
M. Maziah
Department of Biochemistry, Faculty of Biotechnology and Molecular Sciences, Malaysia
M. Sariah
Department of Plant Protection, Faculty of Agriculture, University Putra Malaysia,
43400, Serdang, Selangor Darul Ehsan, Malaysia
S. Sreeramanan
Department of Biotechnology, Asian Institute of Medicine,
Science and Technology (AIMST), Amanjaya, 08000, Sungai Petani, Kedah
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 271-282







ABSTRACT
A soybean endo β-1,3-glucanase gene (pROKla-Eg) multiplied in Agrobacterium strain LBA 4404, was simultaneously introduced into single buds of in vitro grown banana cultivar, Rastali (AAB). Plasmid pROKla-Eg contained a neomycin phosphotransferase gene (npt 11) as the selectable marker to identify the transformants. Treatment A contained kanamycin at 100 mg L-1 and treatment B contained geneticin G-418 at 50 mg L-1 in both MS medium supplemented 5 mg L-1 of BAP together with 2.7 g of gelrite agar. Single buds derived from multiple bud clumps (Mbcs), were the target explants for transformation. An assay was performed to identify the minimum concentration required for two antibiotics (carbenicillin and cefotaxime) that is most effective against Agrobacterium strains, LBA 4404 and the effect on tissue regeneration capacity. Even though the transformation frequency based on kanamycin selection medium (treatment A) is higher, but there is no transformant could be confirmed based on PCR and Southern blot analyses, as compared using geneticin G-418 selection medium (treatment B). These results suggested that using geneticin G-418 as selection agent is preferably than kanamycin due to lower concentration required to allow for the small numbers of putative transgenic cells in a large population of non-transformed ones to undergo multiplication and also reduced the occurrence of escapes. The transgenic banana plantlets were inoculated with 2x106 spores mL-1 conidial suspension of Fusarium oxysporum f. sp. cubense (Race 1) to evaluate the degree of tolerance and to investigate the effectiveness of the bioassay system as a potential tool for early screening. Assay of protein extract from the transgenic plantlets showed an significantly increased in β-1,3-glucanase enzymes activity over the untransformed plantlets. The present of Agrobacterium-mediated transformation reported here is suitable for using tiny meristem tissues to obtain fungal disease tolerant or resistant banana through genetic engineering.
PDF Abstract XML References
How to cite this article
M. Maziah, M. Sariah and S. Sreeramanan, 2007. Transgenic Banana Rastali (AAB) with β-1, 3-glucanase Gene for Tolerance to Fusarium Wilt Race 1 Disease via Agrobacterium-mediated Transformation System. Plant Pathology Journal, 6: 271-282.
DOI: 10.3923/ppj.2007.271.282
URL: https://scialert.net/abstract/?doi=ppj.2007.271.282
DOI: 10.3923/ppj.2007.271.282
URL: https://scialert.net/abstract/?doi=ppj.2007.271.282
INTRODUCTION
In the past, Musa species were considered outside the host range of Agrobacterium, but an increasing amount of research reports indicates that banana can be transferred by Agrobacterium tumefaciens (May et al., 1995; Ganapathi et al., 2001; Khanna et al., 2004; Sreeramanan et al., 2006). The recovery of transgenic plants of banana obtained by means of Agrobacterium tumefaciens-mediated transformation has been described precisely. May et al. (1995) reported transformation of meristems and corm slices from cultivar Grand naine (AAA). Ganapathi et al. (2001) successfully transformed Rastali (AAB) by using embryogenic cell suspensions as a starting material. A total of two hundred putative transformants were recovered, of which a set of 16 was tested by histochemical analysis for GUS expression and by Southern blot analysis. Therefore, the controversy over susceptibility of monocotyledonous plants for transformation using Agrobacterium tumefaciens appears to be solved for a continuously increasing number of plant species.
