
A.O. Ogaraku
Unit of Plant Science and Biotechnology, Department of Biological Sciences, Nasarawa State University, P.M.B. 1022, Keffi, Nigeria
Plant Pathology Journal
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 114-117







ABSTRACT
A study was undertaken to determine the separate and combined affects of Fusarium oxysporum f. sp. tracheiphilum and Meloidogyne incognita on growth and yield of cowpea variety Moussa-local. It was observed that growth and yield components of the cowpea were generally reduced in all treatments as compared to that of control. Single infection with only nematode caused more growth and yield reductions than infection with only fungus. Reduction in the growth and yield components were generally higher in simultaneous infection than infection with either of the pathogens. Growth and yield reduction in combined infection did not vary significantly from those caused by successive infection where either of the pathogens was made to precede the other. Infection with both pathogens caused significant increases in the root galls, number of eggs and juveniles when compared with infection with nematode only.
PDF Abstract XML References Citation
How to cite this article
A.O. Ogaraku, 2008. Separate and Combined Effects of Fusarium oxysporum f. sp. tracheiphilum and Meloidogyne incognita on Growth and Yield of Cowpea (Vigna unguiculata L. Walp) Var. Moussa-Local. Plant Pathology Journal, 7: 114-117.
DOI: 10.3923/ppj.2008.114.117
URL: https://scialert.net/abstract/?doi=ppj.2008.114.117
DOI: 10.3923/ppj.2008.114.117
URL: https://scialert.net/abstract/?doi=ppj.2008.114.117
INTRODUCTION
Cowpea is an important food legume especially in the semi-Arid regions of the tropics. About 80% of the world cowpea is produce in west Africa and principally in Nigeria (Mukhtar and Aminu, 2005). Cowpea provides a cheap source of protein. The chemical composition of cowpeas is similar to that of most edible legumes. It contains about 24% protein, 62% soluble carbohydrates and small amounts of other nutrients (Singh et al., 1997). Cowpea protein gave an apparent protein digestibility of 72% (Bressani et al., 1982).
As important as this crop is, its production in Nigeria is hampered by many constraints including diseases from nematodes and fungi (Ojuederie, 2002). In USA and elsewhere, reports abound of field occurrence of root-knot nematodes and phytopathogenic fungi disease complexes on crops (Morgan-Jones et al., 1984). These complexes are characterized by increased severity of disease symptoms. However, research on these complexes in Nigeria remains scanty (Iheukwumere et al., 1996). This is probably because symptoms of infection are sometimes erroneously taken for other causes like nutrient deficiency, lack of moisture or worse still, ignored (Idowu et al., 1990).
The present study is an attempt at bridging the information gap on the infection complexes of crops involving a nematode and a soil fungus, particularly on cowpea in Nigeria.
MATERIALS AND METHODS
Seeds of cowpea var. Moussa-local obtained from International Institute for Tropical Agriculture (IITA) Ibadan previously established for its susceptibility (Ogaraku and Onovo, 2006) to Meloidogyne incognita were used for this study. The research was conducted in the Green house of the Botanical Nursery of University of Jos. The research took place between December/January, 2005/2006 and at the onset of the early rains of the wet season in April/May 2006. Mean temperature of the green house was 22.4°C with a minimum of 17.6°C and maximum of 27.6°C. Mean relative humidity of the green house was 40 with a minimum of 27 and maximum of 53, with a daily 12 h light period.
The seeds were surface sterilized for 5 min in 1.05% sodium hypochlorite and rinsed for 5 min in distilled water prior to sowing (Koenning and McClure, 1981). Seeds were sown in 2 kg of soil steam sterilized at 65°C for 90 min.
Thirty 4-litre plastic pots (15 cm diameter) were divided into six groups of fives. So that a total of thirty pots were used for this study. Four cowpea seeds were planted in each pot, at two-leaf stage, the seedlings were thinned down to one per pot.
Source of nematode and extraction: The roots of Celosia argentea on which Meloidogyne incognita was cultured in the green house after identifying its perineal pattern with reference to those published by Eisenback et al. (1981), were macerated in a warring blender for 30 sec in about 250 mL of processed tap water. The comminute was passed through a bank of three Cobb (1917) sieves of 100, 200 and 250 meshes in that order from the top. Concentrated M. incognita egg suspension was obtained from the filtrate by using the settling siphon method of Caveness (1975). The suspensions were examined under a stereoscopic microscope and a few juveniles that passed through the 250 mesh sieve were picked out with eyelash affixed unto a mounted needle. Estimation of nematode eggs was done by counting the number of eggs in 1 mL of a homogenized suspension of the inoculums using a stereoscopic microscope at 40X magnification then projecting to the requisite quantity that would give the required number of eggs to be used for inoculation. One milliliter of the egg suspension was estimated to contain 500 eggs; therefore 8 mL would contain 4,000 eggs. The pathogenicity of the nematode on plants increases with an increase in its population (Oostenbrink, 1966).
