
Fidele Tiendrebeogo
Laboratoire de Biochimie and Biologie Moleculaire, CRSBAN/UFR/SVT, Universite de Ouagadougou, 03 BP 7021 Ouagadougou 03, Burkina Faso
V.S. Edgar Traore
Institut de l`Environnement et de Recherches Agricoles (INERA), 01 BP 476 Ouagadougou 01, Burkina Faso
Nicolas Barro
Laboratoire de Biochimie and Biologie Moleculaire, CRSBAN/UFR/SVT, Universite de Ouagadougou, 03 BP 7021 Ouagadougou 03, Burkina Faso
Alfred S. Traore
Laboratoire de Biochimie and Biologie Moleculaire, CRSBAN/UFR/SVT, Universite de Ouagadougou, 03 BP 7021 Ouagadougou 03, Burkina Faso
Gnissa Konate
Institut de l`Environnement et de Recherches Agricoles (INERA), 01 BP 476 Ouagadougou 01, Burkina Faso
Oumar Traore
Institut de l`Environnement et de Recherches Agricoles (INERA), 01 BP 476 Ouagadougou 01, Burkina Faso
Plant Pathology Journal
Year: 2008 | Volume: 7 | Issue: 2 | Page No.: 155-161







ABSTRACT
The aetiology of a disease characterized by severe leaf curling and leaf distortion symptoms in peppers was studied in Burkina Faso. Surveys were conducted in fields planted with sweet pepper (Capsicum annuum) and hot pepper (Capsicum frutescens) to collect leaf samples and assess disease incidence. Collected samples were analyzed using biological tests (mechanical inoculation and whitefly transmission), triple-antibody-sandwich- enzyme linked-immunosorbent assay (TAS-ELISA), Polymerase Chain Reaction (PCR) and sequence analysis. Mechanical inoculation tests were negative while transmission by the whitefly Bemisia tabaci led to infection rates of more than 80%. TAS-ELISA and PCR tests indicated that symptoms observed were associated with begomovirus infections. Involved virus isolates showed pairwise nucleotide identity of more than 99% with Pepper yellow vein mali virus (PepYVMV) and formed with this virus a distinct clade in phylogenetic analyses. Altogether, these results indicated that PepYVMV was the causal agent of leaf curling symptoms in peppers.
PDF Abstract XML References Citation
How to cite this article
Fidele Tiendrebeogo, V.S. Edgar Traore, Nicolas Barro, Alfred S. Traore, Gnissa Konate and Oumar Traore, 2008. Characterization of Pepper yellow vein mali virus in Capsicum sp. in Burkina Faso. Plant Pathology Journal, 7: 155-161.
DOI: 10.3923/ppj.2008.155.161
URL: https://scialert.net/abstract/?doi=ppj.2008.155.161
DOI: 10.3923/ppj.2008.155.161
URL: https://scialert.net/abstract/?doi=ppj.2008.155.161
INTRODUCTION
Geminiviruses (family Geminiviridae) have emerged as economically important plant pathogens worldwide (Harrison and Robinson, 1999). The Geminiviridae family owns its name from a common feature shared by all virus member species which have twin or geminate particles. Most virus species in this family have bipartite genome, the two genomic components being called DNA-A and DNA-B both of which are circular and single-stranded DNA. DNA-A encodes functions necessary for viral replication and encapsidation, while genes on DNA-B are required for viral movement in plants (Lazarowitz, 1992). Some virus species have monopartite genomes and possess DNA-A component only. Part of monopartite begomoviruses have been found associated with a satellite molecule called DNAβ which function is not fully known. However, it was found that DNAβ plays an essential role in symptom induction by the begomovirus to which they are associated and in the increases of the level of viral nucleic acids in infected plants (Saunders et al., 2000; Mansoor et al., 2003).
The Geminiviridae family is composed of four virus genera (Mastrevirus, Curtovirus, Begomovirus and Topocuvirus) which have been distinguished in relation to host range, insect vector and genome organization (Stanley et al., 2005). Begomoviruses have monopartite or bipartite genomes, are transmitted by the whitefly Bemisia tabaci Genn. and infect dicotyledonous plants. They formed the group of viruses formerly known as whitefly-transmitted geminiviruses (WTGs) (Harrison and Robinson, 1999). Common symptoms induced by begomoviruses include leaf-curling, mosaic, vein yellowing or more generalized leaf yellowing, often accompanied by stunting of infected plants and therefore severe yield losses. Besides their transmission by the same vector B. tabaci, begomoviruses exhibit close serological relationships among member species (Harrison and Robinson, 1999). This feature makes it possible to detect almost all begomoviruses with antibodies raised against a particular virus species of the genus. However, serology is not fully suitable for distinguishing between virus species. For this purpose, new molecular tools especially those targeting the viral genome had to be developed (Wyatt and Brown, 1996).
