
Shirin Farzadfar
Department of Plant Protection, College of Agriculture, Tehran University, Karaj, Iran
Ali Ahoonmanesh
Department of Plant Pathology, College of Agriculture, Esfahan University of Technology, Esfahan, Iran
Gholam Hossein Mosahebi
Department of Plant Protection, College of Agriculture, Tehran University, Karaj, Iran
Kazusato Ohshima
Laboratory of Plant Virology, Faculty of Agriculture, Saga University, P.O. Box 840-8502, Saga, Japan
Mina Koohi-Habibi
Department of Plant Protection, College of Agriculture, Tehran University, Karaj, Iran
Reza Pourrahim
Department of Plant Virology, Plant Pests and Diseases Research Institute, P.O. Box 19395-1454, Tehran, Iran
Ali Reza Golnaraghi
Department of Plant Protection, Science and Research Branch, Islamic Azad University, P.O. Box 14515-775, Tehran, Iran
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 291-298







ABSTRACT
The biological and molecular characteristics of nine Cauliflower mosaic virus (CaMV) isolates from infected cauliflower plants (Brassica oleracea var. botrytis), grown in different Iranian regions, were evaluated. Based on mechanical inoculation to a range of herbaceous indicators, the Iranian CaMV isolates were distinguished for their different capacity to infect cruciferous plants and Datura stramonium. Only a part of the isolates under study infected kohlrabi (B. oleracea var. gongylodes) and D. stramonium seedlings, whereas, in experimental trials, all of them were transmitted on healthy cauliflowers by green peach aphids (Myzus persicae). In PCR, 1.58 kb genomic fragments, corresponding to the CaMV ORF VI, were amplified. Their sequences showed close identities (96.1-96.7%) with that of D/H CaMV-Hungarian isolate. In phylogenetic analysis the isolates under study were clustered in a separate branch, together with the non-North American isolates group. The Iranian isolates were also well differentiated from other exotic CaMV isolates by restriction analysis using Hpy99I.
PDF Abstract XML References
How to cite this article
Shirin Farzadfar, Ali Ahoonmanesh, Gholam Hossein Mosahebi, Kazusato Ohshima, Mina Koohi-Habibi, Reza Pourrahim and Ali Reza Golnaraghi, 2007. Partial Biological and Molecular Characterization of Cauliflower mosaic virus Isolates in Iran. Plant Pathology Journal, 6: 291-298.
DOI: 10.3923/ppj.2007.291.298
URL: https://scialert.net/abstract/?doi=ppj.2007.291.298
DOI: 10.3923/ppj.2007.291.298
URL: https://scialert.net/abstract/?doi=ppj.2007.291.298
INTRODUCTION
CaMV, the type member of the genus Caulimovirus, is one of the most common and important viruses of brassica crops in the world. This virus is widespread in temperate regions wherever species of Brassica are grown (Sutic et al., 1999). CaMV has isometric particles about 50 nm in diameter, composed of 420 capsid protein subunits and a circular double-stranded DNA genome of 8 kbp (Cheng et al., 1992). The CaMV genome contains seven major ORFs (I to VII) and is replicated by the reverse transcription of an RNA intermediate (Mason et al., 1987; Haas et al., 2002). This virus induces a variety of systemic symptoms (chlorosis, mosaic, vein clearing and stunting) on many cruciferous plants, particularly on various Brassica campestris and B. oleracea cultivars, often in mixed infection with Turnip mosaic virus (TuMV) (Shepherd, 1981). All CaMV isolates can infect a wide variety of cruciferous species, but only a few of them, i.e., D4 and W260, can infect solanaceous species, including those of Datura and Nicotiana genera (Daubert et al., 1984; Daubert and Routh, 1990; Anderson et al., 1991; Qiu et al., 1997). CaMV is naturally transmitted by aphids in a non-circulative manner (Palacios et al., 2002), except for some isolates that are not at all transmitted (Lung and Pirone, 1973) and it uses the helper strategy for the transmission process (Pirone and Blanc, 1996; Gray and Banerjee, 1999). At least 27 aphid species are recognized vectors of CaMV (Kennedy et al., 1962). This virus is not transmissible by seed or pollen (Blanc et al., 2001). CaMV isolates have been previously characterized and differentiated on the basis of their reactions on various indicator plants, including turnip, kohlrabi and some solanaceous species (Lung and Pirone, 1972; Schoelz et al., 1986; Al-Kaff and Covey, 1995) and of the nucleotide sequence of their ORF VI gene. This ORF, in fact, is characterized by having high levels of variation if compared to the other CaMV ORFs and is therefore used to distinguish different isolates of the virus (Sanger et al., 1991; Chenault and Melcher, 1994a, b; Pique et al., 1995).