This study describes a series of experiments that carried out using LBA 4404 which contained plasmid with β-1,3-glucanase gene (pROKla-Eg). Comparison between using kanamycin and geneticin G-418 as selection agents were demonstrated in this study since both plasmid contained npt11 gene. The inhibition of explants and bacterial growth by carbenicillin and cefotaxime were demonstrated by using different concentrations.
Generally, it is known that plants have defense systems which involve pathogenesis-related proteins such as β-1,3-glucanase (Nakamura et al., 1999). β-1,3-glucanase catalyzes the hydrolysis of β-1, 4 linkages of the N-acetyl-D-glucosamine polymer β-1, 3-glucan, which is a component of mycelial cell walls. Nakamura et al. (1999) reported that transgenic kiwi fruit engineered with β-1, 3-glucanase, shown increased resistance against Botrytis cinerea. Similarly, transgenic potato expressing soybean β-1,3-endoglucanase gene exhibit an increased resistance to Phytophthora infestans (Borkowska et al., 1998). Recently, we have reported development of bioassay method for testing Fusarium wilt disease tolerance in transgenic banana by relating disease severity to level of production of various plant compounds and enzymes (Sreeramanan et al., 2006).
For the analyses of transgene integration patterns, a Polymerase Chain Reactions (PCR) and Southern blot hybridization techniques have been elaborated and evaluated. Here, we report that an efficient transformation frequency in banana cultivar, Rastali (AAB) was obtained using Agrobacterium tumefaciens, LBA 4404 with β-1,3-glucanase (Eg) and neomycin phosphotransferase (npt 11) genes.
MATERIALS AND METHODS
Plant materials: Corm slices of in vitro banana cultivar, Rastali (AAB) plantlets were cultured in MS medium (Murashige and Skoog, 1962) supplemented with 10 mg L-1 of 6-Benzylaminopurine (BAP) to obtain multiple bud clumps. The cultures were incubated at 25±2°C and a 16 h photoperiod with cool white fluorescent light of 150 μmol m-2 sec-1 (supplied by Philips TLD fluorescent light tubes). Single buds (3 mm), excised from multiple bud clumps, were used for this study.
Bacterial strain and binary vector
LBA 4404 (pROKla-Eg): Disarmed Agrobacterium tumefaciens strain LBA 4404 (Hoekema et al., 1983) harboring a binary vector pROKla-Eg (Yoshikawa et al., 1993) which originated in BIN 19 was used. The plasmid pROKla-Eg contains the npt 11 gene linked to the nopaline synthase gene (nos) promoter and the soybean β-1,3-endoglucanase cDNA linked to the CaMV 35S (35S) promoter in the T-DNA region (Fig. 1).
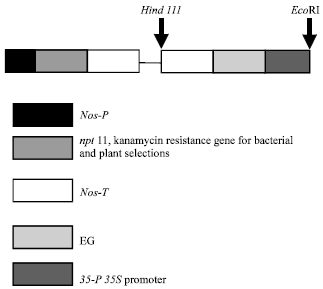 | |
| Fig. 1: | T-DNA region of pROKla-EG (RB: Right border, LB: Left border, Nos-P: Nopaline synthase gene promoter; npt 11: Neomycin phosphotransferase gene, Nos-T: Nopaline synthase gene terminator, 35-P: 35S promoter, EG: Soybean β-1,3-endoglucanse cDNA) |
Bacterial strain and culture conditions: Agrobacterium tumefaciens, LBA 4404 (pROKla-Eg) were streaked out on LB medium supplemented with 50 mg L-1 of kanamycin and grown at 28°C for colonies to form. Single colony was transferred to 20 mL LB liquid contained the same antibiotic and the culture was allowed to shake at 28°C and 120 rpm for 20 h. After adjusting the Optical Density to 0.6 units at 600 nm (OD600), the bacterium culture was used for transformation.
Explant and bacterial toxic effects of carbenicillin and cefotaxime: For explant toxicity tests, single buds of 3 mm in size were transferred to MS medium contain 5 mg L-1 of BAP and different concentrations of carbenicillin and cefotaxime (0, 100, 200, 300, 400, 500 and 600 mg L-1). After 30 min co-cultivation of explants in bacterial cultures, the percentage of single buds with Agrobacterium strains were determined after three days using various carbenicillin and cefotaxime concentrations.