Early studies showed that tubers plants such as cassava (Manihot esculenta Crantz) and vegetables such as tomato (Lycopersicon esculentum L. Mill) and okra (Abelmoshus esculentus L. Moench) were severely affected by WTGs in different parts of Africa (Fargette et al., 1987; Konate et al., 1995; Thresh et al., 1998). Since the last decade, with the emergence of the Bemisia tabaci in West Africa due to resistance to insecticides (Otoidobiga et al., 2003), begomoviruses have become a major constraint to vegetable production in this region. Particularly, in Burkina Faso, new diseases most of which involved leaf curling symptoms appeared in peppers (Capsicum sp.). These symptoms resemble those induced by begomovirus infections and were clearly distinct from mosaic symptoms already reported to be caused by the Pepper veinal mottle virus in the country (Konaté and Traoré, 1999). Although pepper plantings were severely affected, the problem was not payed much attention and even the causal agent of observed the symptoms has not been clearly identified. The research described here was aimed at establishing the etiology of leaf curling symptoms commonly observed in pepper plantings in Burkina Faso.
MATERIALS AND METHODS
Leaf samples collection: During the years 2005 and 2006, pepper fields were surveyed in five localities of Burkina Faso (3 to 10 fields per locality) in some of the country main vegetable crops producing areas (Fig. 1). Virus samples were collected in sweet pepper (Capsicum annuum L.) and hot pepper (Capsicum frutescens) field plantings. Leaves were collected from symptomatic plants and some leaves were also taken from symptomless plants to be used as negative controls. Leaf samples were put in small sampling plastic bags maintained on ice for two to three days. Upon arrival in the laboratory, they were immediately analysed or kept at -20°C for future processing.
Plant inoculation: All transmission tests were done in an insect-proof greenhouse with temperature between 25 and 30°C and 80-90% relative humidity. Seeds of sweet pepper (C. annuum cv. Yolo Wonder) and hot pepper (C. frutescens cv. JBF) were sown in steamed-sterilized soil. Emerged seedlings were transplanted in plastic pots 30 days after sowing and inoculated 10 days later. In a first experiment, leaves from infected pepper plants were ground in 1 M phosphate buffer, pH 8.0 and the sap was subsequently rubbed onto leaves of healthy seedlings. In a second experiment, viruliferous adult insects (B. tabaci) were allowed a 48 h acquisition-access period on diseased plants of both C. annuum and C. frutescens collected from the fields. Three diseased plants per species and from distinct localities were used. Then, 20 insects were placed on healthy pepper seedlings maintained in individual transmission cages for an inoculation access-period of 48 h.
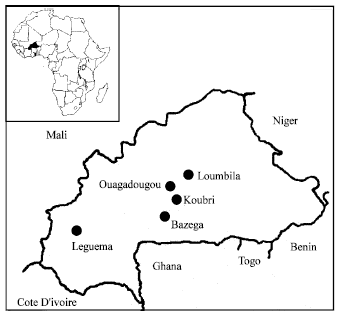 | |
| Fig. 1: | Map of Burkina Faso showing locations of collection of leaf samples |
Transmission tests from every diseased plant were done to five healthy seedlings of each of the two pepper species. Insects were subsequently removed and the plants were treated with a systemic pyrethroid insecticide. All inoculated plants were monitored for symptom development during 45 days post-inoculation.
Seed transmission tests: Seeds were collected form field-infected pepper plants (C. annuum and C. frutescens) and air-dried for three months. A total of 1000 seeds (500 seeds per pepper species) were subsequently sown in sterile soil. After germination, seedlings were maintained in the greenhouse for two months to monitor any development of symptoms.
Serological tests: Leaf samples were analyzed by triple antibody sandwich enzyme linked-immunosorbent assay (TAS-ELISA), as described in Zhou et al. (1997). The monoclonal antibodies (MAbs) to African cassava mosaic virus (ACMV) and Okra leaf curl virus (OkLCuV) were kindly provided by Dr. B.D. Harrison, Scottish Crop Research Institute, UK. Absorbance readings were recorded at 405 nm using a Metertech Σ960 automatic microplate reader.