In recent years, farm acreage of brassica crops, especially cauliflower, has significantly increased in Iran; the area covered by cauliflower passed from 800 ha with 20,000 tones production in 2000 to 1000 ha with 25,000 tones in 2003 (FAO, 2000, 2003). CaMV is a naturally occurring virus that infects cruciferous hosts in Iran (Shahraeen et al., 2003; Farzadfar et al., 2005, 2007), however, there is no information about the biological and molecular properties of Iranian isolates of the virus. In this study nine CaMV isolates from cauliflowers collected in different geographical areas of Iran were characterized using biological approaches. The molecular properties of these isolates were also studied by polymerase chain reaction (PCR) and sequencing of their ORF VI.
MATERIALS AND METHODS
Plant materials and serological assays: Cauliflower leaves with symptoms of deformation, mosaic, mottling, necrosis, rugosity, stunting, vein banding, vein clearing and/or yellowing were collected from 32 fields in Azarbayejan-e-gharbi, Esfahan, Khorasan, Fars, Tehran and Qazvin provinces of Iran during the summers of 2004 and 2005. These samples were tested for CaMV by double-antibody sandwich enzyme linked immunosorbent assay (DAS-ELISA) (Clark and Adams, 1977) using specific polyclonal antibodies purchased from Loewe (Sauerlach, Germany). In ELISA tests, all buffers were prepared according to the manufacturer's instructions. One hundred microliter of each leaf extract, ground in 1:5 (wt/vol) extraction buffer, were added to the wells of polystirene plates (Maxisorb, NUNC, Denmark) previously coated with 1:200 CaMV-IgG in carbonate buffer and incubated overnight at 4°C. The IgG-conjugated antibody incubation was done for 3 h at 35°C. Samples were considered positive if the absorbance at 405 nm, measured by using Multiscan-334 (Lab system, Finland), was greater than or equal to three times the healthy control mean values after 60 min of reaction with p-nitrophenyl phosphate at 1 mg mL-1 in substrate buffer.
Host range studies: Samples with positive ELISA readings were ground in 0.1 M Na-phosphate buffer (pH 7.4) containing 0.02% 2-mercaptoethanol and inoculated into radish (Raphanus sativus) dusted with carborundum. Virus isolates were purified by three serial single-lesion inoculation passages on turnip (Brassica rapa) or Datura stramonium. For subsequent experiments, nine CaMV isolates, different for provenience and severity of symptoms (Table 1), were propagated in radish. The infected leaf extracts (1:5 wt/vol) were rubbed onto a range of herbaceous plants shown in Table 2. In this study, the most common cruciferous weeds present in brassica fields, including Eruca sativa (rocket salad), Hirschfeldia incana (shortpod mustard), Rapistrum rugosum (rugose rapistrum), Sisymbrium loeselii (small tumble-mustard) and S. irio (London rocket) were also tested. Plants used in the host range studies were inoculated at four-six leaves stage, using at least three plants/indicator and kept under greenhouse conditions for 4-6 weeks; afterwards symptoms were recorded and virus presence determined by ELISA.
Aphid transmission: Aphid transmissibility of the selected CaMV isolates (Table 1) was performed with green peach aphids (Myzus persicae Sulzer). Aphids were starved for 1 hr, given a 5 min acquisition access on young leaves of B. rapa plants which had been previously infected (from 2-3 weeks) with the isolates under study and then moved on non-infected young turnips. Aphids were left to feed for 24 h and then killed by spraying with Confidor (Bayer, Germany). For each plant, 5 aphids were used and at least 10 test plants were assayed. The presence of CaMV in the inoculated plants was tested by ELISA.