Transformation protocols of single meristematic buds: Single buds (∼3 mm) were precultured for three days prior to Agrobacterium infection. The explants injured mildly using scalpel. About 100 single buds immersed in Agrobacterium suspension for 30 min together with an optimized acetosyringone concentration at 100 μM. Acetosyringone is known to activate the virulence genes of the Ti plasmid and to initiate the transfer of the T-DNA. The explants were then blotted dry on sterile filter paper and cocultivated for three days on hormone free MS medium (without CaCl2 during co cultivation) designated as M1 medium containing 100 μM of acetosyringone, 60 mM D-(+)-glucose, 2 mM sodium phosphate and 40 mg L-1 of L-Cysteine using Petri dish at 22°C temperature in the dark condition. After co-cultivation period for three days, the buds were transferred on same medium but without acetosyringone concentration together with 300 mg L-1 of carbenicillin or 200 mg L-1 cefotaxime designated as M2 medium for 5 days and transferred back on hormone free MS liquid medium with the same antibiotic concentrations for another 5 days. Then, the buds were transferred to MS solid medium containing 10 mg L-1 of BAP with 100 mg L-1 of kanamycin or 50 mg L-1 of geneticin G-418 for 2 weeks and transferred again on liquid medium with the same treatments for 5 days in 100 mL conical flasks. During this period, death buds were removed before subcultured back on MS solid medium containing 10 mg L-1 of BAP with 25 mg L-1 of geneticin G-418. The transformed buds regenerated into clumps which are known as multiple bud clumps (Mbcs). Single buds were separated from mbcs and subcultured onto solid MS medium with 1 mg L-1 of BAP until the single plants were regenerated. All plants regenerated from each putatively independent transformed bud line were maintained under in vitro conditions.
Total genomic DNA extraction: Genomic DNA was extracted from putative transformants using an improved and modified CTAB method adopted from Pasakinskiene and Paplauskience (1999).
Polymerase Chain Reactions (PCR): PCR was done using the DNA Thermal Cycler 480 machine (Perkin-Elmer). β-1,3- glucanase (pROKla-Eg) and npt 11 genes were amplified using standard PCR protocols (Sambrook et al., 1989). The following primers were used to amplify the transgenes and part of the regulatory sequences (Table 1).
PCR amplications were carried out in 100 μL reactions volume containing template DNA (500 ng genomic DNA or 60 ng plasmid DNA), 200 ng of each primer (forward and reverse), 0.2 mM dNTP mix, 1.5 mM MgCl2, 1X PCR buffer and 5 U Taq DNA polymerase (MBI Fermentas). Amplification of β-1,3-glucanase gene (Eg) fragments was performed for 45 cycles at 94°C for 1 min, 40°C for 2 min and 72°C for 3 min, for denaturing, annealing and primer extention, followed by 72°C for 7 min. Amplication of npt11 gene fragments was performed for 35 cycles at 94°C for 1 min, 62°C for 1 min and 72°C for 2 min, for denaturing, annealing and primer extention, respectively. All amplified PCR products were checked on 1.2% agarose gel.
Southern blot analyses: Genomic DNA (20 μg) from PCR positive transformants were subjected to digestion with Hind111 in buffer H (Promega). Digestions were carried out for 2 h at 37°C in 20 μL final volumes. DNA fragments were separated by electrophoresis through a 1.0% agarose gel. A nonradioactive method was used to confirm stable integration of β-1,-3 glucanase (Eg) transgenes in the host banana genome. DIG DNA Labeling and Detection Kit (Roche) were used in this study.