Source of fungi and propagation: Fusarium oxysporum f. sp. tracheiphilum used was isolated from soil using Colocynthis citrullus L. seeds; melon seeds as bait (Thomason et al., 1959). The isolated fungus was sub-cultured and maintained on Potato Dextrose Agar (PDA) for multiplication at laboratory room temperature of 27±3°C for six days.
Plants were inoculated by pouring the egg suspension containing the required number of eggs into a shallow trench around the base of the plant and covering with sterile top soil (Iheukwumere et al., 1996). Seedlings were also inoculated with the fungus propagated and maintained on potato dextrose agar by introducing 5 g of its mycelia into shallow holes made in the root rhizosphere of the cowpea. The 5 g mycelia were deposited in a 5 L Hozelok sprayer containing 20 mL of distilled water, the content sprayed around the base of the seedlings until runoff (Thomason at al., 1959) then covered with sterile top soil (Iheukwumere et al., 1996).
At two-leaf stage, the following treatments were applied to them:
| • | Inoculation of plants with 4,000 eggs of Meloidogyne incognita only (N). |
| • | Inoculation of plants with 5 g of F. oxysporum f. sp. tracheiphilum mycelia only (F). |
| • | Simultaneous inoculation of plants with 4,000 eggs of the nematode and 5 g of fungus (N + F). |
| • | Inoculation of plants with 4,000 eggs at the two-leaf stage followed by inoculation with 5 g of fungus mycelia 7 days later (N + f). |
| • | Inoculation of plants with 5 g of fungus mycelia at two-leaf stage followed by inoculation with 4,000 eggs of the nematode 7 days later (F + n). |
| • | The uninoculated control plants (c). |
The plants were placed on a bench kept in the green house. A Completely Randomized Design (CRD) was used with 5 replicates per treatment. Plants were watered on alternate days and inspected for symptom expression weekly.
Galls were rated on a scale of 0-5 according to the method of Taylor and Sasser (1978) as follows: 0 = no gall; 1 = ≤20% of root galled; 2 = 21-40% of root galled; 3 = 41-60% of root galled; 5 = ≥ 80% of the root galled. Number of eggs per plant was extracted from each root system using the sodium hypochlorite method of Hussey and Barker (1973). While number of second stage juvenile was extracted from each pot soil using pie-pen modification of Baerman funnel method (Ononuju and Fawole, 1999).
At the termination of the experiment 120 days after planting, data were collected on various growth and yield parameters and subjected to analysis of variance by Duncan’s multiple range test.
RESULTS
Separate infection of plants with the nematode or fungus caused reductions in the various growth and yield components of the cowpea as compared to the control (Table 1). These reductions were statistically significant for shoot length, root dry weight, number of pods and pod dry weight. However, shoot fresh and dry weights, root length, root fresh and dry weights of control plants were significantly higher than single infection with the fungus or the nematode. Infection with only nematode caused higher reduction in the growth and yield components than infection with only fungus although this was statistically significant.
In general, concomitant infection led to highest reductions in most of the growth and yield components, these were statistically different from single infection with either of the pathogens. This trend was similarly established for most of the other parameters between simultaneous and successive infection. The gall rating followed this order of decreasing magnitude: N + F, N + f, F + n and N (Table 2).