DNA extraction and polymerase chain reaction (PCR): Upstream and downstream primers used in this study were, respectively, CaMV-6-F (5’-ACGCGTCGACATGGAGAACATAGAAAAAC-3’) and CaMV-6-R (5’-ACGCGTCGACTCAATCCACTTGC TTTGAA-3’), designed by U. Melcher (Department of Biochemistry and Molecular Biology, Oklahoma State University). These primers, synthesized by MWG Biotech Co. (Ebersberg, Germany), direct the amplification of about 1,580 bp fragments containing the complete ORF VI sequence, except for small regions on the 3' and 5' ends.
| Table 1: | Iranian isolates of Cauliflower mosaic virus studied in this work |
 | |
| Table 2: | Symptomatological response of herbaceous indicator plants inoculated with different Iranian Cauliflower mosaic virus isolates |
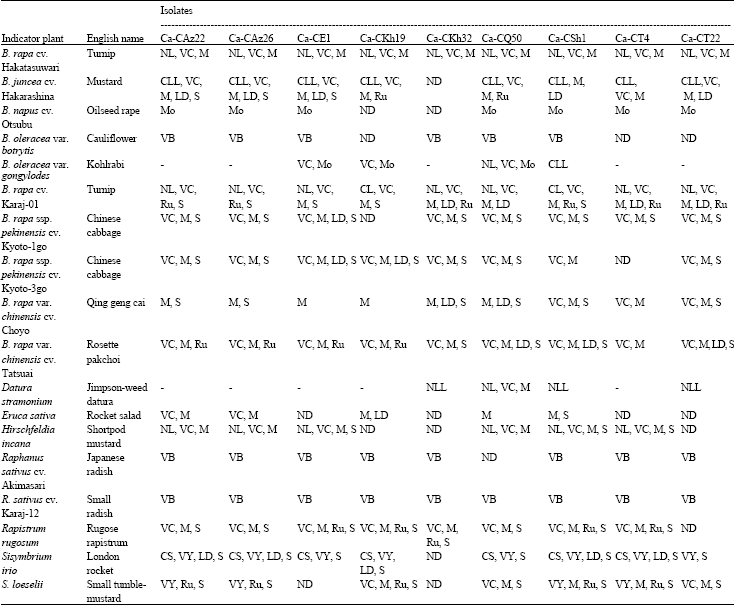 | |
| CL, Chlorotic Lesions; CS, Chlorotic Spots; LD, Leaf Deformation; M, Mosaic; Mo, Mottling; NL, Necrotic Lesions; NLL, Necrotic Local Lesions; Ru, Rugosity; S, Stunting; VB, Vein Banding; VC, Vein Clearing; VY, Vein Yellowing; ND, Not Determined; -, No Infection | |
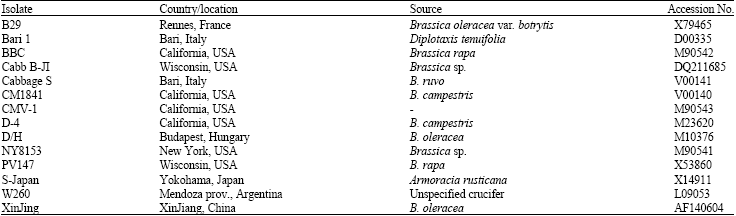
| Table 3: | List of available Cauliflower mosaic virus (CaMV) sequences in GenBank, used for comparative analysis with Iranian isolates |
 | |
DNA was extracted from young turnip infected leaves 30 days after inoculation (Agama et al., 2002) and used as template for PCR analyses. Healthy turnip extracts were used as negative controls. PCR amplification was conducted in 50 μL reaction solution containing 5 μL DNA template, 1 μL of each primer (20 pmol μL-1), 5 μL of 10X reaction buffer (200 mM Tris-HCl, 500 mM KCl, pH 8.4), 1.5 μL MgCl2 (50 mM), 1 μL dNTPs (10 mM) and 2.5 units Taq DNA polymerase (CinnaGen Inc., Tehran, Iran). Thermocycling was done as follows: 94°C for 2 min, 30 cycles of 94°C for 30 sec, 45°C for 1.5 min, 72°C for 1 min and finally 72°C for 10 min. PCR reactions were done in a Primus (MWG Biotech Co., Germany) thermal cycler. PCR products and DNA ladder (GeneRulerTM 1 kbp DNA Ladder, Farmentas, Lithuania) were fractionated by electrophoresis in 1% agarose gel, in presence of 1 μg mL-1 ethidium bromide (Sambrook et al., 1989).