Development of fusarium bioassay method: Development of Fusarium bioassay method in transgenic banana in this study was referred using an improved method adopted by our recent published study (Sreeramanan et al., 2006). For plant inoculation bioassay, untransformed banana cultivar, Rastali (AAB) plantlets roots sections were cut at 3 cm from the root tips. The plants dipped into spore concentrations of 106 for one hour and were then transplanted in a plastic container (8x8 cm) with sterile perlite. One milliliter of 106 spore concentrations was again inoculated near the root/stem regions for double confirmation. The total spores used were 2x106 spores mL-1). The plantlets were watered using hormone and sucrose free MS liquid medium. Disease development and severity were monitored and measured over for a period of four weeks. Necrosis in new and old leaves tissue (yellowing) indicated Fusarium wilt symptoms occurred.
Extraction and determination of total acid soluble protein and β-1,3-glucanase activity: Tissue samples (0.25 g) were homogenised in 5 mL extraction buffer (0.1 M sodium buffer, 1 mM EDTA, 0.1% mercaptoethanol, pH 5.2), with 100 mg insoluble polyvynil polypyrroline. Protein was quantified by Bradford (1976) methods, at wavelength 595 nm. The β-1,3-glucanase activity assay methods of Tonon et al. (1998) were followed with a slight modification. β-1,3-glucanase activity determination was based on the rate of reducing sugar production using Laminarin (Sigma) as the substrate.
| Table 1: | Genes, primer, primer sequences and expected product length from PCR |
 | |
Amount of enzyme catalyzing the formation of 1 nm glucose equivalent in one second under assay conditions refers as 1 nkat (Anfoka and Buchenauer, 1997).
Statistical analysis: Data were analysed using one-way ANOVA and the differences contrasted using Duncan’s multiple range test. All statistical analyses were performed at 5% using SPSS 10.0 (SPSS Inc. USA).
RESULTS AND DISCUSSION
Toxicity of carbenicillin and cefotaxime on single buds and agrobacterium growth: For effective Agrobacterium-mediated transformation, the antibiotic regime should control bacterial growth without inhibiting the regeneration of the plant cells. The effect of carbenicillin and cefotaxime on the percentage of single buds growth, were evaluated after four weeks of culturing on the solid and liquid medium containing an appropriate concentrations (Fig. 2 and 3). No significantly differences were observed between the cultures when the percentage number of single buds growth was counted.
However, using 600 mg L-1 cefotaxime in solid medium reduced average growth from 100% (control) to 30%. The percentage of growth in liquid medium using same concentration is lower 12% than solid medium. In comparison to carbenicillin, using 600 mg L-1 reduced average growth from 62% in solid medium and 54% in liquid medium. The percentage of growth is reduced to 8 and 12% in both solid and liquid containing 150 mg L-1 carbenicillin. However in cefotaxime, percentage of growth dropped tremendously to 82% in solid medium and 63% in liquid medium using the same concentration of 600 mg L-1.
The Agrobacterium tumefaciens, strain LBA 4404 consistently sensitive to the antibiotics tested. Using carbenicillin, the inhibition of cell growth started at a concentration of 100 mg L-1 but complete inhibition obtained using 400 mg L-1 for LBA 4404 (Fig. 4). For cefotaxime, the inhibition of cell growth started at 100 mg L-1 and complete inhibition of cell growth at 200 mg L-1 for both Agrobacterium strains (Fig. 4). The results demonstrated that, Agrobacterium cells were more sensitive to cefotaxime than carbenicillin.
Previous studies had shown that a higher concentration of Agrobacterium cells significantly increases the transformation efficiency of tobacco and Arabidopsis thaliana (Lin et al., 1994). Therefore, for transformation with high concentrations of Agrobacterium, LBA 4404 is the preferred strain since it is easier to eliminate from the infected explants after co-cultivation period compared to other strains.
