
K.F. Chang
Field Crop Development Centre, Alberta Agriculture and Food (AAF), Lacombe, Alberta, T4L 1W8, Canada
S.F. Hwang
Crop Diversification Centre, North, AAF, Edmonton, Alberta, T5Y 6H3, Canada
A.H. Khadhair
Crop Diversification Centre, North, AAF, Edmonton, Alberta, T5Y 6H3, Canada
H.U. Ahmed
Field Crop Development Centre, Alberta Agriculture and Food (AAF), Lacombe, Alberta, T4L 1W8, Canada
S.E. Strelkov
Department of Agricultural, Food and Nutritional Science, University of Alberta, Edmonton, Alberta, T6G 2P5, Canada
M. Deyholos
Department of Biological Sciences, University of Alberta, Edmonton, Alberta, T6G 2E9, Canada
G.D. Turnbull
Crop Diversification Centre, North, AAF, Edmonton, Alberta, T5Y 6H3, Canada
J. Feng
Crop Diversification Centre, North, AAF, Edmonton, Alberta, T5Y 6H3, Canada
Plant Pathology Journal
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 20-26







ABSTRACT
During the summer of 2003 and 2004, a total of 58 isolates of Ascochyta rabiei were collected from chickpea plants grown in southern Alberta, Canada. RAPD analysis of genomic DNA extracted from these isolates was conducted using six short sequence primers (OPA-03, OPA-13, OPB-07, OPC-01, OPC-20 and OPJ-15) and analyzed to establish the genetic relationship and distance between isolates. A total of 75 bands were polymorphic. The 58 isolates were found to belong to five genotypes, indicating that the A. rabiei population of southern Alberta is genetically diverse. No relationship was found between the genotype groupings obtained through RAPD analysis and previously determined pathotype classifications of the same isolates.
PDF Abstract XML References Citation
How to cite this article
K.F. Chang, S.F. Hwang, A.H. Khadhair, H.U. Ahmed, S.E. Strelkov, M. Deyholos, G.D. Turnbull and J. Feng, 2008. Molecular Diversity of Ascochyta rabiei Isolates from Chickpea in Alberta, Canada. Plant Pathology Journal, 7: 20-26.
DOI: 10.3923/ppj.2008.20.26
URL: https://scialert.net/abstract/?doi=ppj.2008.20.26
DOI: 10.3923/ppj.2008.20.26
URL: https://scialert.net/abstract/?doi=ppj.2008.20.26
INTRODUCTION
Chickpea (Cicer arietinum L.) is one of the three most important pulse crops, ranked second to dry bean (Phaseolus vulgaris L.) and third in production among the legume crops. Its high ranking among the legume crops results from its high nutritional value and capacity to grow in arid and semi-arid lands (FAOSTAT, 2005). According to the Food and Agriculture Organization (FAO), the average yield of chickpea is very low, at 0.8 t ha-1. Both biotic and abiotic factors contribute to its low, unpredictable yield. Infection by Ascochyta rabiei (Pass.) Lab. syn. Didymella rabiei (Kovacevski) v. Arx is one of the primary challenges in chickpea yield improvement and the primary impediment to adoption of the chickpea crop by farmers worldwide. In western Canada, there was a great expansion in chickpea cultivation from 1995 to 2001. During this period, chickpea production increased 350-fold. However, since 2001, production has declined substantially (McVicar et al., 2006), partially because of the spread of ascochyta blight (Ahmed et al., 2005, 2006; Chang et al., 2000, 2007; Chongo and Gossen, 2003). It has been reported that severe epidemics of ascochyta blight have occurred in various chickpea producing regions, including those regions where ascochyta-resistant cultivars have been adopted (Navas-Cortés et al., 1998; Nene, 1982; Singh et al., 1984; Nene and Reddy, 1987).
The severity of symptom development in chickpea cultivars (Porta-Puglia et al., 1996), lines (Chen et al., 2004; Navas-Cortés et al., 1998) and accessions (Jamil et al., 2000) has been used to group pathogens causing ascochyta blight into pathotypes. In order to test the virulence of A. rabiei isolates, the reactions of accessions included in a differential set have been categorized as resistant, tolerant and susceptible to the pathogen. In Canada, 14 pathotypes of A. rabiei were identified among 40 isolates tested, based on the reactions of eight chickpea differentials (Chongo et al., 2004).