Nucleic acid extraction, PCR and sequencing: Total nucleic acids were extracted from two selected leaf samples using the CTAB method as described by Doyle and Doyle (1987). Polymerase Chain Reaction (PCR) tests were performed using degenerate primers designed for amplification of the core region of begomoviruses coat protein located on DNA-A. Two pairs of primers (P1 and P2) were used.
| Table 1: | List of begomoviruses from Genbank database used in phylogenetic analyses |
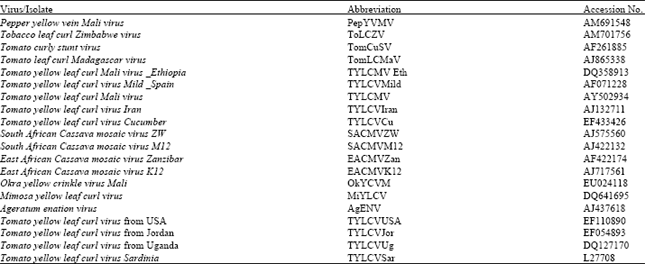 | |
Primer pair P1 consisted of primers PTYv369 (5’-ACGCCCGCTCGAAGGTTCG-3’) and PTYc1023 (5’-CGTCAGCCATATACAATAACAA GGC-3’. Primer pair P2 comprised primers V324 (5’-GCC(CT)AT(GA)TA(TC)AG (AG)AAGCC(AC)AG-3’) and C889 (5’GG(AG)TT(ATG)GA (GA)GCATG(TCA)GTAC ATG-3’). PCR conditions were those described in Navot et al. (1991) and in Wyatt and Brown (1996), respectively for primers pairs P1 and P2. To check amplifications, 10 μL of PCR products were electrophoresed in 1% agarose gels, subsequently stained with ethidium bromide and viewed under UV transillumination. PCR products were sequenced using the Taq terminator sequencing kit (Applied Biosystems) and analyzed on an Applied Biosystems 373A sequencer. To get adequate sequence accuracy, two readings per base (in the 3' to 5' and 5' to 3' directions) were done (Fargette et al., 2004). Sequences obtained were assembled by Seqman (DNASTAR) and a blast search (http://www.interface) was performed to retrieve homologous sequences. The database accession numbers of the begomovirus sequences retrieved are indicated in Table 1. Then, the whole corpus of sequences was aligned by Clustal W (Thompson et al., 1994) and the phylogenetic software Paup (Swofford, 2000) was used to analyse phylogenetic relationships between begomoviruses in the corpus.
RESULTS
Description of field symptoms: The most prominent symptoms were severe distortions of the leaves of infected plants (Fig. 2). There was an overall upward curling of the leaves especially when the plants were infected at an early stage. When infection happened in late stages, symptoms were always present on top leaves whereas basal leaves maintained a relatively normal shape. In addition to leaf curling, leaf distortions also included formation of several swellings with dark green margins resembling mosaic symptoms (Fig. 2a). At the whole plant scale, infected plants were stunted, giving a stacked aspect to the leaves. Moreover, infected plants produced smaller fruits with irregular maturity. Most of the time, disease incidence reached 100% in pepper fields surveyed, regardless of the pepper species and the localities of surveys. However, marked differences were noticed in disease severity, indicating discrepancies in time of disease establishment between fields.
Transmission tests: No plant inoculated mechanically developed any disease symptom. Inoculated plants showed aspects similar to that of healthy uninoculated plants. By contrast, in transmission test through the whitefly B. tabaci, inoculated seedlings of sweet and hot peppers showed symptoms which were similar to those observed on field infected plants. Transmission tests were highly successful as infection rates were between 80 and 100%, regardless of the virus source and the tested plants. In almost all cases, infected plants become stunted and produce no fruit. When seeds from diseased plants were sown, emerged plantlets remained healthy throughout the experiment, indicating the absence of seed transmission.