Cloning, sequencing and sequence analysis: The amplified DNA for each CaMV isolate under study (Table 1) was cleaned from the amplification reaction mixture using Nucleospin (Macherey-Nagel, Germany) and cloned into pGEM-T Easy vector (Promega Corp., Madison, WI) according to the manufacturers’ instructions. Sequences from both strands of the cloned DNA in three independent clones were determined by the custom sequencing service of MWG Biotech Co. (Germany) using standard M13 primer pairs. The sequences obtained were compared among them and with those of other CaMV isolates available in GenBank using Basic Local Alignment Search Tool (BLAST) (Altschul et al., 1997) at the National Centre for Biotechnology Information. Multiple sequence alignments were generated by the Clustal W program (Thompson et al., 1994). Phylogenetic trees were constructed using MegAlign program ver. 5.00 from the DNASTAR package (DNASTAR, Madison, WI, USA) (Burland, 2000). In this study, the corresponding region of Horseradish latent virus (HRLV) was used as outgroup in the alignment of CaMV sequences. For further analysis and to obtain potential restriction enzyme(s) for their differentiation, Iranian and other exotic CaMV isolates (Table 1 and 3) were mapped and compared using Mapdraw program (ver. 5.00, DNASTAR, Burland, 2000).
RESULTS
Serological assays: Almost all cauliflower leaf samples (292/323, corresponding to 90.4% of infection) collected from 32 commercial fields over six provinces surveyed, reacted positively in DAS-ELISA using CaMV-specific antibodies.
Host range and symptoms: The herbaceous host range of the CaMV isolates from cauliflower of different Iranian provinces (Table 1) were substantially similar; some significant differences were observed in the kind of symptoms and in the capacity to infect some discriminating plant species (Table 2). All nine isolates
under study systemically infected Brassica napus cv. Otsubu (oilseed rape), B. oleracea var. botrytis (cauliflower), B. rapa ssp. pekinensis cvs. Kyoto-1go and Kyoto-3go (Chinese cabbage), B. rapa var. chinensis cvs. Choyo (Qing geng cai) and Tatsuai (Rosette pakchoi), Eruca sativa (rocket salad), Raphanus sativus cvs. Akimasari (Japanese radish) and Karaj-12 (small radish), Rapistrum rugosum (rugose rapistrum) and Sisymbrium loeselii (small tumble-mustard) and induced chlorotic/necrotic local lesions and systemic symptoms on B. rapa cv. Hakatasuwari (turnip), B. juncea cv. Hakarashina (mustard) and Hirschfeldia incana (shortpod mustard). On B. rapa cv. Karaj-01 (turnip) these isolates induced chlorotic/necrotic lesions 6-15 days after inoculation. Systemic symptoms appeared initially as vein clearing, evolving, according to the isolates, in mosaic, rugosity, leaf deformation and stunting. Only Ca-CE1, Ca-CKh19, Ca-CQ50 and Ca-CSh1 isolates were able to infect kohlrabi plants (B. oleracea var. gongylodes). Contrarily to Ca-CSh1, which infected only locally this indicator plant, the other three isolates induced systemic vein clearing 2-3 weeks after inoculation. Among the studied isolates, only Ca-CKh32, Ca-CQ50, Ca-CSh1 and Ca-CT22 infected Datura stramonium (Jimpson-weed datura). On this host Ca-CKh32, Ca-CSh1 and Ca-CT22 isolates induced only necrotic local lesions, whilst Ca-CQ50 caused also systemic vein clearing and mosaic (Table 2).
Aphid transmission: All CaMV isolates under study were transmitted by Myzus persicae on B. rapa plantlets, which showed vein clearing, leaf deformation and mosaic symptoms ca. 16 days after inoculation. The presence of CaMV in the inoculated plants was confirmed by ELISA.