Molecular marker studies have been conducted on several isolates of A. rabiei. RAPD analysis was used to genotype A. rabiei isolates in most of these studies. This technique has been used to differentiate isolates and to reveal significant genetic diversity (Fischer et al., 1995). Two RAPD markers were selected for these fungal isolates in Pakistan (Jamil et al., 2000), five markers in Spain (Navas-Cortés et al., 1998) and ten markers in Syria (Udupa et al., 1998). However, there have been no molecular studies examining the molecular diversity of A. rabiei isolates from chickpea in Alberta. Therefore, this study was undertaken to (1) evaluate the genetic variability of these isolates using molecular markers and (2) determine the relationship between the genotypes and pathotypes of the A. rabiei isolates from Alberta.
MATERIALS AND METHODS
Fungal sources: During the summer of 2003 and 2004, samples of ascochyta-infected plants were collected from chickpea fields near the Carmangay, Coaldale, Foremost and Vulcan areas of southern Alberta, Canada. Small pieces of leaf, stem or pod lesions on which pycnidia were visible were surface sterilized in a 1% solution of commercial bleach for 30 sec to 1 min and plated onto a water agar medium (1.5% agar). The plates were incubated at room temperature until pycnidiospores oozed out (48 to 72 h after plating). A small amount of the pycnidiospore mass was transferred into 0.5 mL of sterile water and a 300 μL aliquot of spore suspension was transferred and spread onto chickpea-agar medium plates. The plates were incubated at room temperature for 48 h and examined under a stereomicroscope for germination of pycnidisiospores. Single spores were transferred to agar plates for subsequent use (Ahmed et al., 2007).
Ascochyta isolation and preparation: The single spore was transferred to fresh chickpea-agar medium and incubated for 2 weeks at room temperature. The spores of each ascochyta isolate were collected in sterile water and 250 μL of pycnidiospore suspension (approximately 106 mL-1) was inoculated into 75 mL of potato dextrose broth. After incubation for 7 days at room temperature, a mycelial mat formed on the surface of the medium. The mycelial mat was removed from the medium with sterile forceps, rinsed three times in sterilized water, drained on sterile filter paper and the excess water was removed by compressing the mycelial mat with sterile paper towel. The mycelial mat was then wrapped with aluminium foil and stored at -80°C until DNA extraction. Mycelia of 58 isolates of A. rabiei were collected separately.
DNA extraction: Mycelial mats were placed in liquid nitrogen and ground into a fine powder using a pre-cooled pestle and mortar. Total genomic DNA from each isolate was extracted by following a previously reported method (Rogers and Bendich, 1994) with some modifications. The ground mycelia were suspended in 700 μL of preheated (65°C) 2% CTAB buffer containing 1% 2-mercaptoethanol and incubated at 65°C for 30 min. The suspension was mixed with a 600 μL phenol: chloroform: isoamyl alcohol (25:24:1) solution and blended briefly in a vortex mixer. The tubes were centrifuged at 5000 rpm for 10 min and the aqueous layer was transferred to clean tubes. The DNA was precipitated with 1 volume of isopropanol and centrifuged at 14000 rpm for 10 min at room temperature. The supernatant was discarded and each DNA pellet was dissolved with 400 μL TE buffer (10 mM Tris-Cl, 1 mM EDTA, pH 8.0) containing 50 μg mL-1 RNase A. The products were further purified by adding 0.1 volume of 3 M sodium acetate and the DNA was precipitated with cold 95% ethanol. The air-dried DNA in each tube was dissolved in 50 μL TE buffer (pH 8.0). DNA concentration was determined using a NanoDrop ND-1000 Spectrophotometer (NanoDrop Technologies, Wilmington, DE) and quality was assessed by running on a 0.8% agarose gel.