Begomovirus detection by serology: MAbs directed to ACMV and OkLCuV reacted with all tested symptomatic samples in TAS-ELISA, indicating virus detection in these samples (Table 2).
| Table 2: | Detection of begomoviruses in infected pepper leaf extracts by TAS-ELISA using monoclonal antibodies (MAbs)a |
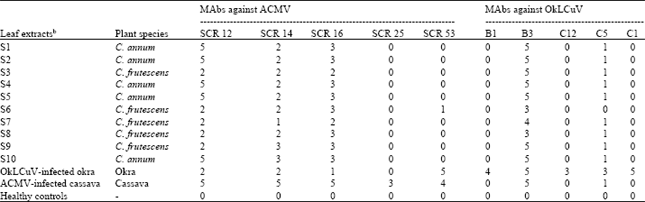 | |
| a: MAbs were directed to African cassava mosaic virus (ACMV) and Okra leaf curl virus (OkLCuV), respectively. Figures in the table are reaction scores that represent intensity of reactions as follows: 5 (A405 nm> 1.8), 4 (1.21-1.8), 3 (0.61-1,2), 2 (0.31-0.6), 1(0.15-0.3), 0 (<0.15), b: Leaf extracts of okra and cassava infected by OkLCuV and ACMV respectively, were used as positive controls. Healthy controls included leaf extracts of C. annum, C. frutescens, cassava and okra, respectively | |
 | |
| Fig. 2: | Field symptoms of severe distortions (a) and upward curling (b) of the leaves of diseased peppers |
Reactions of respective healthy leaf extracts were all negative and allowed clear distinction from those of extracts from diseased leaves. As expected, OkLCuV-infected leaf extract reacted more strongly with anti-OkLCuV MAbs than anti-ACMV MAbs and vice versa. Both OkLCuV-infected and ACMV-infected extracts reacted differently from any of the symptomatic pepper leaf extracts. Half of the MAbs (SCR12, SCR14,SCR16, B3 and C5) reacted with most pepper leaf extracts.
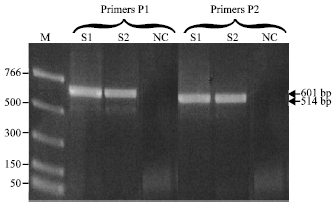 | |
| Fig. 3: | PCR-amplified fragments from four virus samples (S3, S4, S6 and S10) visualized in ethidium-bromide-stained 1% agarose gel. PCR amplification was done using primers pairs P1 (PTYv369/PTYc1023) and P2 (V324/C889) both directed to the core region of begomoviruses. M: PCR markers (sizes in bp); S3 and S6: infected Capsicum frutescens cv. JBF; S4 and S10: infected Capsicum annuum; NC: negative controls (healthy C. frutescens and C. annuum, respectively). Expected fragments from each pair of primers are indicated by arrows |
In contrast, no pepper leaf extract reacted positively with four MAbs (SCR25, B1, C1 and C12) while extract S6 weakly reacted with SCR53. Overall, pepper extracts could be divided in three main groups according to their serological reaction profiles. Group 1 included samples S1, S2, S4, S5 and S10 which reacted strongly with MAbs SCR12 and B3. Samples in group 2 (S3, S7 and S9) reacted strongly with MAb B3 but weakly with SCR12. In the last group (S6 and S8), samples reacted weakly with both MAbs B3 and SCR12.
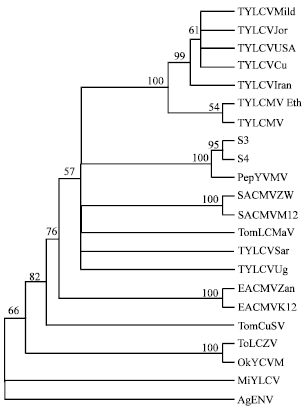 | |
| Fig. 4: | Phylogenetic tree showing relationships among pepper-infecting begomoviruses and other representative begomoviruses based on multiple alignments of nucleotide sequences of the coat protein core region. The tree was constructed by the neighbour-joining method as implemented in PAUP (Swofford, 2000). Bootstrap supports of the nodes are indicated as percentages (from 1000 replicates) branches with bootstrap values less than 50% were collapsed |
PCR amplification and sequence analysis: Two fragments of 601 and 550 bp were amplified using the two sets of degenerate primers P1 (PTYv369/PTYc1023) and P2 (V324/C889), respectively (Fig. 3). Normally, primer set P2 was the one chosen to be used in all PCR tests. Unfortunately, no amplification was obtained from samples S3 and S6 when primer set P2 was used. Therefore, primer set P1 was tested and successfully used for DNA amplification in these two samples.