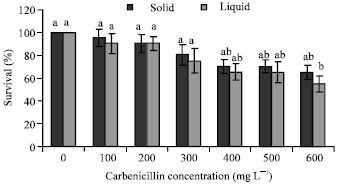 | |
| Fig. 2: | Percentage of single buds growth obtained after 4 weeks in MS media using different carbenicillin concentration. Forty single buds of 3 mm in size were plated onto 5 mg L-1 of BAP containing antibiotic in various concentrations in both solid and liquid media. The data plotted were the means of four replicates. Data were analysed using one-way ANOVA and the differences contrasted using Duncan’s multiple range test. Different letters indicate values are significantly different (p<0.05) |
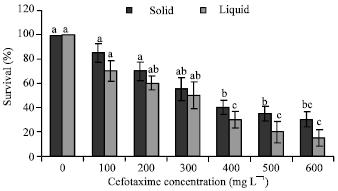 | |
| Fig. 3: | Percentage of single buds growth obtained after 4 weeks in MS media using different cefotaxime concentration. Forty single buds of 3 mm in size were plated onto 5 mg L-1 of BAP containing antibiotic in various concentrations in both solid and liquid media. The data plotted were the means of four replicates. Data were analysed using one-way ANOVA and the differences contrasted using Duncan’s multiple range test. Different letters indicate values are significantly different (p<0.05) |
Although cefotaxime is more effective in inhibiting Agrobacterium cell growths, carbenicillin is the antibiotic of choice in Agrobacterium-mediated transformation because cefotaxime has shown high toxicity in many different plant tissues (Antunez de Mayolo et al., 2003). In cacao, the addition of the cefotaxime decreased somatic embryo production by 86% (Antunez de Mayolo et al., 2003). However, it has been reported that cefotaxime did not show negative effect in apple tissues (Maximova et al., 1998), adventitious buds and shoots induction in Pinus radiate (Holland et al., 1997) and chrysanthemum (Teixeira da Silva and Fukai, 2002). The other antibiotic, augmentin, timentin and moxalactam were also used after co-cultivation to kill the Agrobacterium as it was not reduce regeneration capacity (Park and Facchini, 2000; Antunez de Mayolo et al., 2003).
Therefore, different concentration of carbenicillin and cefotaxime required in solid and liquid medium of banana cultivar, Rastali (AAB) explants in order to avoid toxicity or overgrowth of Agrobacterium tumefaciens which could inhibit the explants growth and caused contamination which could reduces transformation efficiency.
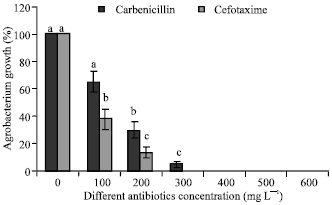 | |
| Fig. 4: | Percentage of single buds with Agrobacterium growth using with different carbenicillin and cefotaxime concentrations. Forty single buds of 3 mm in size were plated onto 5 mg L-1 of BAP containing antibiotic in various concentrations in both solid and liquid media. The data plotted were the means of four replicates. Data were analysed using one-way ANOVA and the differences contrasted using Duncan’s multiple range test. Different letters indicate values are significantly different (p<0.05) |
Transformation of single meristematic buds and plant regeneration: Single buds of banana cultivar, Rastali (AAB) were infected and co-cultivated with induced Agrobacterium strains harboring different binary vectors, following the condition described in materials and methods. However, LBA 4404 (pROKLa-Eg) did not consist any of gusA or gfp genes as a visual marker gene. This strain was used as a vector for transformation system using two different treatments which designed as treatment A (selection using kanamycin) and treatment B (selection using geneticin G-418).
Co-cultivated single buds were selected on MS medium consisting of 10 mg L-1 of BAP containing kanamycin at 100 mg L-1 (treatment A) or geneticin G-418 (treatment B) at 50 mg L-1, during a period of 2 to 3 months, depending on the experiments. Selection medium containing geneticin G-418 produces lower number of buds resistant to this antibiotic. However, none of the negative control plates did ever give rise to surviving cell aggregates under any of the selective conditions used in this experiment. In the absence of antibiotics, uninfected single buds proliferated normally on 10 mg L-1 of BAP, whereas they turned to black and died after two months under selection condition even at the lower concentration tested.
Percentage of transformation frequency were evaluated based on the number of explants regenerated (Table 2 and 3). Transformation frequency from Treatment A is higher than Treatment B. More than 30 cell clusters arose per individual co-cultivated sample after selection with many of which initiated the formation of multiple bud clumps, while the rest of untransformed cells which did not stand selection, blackened and died (Fig. 5).
