm those of Dabire (1992). The results also showed that the development of the CABMV epidemic is faster and more precocious in the western area. With the varieties of high powers to transmit virus by seeds represented by the variety Local Gorom, the curves had a sigmoid pace whatever the initial contamination rate. The secondary infections have especially been observed from the seventeenth day after sowing. They also marked the beginning of the logarithmic phase that ends since the 45th DAS because most plants have been infected giving impact rates of 100%. There was no impact rate difference between the average of plots sowed with virus-free seeds and those sowed with seeds at a rate of 0.5%. It could be due to the effect of the secondary contaminations crossed assured by aphids (Frison, 1988) between plots corresponding to the various seeds initial contamination rates. Aphid’s high number and inoculum availability in the plots were undoubtedly at the origin of this result. According to Dabire (1992), it is indeed, evident from her works that aphid flights are more precocious and more intense in the west and decreases from the centre to the north. This situation would explain the decrease of the impact in the same sense that wants to say from West to North.
In the case of the varieties of low power of virus transmission by seeds represented by the variety Kvx61-1, the secondary infections have been observed between 31 and 38 DAS generating impact rats lower than 50% in Niangoloko.
In Sapouy locality, the diseases propagation is more belated and less fast than to the west with Local Gorom. The impact rate of 100% is reached with one or two weeks of delay on the west zone.
In the case of the varieties of low faculty of the CABMV transmission by seeds as the variety Kvx61-1, the secondary infections have been observed between 31 and 38 DAS. In any case, the disease progressed slowly with constant speed leading to impacts lower than 30% 59 DAS in spite of relatively a high seeds initial contamination rates.
Finally, in the centre zone (Somnawaye), Local Gorom gave some lower impact rates than those of Sapouy region. The impact rates of 100% have been reached in the last two weeks of the study in year of high epidemic as 2004 or some times with contaminated seeds at 0.5%. With Kvx61-1 variety, the CABMV propagation was low giving some low impact rates thus even with seeds with contamination rate of 5%. For this variety Kvx61-1, the disease progression curves in the three localities indicate a high resistance level to the infection. So, the use of seeds little or not contaminated can in this case being favourable to face the CABMV.
The high levels of infection especially observed with the variety Local Gorom representing the varieties of high faculty of virus transmission to seed translate the necessity to reduce aphids’ populations’ density notably by the use of insecticides during cowpea vegetative cycle. Aphids were responsible for the secondary contaminations, what bears out the results of Frison (1988). Insecticides application should be made in the third week after the sowing, most infections occurring between 17 and 24 days.
The obtaining of impact rates of 100% from virus-free seeds of virus with Gorom local settles then the problematic of these seeds use in some localities. The solution would be in the setting of an integrated biological where it is necessary to isolate the fields of inoculum external important sources and to apply a precocious treatment against aphids.
The study of epidemics development in the three zones from contaminated seeds or not showed that:
| • | The disease development is faster and higher in Niangoloko representative of the western area |
| • | This development is medium in Sapouy that represents the western centre |
| • | In Somnawaye that represents the centre zone, the disease impact is low |
| • | In case of year of high epidemic as the case of 2004, there is no significant difference of the disease with the variety Local Gorom between the virus-free seeds and those contaminated at 0.5% in Niangoloko and Sapouy zones |
| • | The variety Kvx61-1 representing the group of the resistant or tolerant varieties had a good behaviour in all zone even in 2004 |
| • | Aphids infestation is more intense in the west and decrease as one goes back up northwards. It would entail the development of the epidemics presumably in the same sense. More the pressure of the vector is high, more the epidemic is fast |
ACKNOWLEDGMENTS
My acknowledgments first of all go then to Dr. Oumar TRAORE for his very big contribution in the realization of this document, then to the project Seed Health Improvement Programme (SHIP) for his financial support, finally to all the team of this project to know lady Elisabeth ZOMA the national coordinator, Dr. Adama NEYA and Dr. Irénée SOMDA.
REFERENCES
- Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Kareem, K.T. and M.A. Taiwo, 2007. Interaction of viruses in cowpea: Effect on growth and yield parameters. Virol. J., 4: 15-15.
CrossRefPubMedDirect Link - Konate, G. and B.J. Neya, 1996. Rapid detection of cowpea aphid-borne mosaic virus in cowpea seeds. Ann. Applied Biol., 129: 261-266.
CrossRefDirect Link - Taiwo, M.A. and O.J. Akinjogunla, 2006. Cowpea viruses: Quantitative and qualitative effects of single and mixed viral infections. Afr. J. Biol., 5: 1749-1756.
Direct Link