PCR amplification, sequencing and sequence analysis: In PCR, DNA fragments of approximately 1,580 bp were amplified from all the isolates under study using the primer pair CaMV-6-F and CaMV-6-R. No amplicons were obtained from healthy turnip leaf tissues (Fig. 1). The nucleotide sequences of the amplified fragments for the Iranian CaMV isolates were determined and submitted to the GenBank, with the following accession numbers: DQ119040, DQ119041, DQ870907, DQ870908, DQ870910, DQ870912, DQ870914, DQ870915 and EF503597 (Table 1). BLAST search results confirmed that they contained the complete sequence of the ORF VI, apart from short fragments in the 3' and 5' ends. Multiple nucleotide sequences alignment using the CLUSTAL W program showed 96.9-100% identity among the ORF VI genes of the nine isolates under study. When compared with other non-Iranian CaMV isolates, the Iranian isolates showed the highest and lowest level of identity to D/H (96.1-96.7%) and XinJing (83.7-84.4%) isolates, respectively.
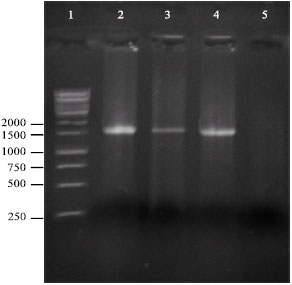 | |
| Fig. 1: | CaMV detection by PCR using CaMV-ORF VI specific primers. Lane 1, DNA marker (GeneRulerTM 1 kbp DNA Ladder, Farmentas, Lithuania); lane 2-4, positive cauliflower samples (amplification of approximately 1,580 bp); lane 5, negative control |
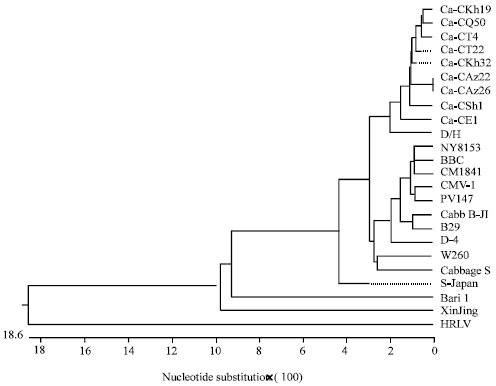 | |
| Fig. 2: | Phylogenetic tree constructed by CLUSTAL W program showing the genetic relationship among ORF VI gene sequences of Iranian and other CaMV isolates |
In the phylogenetic tree constructed from the nucleotide sequences of the CaMV isolates, all Iranian isolates clustered in the same branch, whereas all North-American isolates as well as B29 clustered in a separate branch. Also, the isolates Cabbage S and W260, S-Japan, Bari 1 and XinJing were located in separate clusters (Fig. 2). For the same ORF, at the amino acid level, there was 96.1-100% identity among Iranian isolates. The deduced amino acid sequences of the Iranian CaMV isolates had the highest and lowest identity to D/H (94.5-95.9%) and XinJing (81.3-82.3%) isolates, respectively.
Restriction Fragment Length Polymorphism (RFLP) profiles of the sequences under study (Table 1 and 3) were assessed using Mapdraw program.
 | |
| Fig. 3: | Comparison of Iranian Cauliflower mosaic virus (CaMV) isolates with others from different countries on the basis of restriction fragment length polymorphism (RFLP) pattern using Hpy99I in Mapdraw program. (a) RFLP profile. The size of restricted fragments (bp) is given between restriction sites. (b) Placement of the isolates in different RFLP groups |
The results showed that with Hpy99I eight RFLP groups containing from 0 to 3 restriction sites were created. All nine Iranian isolates were placed separately in RFLP group VIII with three restriction sites at positions 408, 771 and 1458. The majority of North-American isolates together with B29 and W260 isolates were located in RFLP group I, whilst Bari 1, Cabb B-JI, Cabbage S, D/H, S-Japan and XinJing isolates were placed individually in RFLP groups II to VII, respectively (Fig. 3).