Sequences of PCR-amplified fragments from samples S3 and S4 were compared to those of 20 begomoviruses retrieved from Genbank database. Sequence identity was 99.8% between S3 and S4. Of begomoviruses included in the analysis, only Pepper yellow vein Mali virus (PepYVMV) had particularly high sequence identity with samples S3 and S4 (99.0 and 99.2%, respectively). Sequence identity between the two samples and other begomoviruses ranged from 72.6 and 84.8%. Phylogenetic analyses indicated that both samples S3 and S4 clustered with PepYVMV and their clade was clearly distinct from any other clade (Fig. 4).
DISCUSSION
Begomoviruses are known to induce a diversity of symptoms in the plants they infect (Harrison and Robinson, 1999). However, leaf curling and other leaf distortions seem to be the most frequent symptoms associated with infections caused by begomoviruses (Paximadis et al., 1999; Wang et al., 2004; Seal et al., 2006). Severe leaf curling symptoms were observed in recent years in peppers in Burkina Faso but no virus associated with such symptoms was reported. In this study, mechanical inoculation tests of the causal agent were unsuccessful. Because whiteflies were the most frequent insects observed in affected pepper fields, transmissions through these insects were attempted. Unlike in mechanical inoculation tests, disease symptoms were readily induced through transmissions by whiteflies, as infections rates were higher than 80%.
Virus transmission by B. tabaci is somewhat specific. Particular genes involves in the ability of this species to transmit plant viruses have been reported (Liu et al., 1998). However, more than 111 plant viruses including begomoviruses, criniviruses, carlaviruses and ipomoviruses are transmitted by the insect (Jones, 2003). Therefore, virus transmissibility by B. tabaci cannot be used as the only criterion for virus identification. In this study, leaf samples taken from diseased pepper plants were positive in TAS-ELISA using antibodies raised against begomoviruses. Added to transmission by B. tabaci, this result indicated that leaf samples were infected by viruses which belong to the Begomovirus genus.
Although transmission by B. tabaci and serological tests, especially using monoclonal antibodies are two useful criteria for identification of begomoviruses, they are not suitable for taxonomic demarcation of begomovirus species (Brown et al., 2001; Fauquet et al., 2003). The core CP gene was successfully amplified from diseased pepper leaf samples by PCR. This result is consistent with transmission and serological data and confirms the begomoviral aetiology of the disease observed in peppers. Moreover, analysis of sequences obtained from PCR-amplified fragments indicated high pairwise nucleotide identity (>99%) with Pepper yellow vein mali virus (PepYVMV). No other begomovirus species had pairwise nucleotide identity exceeding 85%. Nucleotide identity not less than 89% has been advocated as the threshold for demarcating begomovirus species (Fauquet et al., 2003). Therefore, begomoviruses identified in diseased pepper leaf samples belong to the PepYVMV species (Vadivukarasi et al., 2007).
Altogether, the results obtained in this study indicated that leaf curling symptoms observed in sweet and hot pepper in Burkina Faso were caused by PepYVMV. However, Zhou et al. (2008) recently proposed the name tomato yellow leaf crumple virus (ToYLCrV) instead of PepYVMV because of marked discrepancies in symptoms caused by the virus in pepper and tomato. They reported that first symptoms induced by the virus were upcurled leaves and yellow vein symptoms in pepper. But, subsequently, surveys in a few West African country including Burkina Faso, Benin, Ghana, Niger, Senegal and Togo indicated that the virus was mostly associated with yellow leaf crumple symptoms in tomato. In the present study, the virus incidence in tomato was not assessed. However, high incidences observed in pepper fields indicated that the virus is widely spread in pepper.
ACKNOWLEDGMENTS
We are indebted to the following institutions for providing laboratory facilities for this work: CRSBAN/UFR-SVT (University of Ouagadougou) and INERA (CNRST, Burkina Faso). This study is part of F. Tiendrébeogo's Ph.D project supervised by Dr. O. Traoré (INERA) and Prof. A.S. Traoré (University of Ouagadougou).
REFERENCES
- Brown, J.K., A.M. Idris, I. Torres-Jerez, G.K. Banks and S.D. Wyatt, 2001. The core region of the coat gene is highly useful for the establishing the provisional identification and classification of begomoviruses. Arch. Virol., 146: 1581-1598.
Direct Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Fargette, D., J.C. Thouvenel and C. Fauquet, 1987. Virus content of cassava infected by African cassava mosaic virus. Ann. Applied Biol., 110: 65-73.
CrossRef - Fargette, D., A. Pinel, Z. Abubakar, O.Z. Traore and C. Brugidou et al., 2004. Inferring the evolutionary history of rice yellow mottle virus from genomic, phylogenetic and phylogeographic studies. J. Virol., 78: 3252-3261.