Primer selection and RAPD assay: Preliminary screening was conducted using 28 short sequence oligonucleotide primers reported in the literature. Six short-sequence primers (OPA-03, OPA-13, OPB-07, OPC-01, OPC-20 and OPJ-15) were selected for further analysis based on the polymorphisms and banding patterns obtained after amplification (Navas-Cortés et al., 1998; Udupa et al., 1998; Jamil et al., 2000). The PCR reaction mixture (25 μL) consisted of 1X PCR buffer, 200 μM dNTP, 0.2 μM primer and 1 U Taq DNA polymerase. The DNA concentration was adjusted to 5 ng μL-1 and 5 ng were used as the template. Amplification was carried out using an Eppendorf Master Cycler. Thermal conditions included 35 cycles of denaturation at 94°C for 45 sec (except 2 min for the initial denaturation), annealing for 2 min at 33°C and extension for 2 min at 72°C. A final extension step of 3 min at 72°C was also included. The RAPD products were electrophoresed on 1.5% agarose and visualized using an Alpha Imager 2200 ver. 5.5 Documentation and Analysis System (Alpha Innotech Corp., San Leandro, CA).
Data analysis: The RAPD data were assessed for the presence of polymorphic bands on each DNA sample. The less common bands were recorded among A. rabiei isolates and analysed for genotype grouping. The data were analyzed using PopGene ver. 1.32 software (Yeh et al., 1997), in which genetic distance was calculated according to the formula of Nei (1972). The unweighted pair-group method with the arithmetic averages (UPGMA) was used for cluster analysis using the PAUP 4 software package. The coefficient of similarity was calculated based on genetic distance. The robustness of the dendrogram was determined by bootstrap analysis with 1000 replications using the WinBoot program (Yap and Nelson, 1996).
RESULTS
RAPD analysis: The six primers produced a total of 75 polymorphic DNA bands. The banding patterns of the amplified DNA of 58 A. rabiei isolates were reproducible and showed clear polymorphisms.
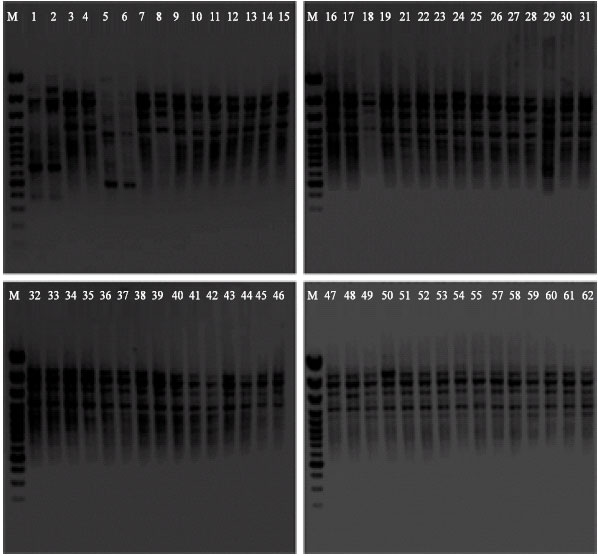 | |
| Fig. 1: | Electrophoretic analysis of RAPD products of 58 Ascochyta rabiei isolates after amplification with the oligonucleotide primer OPJ-15 (DNA of isolates No. 1 and 2 was isolated from lentil and not included in the analysis; numbers 20 and 56 were not included because of missing samples). The M lane in all analyses denotes a 100 bp DNA ladder |
Each primer produced a different number of bands, ranging from zero to nine bands (Table 1). All the A. rabiei isolates shared at least four to seven common bands of the same size, ranging between 400 and 3000 bp. The highest number of visible and prominent bands was produced with primer OPJ-15 (Fig. 1) followed by OPA-03.
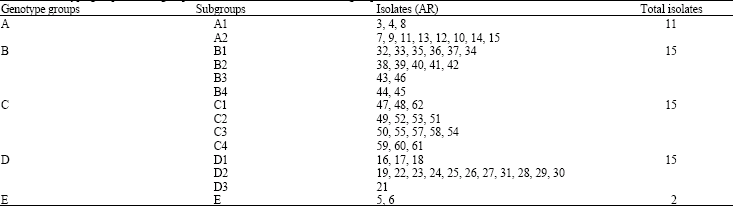
The phylogenetic tree presented in Fig. 2 shows the relationship among the 58 isolates of A. rabiei. Based on this phylogenetic tree, five groups of genotypes (A, B, C, D and E) were identified (Table 2). The A, B, C and D groups were subdivided into 14 subgroups, indicated beside the isolate names on the dendrogram. The groups B (B1, B2, B3 and B4) and D (D1, D2 and D3) comprise the majority of the genotypes and each group included 15 closely related isolates. Group E had no subgroup and formed a unique cluster with only two isolates of A. rabiei (i.e., AR5 and AR6). The groups B and C were more closely related compared to groups A, D and E (Fig. 2). No geographical distribution pattern was observed in the pathogen population except in group B, where all the 15 isolates were from the same location (Table 3, Fig. 2).