DISCUSSION
In this study, CaMV isolates from symptomatic cauliflower plants from different Iranian provinces were selected and analyzed for their biological and molecular properties. Host range studies showed that the isolates under study differed for their reactions especially on turnip, kohlrabi and Datura plants, as reported previously for other isolates (Lung and Pirone, 1972; Schoelz et al., 1986; Al-Kaff and Covey, 1995). Based on the symptoms expressed on kohlrabi seedlings (B. oleracea var. gongylodes) the isolates could be separated into three main groups, thus confirming previous reports (Al-Kaff and Covey, 1995). All isolates were able to infect systemically a wide range of cruciferous hosts tested. Some of the isolates infected not only cruciferous plants but included species of the family Solanaceae (D. stramonium), on which they induced local or systemic symptoms, as described previously (Lung and Pirone, 1972; Garcia and Shepherd, 1985; Schoelz et al., 1986; Schoelz and Shepherd, 1988; Qiu and Schoelz, 1992). All isolates studied in this research were aphid transmissible.
The ORF VI nucleotide sequences of the Iranian CaMV isolates were compared with those of other CaMV isolates available in GenBank. The supposed division of Iranian isolates in two major groups according to their different capacity of infecting cruciferous and solanaceous species did not find a support in molecular comparative analysis of ORF VI genes, confirming previous results (Wintermantel et al., 1993; Pique et al., 1995). The Iranian isolates shared close identity to a Hungarian isolate from B. oleracea (D/H). The phylogenetic analysis among different CaMV isolates based on ORF VI nucleotide sequences did not reveal branching pattern related to the host plant source or aphid transmissibility, as previously reported by Chenault and Melcher (1994b). As indicated by the same authors, the geographic distribution of CaMV sources seems to be the major factor contributing in the evolutionary grouping of this viral species, as the results obtained in this study seem to confirm. The only exception is for B29 isolate from France that clustered with the North-American isolates. The Iranian isolates are more closely related to the Hungarian isolate D/H of CaMV, hence they should be categorized in the non-North American isolates group described previously (Chenault and Melcher, 1994b). RFLP studies performed on the ORF VI gene sequences revealed considerable polymorphism between Iranian and other CaMV isolates. Based on the results obtained, all Iranian isolates were located in a separate RFLP group using Hpy99I. Interestingly, the grouping pattern obtained by using Hpy99I strictly agreed with those obtained in phylogenetic studies of ORF VI sequences.
The present study reports cloning, sequencing and comparison of CaMV ORF VI gene of Iranian CaMV isolates for the first time. The phylogenetic results in agreement with restriction analysis revealed that the Iranian isolates can be assigned to a well separated group. This virus was probably successfully introduced in Iran through plant material transportations and diverged in an independent evolutionary way. Further investigations on molecular properties of CaMV isolates in Iran are necessary to confirm this hypothesis.
ACKNOWLEDGMENTS
This study was supported by the Plant Virology Department of PPDRI (Tehran) and Plant Protection Department of Tehran University. We are grateful to Science and Research Campus of Islamic Azad University for its helps. The authors also wish to thank Dr. U. Melcher (Department of Biochemistry and Molecular Biology, Oklahoma State University) for designing primers and to Dr. M. Digiaro (Istituto Agronomico Mediterraneo, Valenzano, Italy) for his kindly comments in this research. This study was part of Ph.D thesis of the first author.
REFERENCES
- Agama, K., J. Beach, J. Schoelz and S.M. Leisner, 2002. The 5 third of Cauliflower mosaic virus gene VI conditions resistance breakage in Arabidopsis Ecotype Tsu-0. Phytopathology, 92: 190-196.
PubMedDirect Link - Al-Kaff, N.S. and S.N. Covey, 1995. Biological diversity of Cauliflower mosaic virus isolates expressed in two Brassica species. Plant Pathol., 44: 516-526.
Direct Link - Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Anderson, E.J., S.G. Qiu and J.E. Schoelz, 1991. Genetic analysis of determinants of disease severity and virus concentration in Cauliflower mosaic virus. Virology, 181: 647-655.
PubMedDirect Link - Burland, T.G., 2000. DNASTARs lasergene sequence analysis software. Meth. Mol. Biol., 132: 71-91.
PubMedDirect Link - Chenault, K.D. and U. Melcher, 1994. Phylogenetic relationships reveal recombination among isolates of Cauliflower mosaic virus. J. Mol. Evol., 39: 496-505.
CrossRefDirect Link - Cheng, R.H., N.H. Olson and T.S. Baker, 1992. Cauliflower mosaic virus: A 420 subunit (T = 7), multilayer structure. Virology, 186: 655-668.