CrossRefDirect Link - Fauquet, C.M., D.M. Bisaro, R.W. Briddon, J.K. Brown and B.D. Harrison et al., 2003. Virology division news: Revision of taxonomic criteria for species demarcation in the family Geminiviridae and an updated list of begomovirus species. Arch. Virol., 148: 405-421.
CrossRef - Harrison, B.D. and D.J. Robinson, 1999. Natural genomic and antigenic variation in whitefly-transmitted Geminiviruses (Begomoviruses). Annu. Rev. Phytopathol., 37: 369-398.
CrossRefDirect Link - Jones, D.R., 2003. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol., 109: 195-219.
CrossRefDirect Link - Konate, G., N. Barro, D. Fargette, M.M. Swanson and B.D. Harrison, 1995. Occurrence of whitefly-transmitted geminiviruses in crops in Burkina Faso, and their serological detection and differentiation. Ann. Applied Biol., 126: 121-129.
CrossRef - Lazarowitz, S.G. and R.J. Shepherd, 1992. Geminiviruses: Genome structure and gene function. Crit. Rev. Plant Sci., 11: 327-349.
CrossRefDirect Link - Liu, S., R.W. Briddon, I.D. Bedford, M.S. Pinner and P.G. Markham, 1998. Identification of Genes Directly and Indirectly Involved in the Insect Transmission of African cassava mosaic geminivirus by Bemisia Tabaci. Virus Genes, 18: 5-11.
CrossRef - Mansoor, S., R.W. Briddon, Y. Zafar and J. Stanley, 2003. Geminivirus disease complexes: An emerging threat. Trends Plant Sci., 8: 128-134.
CrossRefDirect Link - Navot, N., E. Pihersky, M. Zeidan, D. Zamir and H. Czosneck, 1991. Tomato yellow leaf curl virus: A whitefly-transmitted geminivirus with a single genomic component. Virology, 185: 151-161.
CrossRef - Otoidobiga, L.C., C. Vincent and R.K. Stewart, 2003. Susceptibility of field populations of adult Bemisia tabaci Gennadius (Homoptera: Aleyrodidae) and Eretmocerus sp. (Hymenoptera: Aphelinidae) to cotton insecticides in Burkina Faso (West Africa). Pest Manage. Sci., 59: 97-106.
Direct Link - Paximadis, M., A.M. Idris, I. Torres-Jerez, A. Villarreal, M.E.C. Rey and J.K. Brown, 1999. Characterization of tobacco geminiviruses in the old and new world. Arch. Virol., 144: 703-717.
PubMed - Saunders, K., I.D. Bedford, R.W. Briddon, P.G. Markham, S.M. Wong and J. Stanley, 2000. A unique virus complex causes Ageratum yellow vein disease. Proc. Natl. Acad. Sci. (USA)., 97: 6890-6895.
Direct Link - Seal, S.E., F. VandenBosch and M.J. Jeger, 2006. Factors influencing Begomovirus evolution and their increasing global significance: Implications for sustainable control. Crit. Rev. Plant Sci., 25: 23-46.
CrossRefDirect Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Vadivukarasi, T., K.R. Girish and R. Usha, 2007. Sequence and recombination analyses of the Geminivirus replication initiator protein. J. Biosci., 32: 17-29.
CrossRef - Wang, X., Y. Xie and X. Zhou, 2004. Molecular characterization of two distinct begomoviruses from papaya in China. Virus Genes, 29: 303-309.
CrossRef - Wyatt, S.D. and J.K. Brown, 1996. Detection of subgroup III geminivirus isolates in leaf extracts by degenerate primers and polymerase chain reaction. Phytopathology, 86: 1288-1293.
CrossRefDirect Link - Zhou, X., Y. Liu, L. Calvert, C. Munoz, G.W. Otim-Nape, D.J. Robinson and B.D. Harrison, 1997. Evidence that DNA-A of a Geminivirus associated with severe cassava mosaic disease in Uganda has arisen by interspecific recombination. J. Gen. Virol., 78: 2101-2111.
CrossRefDirect Link - Zhou, Y.C., M. Noussourou, T. Kon, M.R. Rojas and H. Jiang et al., 2008. Evidence of local evolution of tomato-infecting begomovirus species in West Africa: Characterization of tomato leaf curl Mali virus and tomato yellow leaf crumple virus from Mali. Arch. Virol., 153: 693-706.
CrossRefPubMedDirect Link