The groups derived from analysis of genetic variation did not show a close relationship with previously reported pathotype groupings (Ahmed et al., 2007) (Table 3, Fig. 2). The clusters in the genotype groups are more likely related to the geographical proximity of the chickpea fields than to the pathotype groups of A. rabiei from Alberta.
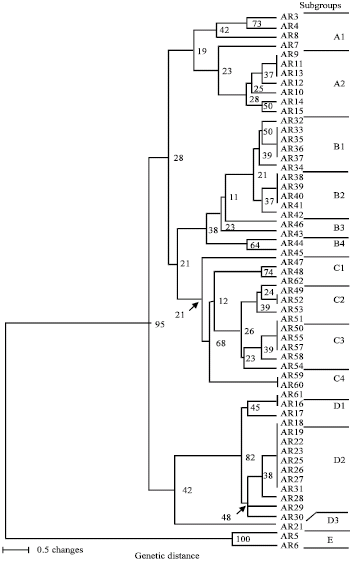 | |
| Fig. 2: | Clustering of 58 Ascochyta rabiei isolates collected in southern Alberta, Canada. The dendrogram is based on similarity coefficients (genetic distance) from random amplified polymorphic DNA (RAPD) analysis and was generated by unweighted paired-group method of arithmetic averages (UPGMA) analysis |
| Table 1: | Band production by selected primers used in RAPD analysis |
 | |
| Table 2: | Genotype groups and subgroups and the isolates included in each group |
 | |
| Table 3: | List of Ascochyta rabiei isolates used for RAPD analysis |
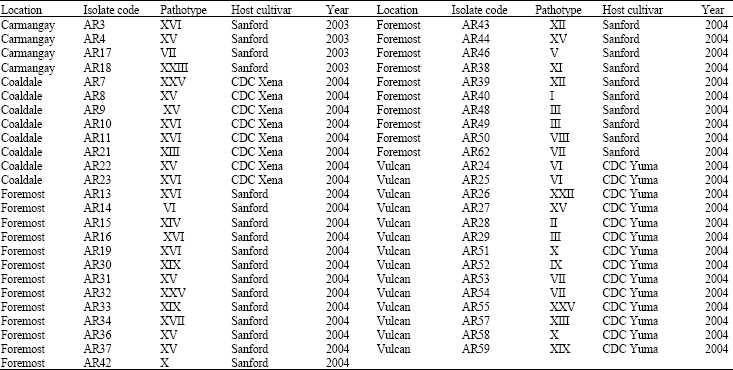 | |
| Source: Derived from Ahmed et al. (2007). List includes only the isolates that have been tested for pathotypes and RAPD profiles | |
DISCUSSION
There are a number of studies on genotypes and pathotypes of ascochyta associated with blight diseases in chickpea worldwide (Ahmed et al., 2005, 2006; Armstrong et al., 2001; Chang et al., 2000, 2007; Chen et al., 2004; Chongo and Gossen, 2003; Chongo et al., 2004; Navas-Cortés et al., 1998; Fischer et al., 1995; Kaiser and Küsmenoglu, 1997; Khan et al., 1999; Morjane et al., 1994; Porta-Puglia et al., 1996; Jamil et al., 2000). These results provide different interpretations of their findings, but share a common conclusion that genotyping of A. rabiei isolates is more reliable than pathotyping. Most of these studies used RAPD markers to study the genotypes and genetic diversity of isolates.