CrossRef - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Daubert, S.D., J. Schoelz, L. Debao and R.J. Shepherd, 1984. Expression of disease symptoms in Cauliflower mosaic virus genomic hybrids. J. Mol. Applied Genet., 2: 537-547.
PubMedDirect Link - Daubert, S. and G. Routh, 1990. Point mutations in Cauliflower mosaic virus gene VI confer host specific symptom changes. Mol. Plant Mic. Interac., 3: 341-345.
Direct Link - Farzadfar, S.H., R. Pourrahim, A.R. Golnaraghi and A. Ahoonmanesh, 2005. Occurrence of Cauliflower mosaic virus in different cruciferous plants in Iran. Plant Pathol., 54: 810-810.
Direct Link - Farzadfar, S.H., A. Ahoonmanesh, G.H. Mosahebi, R. Pourrahim and A.R. Golnaraghi, 2007. Occurrence and distribution of Cauliflower virus on cruciferous plants in Iran. Plant Pathol. J., 6: 22-29.
Direct Link - Garcia, O. and R.J. Shepherd, 1985. Cauliflower mosaic virus in the nucleus of Nicotiana. Virology, 146: 141-145.
CrossRef - Gray, S.M. and N. Banerjee, 1999. Mechanisms of arthropod transmission of plant and animal viruses. Microbiol. Mol. Biol. Rev., 63: 128-148.
PubMedDirect Link - Haas, M., B. Marin, A. Gelderich, P. Yot and M. Keller, 2002. Cauliflower mosaic virus: Still in the news. Mol. Plant Pathol., 3: 419-429.
CrossRefDirect Link - Lung, M.C.Y. and T.P. Pirone, 1973. Studies on the reason for differential transmissibility of Cauliflower mosaic virus isolates by aphids. Phytopathology, 63: 910-914.
CrossRefDirect Link - Mason, W.S., J.M. Taylor and R. Hull, 1987. Retroid virus genome replication. Adv. Virus Res., 32: 35-96.
PubMedDirect Link - Palacios, I., M. Drucker, S. Blanc, S. Leite and A. Moreno, 2002. Cauliflower mosaic virus is preferentially acquired from the phloem by its aphid vectors. J. Genet. Virol., 83: 3163-3171.
Direct Link - Pique, M., J.L. Mougeot, A. Geldreich, T. Guidasci, J.M. Mesnard and G. Lebeurier, R. Yot, 1995. Sequence of a Cauliflower mosaic virus strain infecting solanaceous plants. Gene, 155: 305-306.
CrossRef - Pirone, T. and S. Blanc, 1996. Helper dependent vector transmission of plant viruses. Ann. Rev. Phytopathol., 34: 227-247.
CrossRefDirect Link - Qiu, S.G. and J.E. Schoelz, 1992. Three regions of Cauliflower mosaic virus strain W260 are involved in systemic infection of solanaceous hosts. Virology, 190: 773-782.
PubMedDirect Link - Qiu, S.G., W.M. Wintermantel, Y. Sha and J.E. Schoelz, 1997. Light dependent systemic infection of solanaceous species by Cauliflower mosaic virus can be conditioned by a viral gene encoding an aphid transmission factor. Virology, 227: 180-188.
CrossRef - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sanger, M., S. Daubert and R.M. Goodman, 1991. The regions of sequence variation in caulimovirus gene VI. Virology, 182: 830-834.
CrossRef - Schoelz, J.E. and R.J. Shepherd, 1988. Host range control of Cauliflower mosaic virus. Virology, 162: 30-37.
CrossRefPubMedDirect Link - Schoelz, J.E., R.J. Shepherd and R.D. Richins, 1986. Properties of an unusual strain of Cauliflower mosaic virus. Phytopathology, 76: 451-454.
CrossRefDirect Link - Shahraeen, N., S.H. Farzadfar and D.E. Lesemann, 2003. Incidence of viruses infecting winter oilseed rape (Brassica napus ssp. oleifera) in Iran. J. Phytopathol., 151: 614-616.
Direct Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Wintermantel, W.M., E.J. Anderson and J.E. Schoelz, 1993. Identification of domains within gene VI of Cauliflower mosaic virus that influence systemic infection of Nicotiana bigelovii in a light-dependent manner. Virology, 196: 789-798.
PubMedDirect Link