After first level of selections, the remaining proliferation buds were subcultured into liquid medium containing 10 mg L-1 of BAP using the same concentration of antibiotics that had been used previously in first selections. After one week, the remaining healthy proliferating buds then selected in same concentration of selections in solid MS medium. Multiple bud clumps aggregates formed during the selective process were regenerated in BAP and antibiotics free medium.
| Table 2: | Comparison of transformation frequency of Agrobacterium tumefaciens, EHA 101 (pBI333-EN4-RCC2) using Treatment A (Selection using kanamycin) |
 | |
| aContain kanamycin (100 mg L-1), bContain kanamycin (70 mg L-1), cMS medium without BAP, dTransformation frequency (%) = No. of explants regenerated in hormone free medium (e)/total explants inoculated with Agrobacterium (a)x100% | |
| Table 3: | Comparison of transformation frequency of Agrobacterium tumefaciens, LBA 4404 (pROKla-EG) using Treatment B (Selection using Geneticin G-418) |
 | |
| aContain geneticin (50 mg L-1), bContain geneticin (25 mg L-1), cMS medium without BAP, dTransformation frequency (%) = No. of explants regenerated in hormone free medium (e) / total explants inoculated with Agrobacterium (a) x 100 | |
 | |
| Fig. 5: | Production of transgenic Pisang Rastali. (A) Two months in geneticin G-418 selection media, (B) Some single bud develop into green multiple bud clumps in 5 mg L-1 of BAP concentration, (C) Multiple shoots proliferating and (D) Putative transgenic plantlet confirmed from PCR and Southern Blot analyses |
Table 2 and 3 showed the results of three independent transformation experiments for each LBA 4404 (pROKla-Eg) treatment using two different antibiotic selections. Single plantlets arising from geneticin G-418 resistant cell clumps in hormone free (without BAP) formed shoots and root tips without any necrosis been observed. The observation patterns revealed that the tissue were fully transgenic without evidence of chimaeras. However, this is not true for explants regenerated in vitro from treatment A, which kanamycin were applied as a selection marker. Some of transgenic plantlets produced albinos and semi-necrosis growth. This could be due to kanamycin selection, which caused production of phenotypically abnormal plants confirmed observations in peas (Bean et al., 1997; Nadolska-Orcyzk and Orczyk, 2000). Similarly, kanamycin effect has been reported in transgenic papaya by Yu et al. (2003). In plant cells, kanamycin exerts its effect on mitochondria and chloroplasts by impairing protein synthesis, resulting chlorosis (Weide et al., 1989). For several Malus cultivars and rootstocks, kanamycin at 50 mg L-1 is pyhtotoxic and causes shoot chlorosis and necrosis (Yepes and Aldwinckle, 1994).
For transforming papaya with Agrobacterium, 75-150 mg L-1 kanamycin was used to kill non-transformed cell but with low efficiency in transformation and many abnormal transformed somatic embryos are noticed during the selection process (Yang et al., 1996). Chimeras plantlets developing delay necrosis effect due to kanamycin responses during proliferation of multiple bud clumps and regeneration into a single plantlets. The average frequency of regenerated plantlets raised to 10% when geneticin G-418 was used during third phase of selections indicating that generation of escapes was completely avoided for both transformation using Agrobacterium strain, LBA 4404.
Nevertheless, maintenance of cultures for long periods and with high antibiotics concentration for more than three phase selection resulted in reduced numbers of surviving cell aggregates. Irrespectively of the selective conditions used, average frequencies of plant regeneration ranged from 5 to 20% putatively independent lines per co-cultivation in all samples. This study demonstrated that the use of Agrobacterium tumefaciens as a transformation system for banana cultivar, Rastali (AAB) could be highly efficient when a rapid screening system for the identification of lines of interest developed before performing more detailed molecular analyses. Techniques based on the Polymerase Chain Reactions (PCR) are the best option for analyzing large amount amounts of transformants, since they are fast and demand low quantities of genomic DNA.