| Table 1: | Effects of Fusarium oxysporum and Meloidogyne incognita on growth and yield components of cowpea variety Mousa-local |
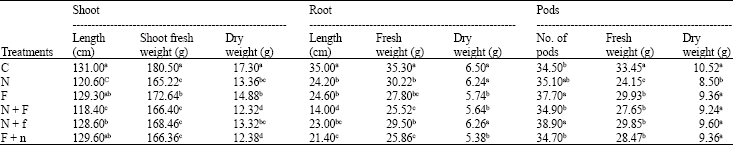 | |
| Each value is a mean of five replicates. Means followed by the same letter in each vertical column are not significantly different according to Duncan’s multiple range text (p = 0.05), C = Uninoculated; N = nematode; F = fungus; N + F = Nematode and fungus inoculated simultaneously; N + f = nematode inoculation following by that of fungus 7 days later; F + n = fungus inoculation followed by that of nematode 7 days later | |
| Table 2: | Effects of infection of Fusarium oxysporum and Meloidogyne incognita on root gall and nematode reproduction |
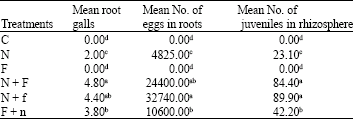 | |
| Each value is a mean of five replicates. Means followed by the same letter in each vertical column are not significantly different according to Duncan's multiple range test (p = 0.05), O = No galls; 1 = ≤20% of the root system galled; 2 = 21-40% of the root system galled; 3 = 41-60% of the root system galled; 4 = 61-80% of the root system galled; 5 = ≥80% of the root system galled (Taylor and Sasser, 1978) | |
The number of eggs extracted from the roots of plants with both the fungus and nematode infection were higher than those treated with nematode alone (Table 2). Similarly, the number of juveniles extracted followed the same trend as that reported for the number of eggs (Table 2).
DISCUSSION
The study has shown that simultaneous co-inflection of cowpea generally decreased growth and yield parameters of this crop more than single infection. This may be the result of competition for nutrients by both pathogens which consequently interrupted the host metabolism thus decreasing growth and development. This is consistent with the findings of Thomason et al. (1959), who reported on the relationship of the root-knot nematode, Meloidogyne juveniles, to Fusarium wilt of cowpea. The contrasting results obtained in the various growth and yield factors in which some were significantly reduced and others not, could be the result of different growth substances (usually produced in varying concentrations) (Hussey, 1985) that were elicited by the interaction of each pathogen with the host. These substances may interact on their own and cause different alterations in the various plant tissues and organs thus causing them to respond differently to the infection. Furthermore, the fact that infection does not necessarily cause diseases at all times in all parts of an infected plant (Mathews, 1981) may also account for why some parameters were significantly impacted by the treatment while others were not.
In gall severity, it appeared that the presence of the fungus in mixed inoculations enhanced gall formation in the roots of inflected plants. This is because plants inoculated with both the nematode and the fungus have significantly higher gall ratings than those infected with nematode alone. This trend was also established for the number of eggs and second stage juveniles, an indication that combined inoculations with both pathogens significantly increased egg and juvenile production by the nematode more than in single inoculation with nematode. This is because nematode infection often causes specific changes in the protein metabolism of infected plants, which may well interact with the physiological changes caused by the fungus to enhance nematode reproduction (Trabulsi et al.,1980). Since nematode infection is known to affect the metabolism and chemical composition of a plant (Raut and Sethi, 1980), it is possible that the Meloidogyne incognita predisposed the cowpea in such a way that its metabolism and physiology became so seriously impaired that the cowpea became less capable of resisting infection by the fungus hence the enhanced nematode development and pathogenicity (Thomason et al., 1959). It is also possible that the nematode feeding on the root tissues impaired water and nutrient uptake by destroying and blocking conducting vessels, which may have weakened the plant to the advantage of the fungus.
This research has established a synergistic relationship between F. oxysporum f. sp. tracheiphilum and M. incognita in cowpea var. moussa-local with consequent reduction in the growth and yield components of the crop. Since both pathogens occur wherever cowpea is grown, effective combative method would be to raise varieties that are resistant to both the fungus and the nematode.
REFERENCES
- Caveness, F.E., 1975. Other contributions: A simple siphon method for separating nematodes from excess water. Nematropica, 5: 30-32.
Direct Link - Hussey, R.S. and K.R. Barker, 1973. Comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Dis. Rep., 57: 1025-1028.
Direct Link - Iheukwumere, C.C., G.L. Atiri, B. Fawole and K.E. Dashiel, 1996. Interaction between soybean mosaic potyvirus and Meloidogyne incognita infection. Afr. Crop Sci. J., 4: 351-357.
Direct Link - Koenning, S.R. and M.A. McClure, 1981. Interaction of two potyviruses and Meloidogyne incognita in chili pepper. Phytopathology, 71: 404-408.
Direct Link - Raut, S.P. and C.L. Sethi, 1980. Studies on the pathogenicity of Meloidogyne incognita on soybean. Indian J. Nematol., 10: 166-174.
Direct Link - Trabulsi, I.Y., M.A. Ali and M.E. Abd-Elsamea, 1980. Response of soybean cultivars to infection by Meloidogyne incognita and Rhizobium japonicum alone and in combination. Nematol. Mediterranean, 80: 171-175.
Direct Link