The RAPD analysis conducted in the present study yielded five distinct genotype groups and 14 subgroups in the phylogenetic tree of the 58 A. rabiei isolates collected from southern Alberta. The results from this study indicate that there is a high degree of genetic variation among the A. rabiei isolates, despite the fact that they were collected from four locations (Carmangay, Coaldale, Foremost and Vulcan) that are within a 60 km radius in southern Alberta, Canada. Group A included isolates from Carmangay, Coaldale and Foremost, group B included isolates from Foremost only, group C included isolates from Foremost and Vulcan, group D included isolates from Coaldale, Foremost and Vulcan and group E included two isolates from Carmangay. These observations suggest that the same pathogen genotypes are distributed between and among the locations of southern Alberta. The teleomorph of A. rabiei (Didymella rabiei) is a heterothallic fungus that requires two mating types to produce a sexual stage. Both mating types are reported to occur in the Canadian prairies and the sexual stage of A. rabiei has been observed in chickpea fields (Armstrong et al., 2001; Kaiser and Kusmenoglu, 1997). The sexual stage of the fungus provides an opportunity for genetic recombination and sexual reproduction results in the production of air-borne ascospores that have the potential for long-distance spread. In addition, the pathogen is also seed-borne and disease can be transmitted through seeds from one location to another. All of these factors may have contributed to the high level of genetic diversity within the A. rabiei population of Alberta. Several previous studies investigating the genetic diversity in the pathogen have examined the relationship between molecular markers and pathotypes (Weising et al., 1991; Fischer et al., 1995; Khan et al., 1999; Morjane et al., 1994; Navas-Cortés et al., 1998; Chongo et al., 2004). However, as was observed in present study, no clear correlations were found between markers and pathotype groups. Present results indicate that the markers are more closely related to geographical origin than to the pathotype groupings, which is in agreement with the report by Udupa et al. (1998). Similar results have also been reported for other closely related pathosystems, such as the Pisum sativum-Mycosphaerella pinodes interaction (Su et al., 2006; Zhang et al., 2003), where no relationships between genotype and pathotype groupings were found.
As mentioned earlier, pathogenicity data on the same isolates of A. rabiei (Ahmed et al., 2007) showed no relationship to the genotyping results in this study. Pathotype classification was carried out based on disease severity ratings. Although assessment of pathogenicity provides some classification information, virulence data alone may not reflect the true genetic variability and evolutionary history of the isolates investigated. For example, isolates that are genetically distinct may have similar or identical virulence patterns because they have been subjected to the same selection pressure by a common set of hosts (Zhang et al., 2003). In contrast, molecular markers that are selectively neutral and randomly distributed in the genome may provide additional information on genetic variation of the pathogen population (Pongam et al., 1999). Pathotype or race characterization of A. rabiei has been conducted in several countries, including Canada (Ahmed et al., 2007; Chongo and Gossen, 2003; Reddy and Kabbabeh, 1985; Qureshi and Alam, 1984; Singh, 1990). However, it is difficult to compare the pathotypes or races from different studies since different disease scoring systems, rating scales and differential hosts were used. In addition, variation in inoculum pressure and environmental conditions also makes direct comparisons difficult (Zhang et al., 2003). Hence, genotypic studies represent the best method to determine genetic groupings of A. rabiei isolates.
As a result of the high level of variation in both the virulence and genetic structure of the A. rabiei population in Alberta, it may be difficult to develop chickpea varieties with durable resistance to the pathogen. Furthermore, environmental conditions favourable for ascochyta blight development and sexual reproduction by the pathogen occur in the chickpea growing areas, further complicating disease management. Udupa et al. (1998) suggested that seed dressing with fungicides reduced the migration of a predominant genotype, thereby reducing the incidence of Ascochyta blight on resistant cultivars. The selection of virulent genotypes of the pathogen by the host plant could be reduced by the use of crop rotation to break the disease cycle, thus slowing changes in the population structure of the pathogen.
A single disease management strategy to control ascochyta blight of chickpea may not be effective in Alberta. For successful disease management, an integrated strategy that includes the use of seed dressings with fungicides, resistant genotypes and crop rotation needs to be practiced.
ACKNOWLEDGMENTS
We thank the Alberta Crop Industry Development Fund, the Alberta Agricultural Research Institute and the Alberta Pulse Growers Commission for their financial support. We also acknowledge Alberta Research Council and the Department of Biological Sciences, University of Alberta, for providing laboratory facilities for DNA extraction and RAPD analysis, respectively.
REFERENCES
- Ahmed, H.U., K.F. Chang, S.F. Hwang, B.D. Gossen, R.J. Howardand and T.D. Warkentin, 2006. Components of disease resistance in desi and kabuli chickpea varieties against ascochyta blight. Plant Pathol. J., 5: 336-342.
CrossRefDirect Link - Armstrong, C.L., G. Chongo, B.D. Gossen and L.J. Duczek, 2001. Mating type distribution and incidence of teleomorph of Ascochyta rabiei (Didymella rabiei) in Canada. Can. J. Plant. Pathol., 23: 110-113.
Direct Link - Chang, K.F., H.U. Ahmed, S.F. Hwang, B.D. Gossen and R.J. Howard et al., 2007. Impact of cultivar, row spacing and seeding rate on ascochyta blight severity and yield of chickpea. Can. J. Plant Sci., 87: 395-403.
Direct Link - Chen, W., C.J. Coyne, T.L. Peever and F.J. Muehlbauer, 2004. Characterization of chickpea differentials for pathogenicity assay of ascochyta blight and identification of chickpea accessions resistant to Didymella rabiei. Plant Pathol., 53: 759-769.
Direct Link - Chongo, G., B.D. Gossen, L. Buchwaldt, T. Adhikari and S.R. Rimmer, 2004. Genetic diversity of Ascochyta rabiei in Canada. Plant Dis., 88: 4-10.
Direct Link - Fischer, C., A. Porta-Puglia and W. Barz, 1995. RAPD analysis of pathogenic variability in Ascochyta rabiei. J. Phytopathol., 143: 601-607.
CrossRefDirect Link - Jamil, F.F., N. Sarawar, M. Sarawar, J.A. Khan, J. Geistlinger and G. Kahl, 2000. Genetic and pathogenic diversity within Ascochyta rabiei (Pass.) Lab. populations in Pakistan causing blight of chickpea (Cicer arietinum L.). Physiol. Mol. Plant Pathol., 57: 243-254.
Direct Link - Kaiser, W.J. and I. Kusmenoglu, 1997. Distribution of mating types and the teleomorph of Ascochyta rabiei on Chickpea in Turkey. Plant Dis., 81: 1284-1287.
CrossRefDirect Link - Khan, M.S.A., M.D. Ramsey, A. Corbere, A. Infatino, A. Porta-Puglia, Z. Bouznad and E.S. Scott, 1999. Ascochyta blight of chickpea in Australia: Identification, pathogenicity and mating type. Plant Pathol., 48: 230-234.
CrossRefDirect Link - Morjane, H., J. Geistlinger, M. Harrabi, K. Weising and G. Khal, 1994. Oligonucleotide fingerprinting detects genetic diversity among Ascochyta rabiei isolates from a single chickpea field in Tunisia. Curr. Genet., 26: 191-197.
CrossRefDirect Link - Nene, Y.L., 1982. A review of ascochyta blight of chickpea. Trop. Pest Manage., 28: 61-70.
CrossRefDirect Link - Porta-Puglia, A., P. Crino and C. Mosconi, 1996. Variability in virulence to chickpea of Italian population of Ascochyta rabiei. Plant Dis., 80: 39-41.
Direct Link - Singh, K.B., M.V. Reddy and Y.L. Nene, 1984. International testing of chickpeas for resistance to ascochyta blight. Plant Dis., 68: 782-784.
Direct Link - Su, H., S.F. Hwang, K.F. Chang, R.L. Conner, R.J. Howard and G.D. Turnbull, 2006. Variations in virulence and genome of Mycosphaerella pinodes from field peas in central Alberta, Canada. J. Plant Dis. Prot., 113: 53-60.
Direct Link - Udupa, S.M., F. Weigand, M.C. Saxena and G. Kahl, 1998. Genotyping with RAPD and microsatellite markers resolves pathotype diversity in the ascochyta blight pathogen of chickpea. Theor. Appl. Genet., 97: 299-307.
CrossRefDirect Link - Yeh, F.C., R.C. Yang, T.B.J. Boyle, Z.H. Ye and J.X. Mao, 1997. POPGENE, the User-Friendly Shareware for Population Genetic Analysis. Molecular Biology and Biotechnology Center, Alberta.
Direct Link - Zhang, J.X., W.G.D. Fernando and A.G. Xue, 2003. Virulence and genetic variability among isolates of Mycosphaerella pinodes. Plant Dis., 87: 1376-1383.
Direct Link