The use of highly regenerable single meristematic buds of banana cultivar, Rastali (AAB) in combination with Agrobacterium as a vector for DNA transfer has been not described elsewhere previously. However using other banana meristem tissue such as corm slices in Agrobacterium-mediated transformation may be limited application because of the risk of generating chimaeric plants even though the transformation frequency obtained with particle bombardment could be markedly improved (May et al., 1995). Besides being more efficient, Agrobacterium-mediated transformation is technically simpler than particle bombardment, only requiring basic microbiology facilities and generally results in high levels of expression due to a simple integration pattern of well defined DNA sequences into transcriptionally active regions of the plant genome, which make it the best option when both transformation systems are available (Hiei et al., 1997; Cheng et al., 1997).
Analysis of T-DNA integration via PCR ampflication: Though Agrobacterium-mediated transformation is the most resorted method for the generation of transgenic plants with single integration of a precisely delimited DNA sequences (Smith and Hood, 1995; Lawrence and Koundal, 2001), the structure of the inserted T-DNA varies widely to include single or multiple copies, individual or tandem repeats, at a unique or several loci in the plant genome (Iglesias et al., 1997).
Total DNA isolated from the putative transformants was tested for the presence of the transgenes. The efficient and simplicity of the PCR analysis, enable screening for transformed plants in a shorter period. PCR amplification confirmed that the β-1,3-glucanase (Eg) gene and npt11 gene were present in a high proportion of the geneticin G-418 resistant regenerants (treatment B) compared to kanamycin selection medium (treatment A). The PCR results of some transgenic plantlets are displayed in Fig. 6 and 7.
In all the experiments, no bands could be detected from DNA extracted from untransformed control. Fourteen DNA samples were picked from kanamycin selection (treatment A) and six from geneticin G-418 selection (treatment B) were subjected for PCR analyses from transformation using Agrobacterium tumefacien strain, LBA 4404 (pROKla-Eg).
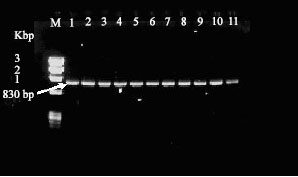 | |
| Fig. 6: | PCR analysis of β-1,3-endoglucanse gene in transgenic banana plantlets selected on geneticin G-418. Lane M = Lambda DNA digested with Hind111 was used as molecular weight marker; Lane 1 = Transforming plasmid, pROKla-Eg; Lane 2-11 = Putative transformed plantlets showing the amplified 830 bp β-1,3-endoglucanse gene |
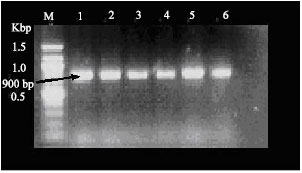 | |
| Fig. 7: | PCR analysis npt11 gene in transgenic banana plantlets selected on geneticin G-418. Lane M = Lambda DNA digested with Hind111 was used as molecular weight marker; Lane 1 = Transforming plasmid, pROKla-Eg; Lane 2-6 = Putative transformed plantlets showing the amplified 900 bp npt11 gene |
No bands (Eg and npt 11 genes) were detected from plants using kanamycin selection (treatment A). These were probably escapes or the kanamycin selection plants and lost the npt 11 gene when the selective pressure was removed or could be chimeric for expression of the npt 11 gene. Similar observations of high escape rates using kanamycin has been observed in oil palm, rice and orchid (Parveez et al., 1996; Kuvshinov et al., 1999; Janna et al., 2001). The glucanase (Eg) gene linked with the selectable marker gene (npt 11) could not detect, respectively.
Two out of the six DNA samples were analyses from Agrobacterium tumefacien strain, LBA 4404 (pROKla-Eg) transformation plantlets, with successfully amplified the expected band size of 830 bp of β-1,3-glucanase gene from geneticin G-418 selection medium (Fig. 6). Co-integration of the npt 11 (900 bp) detect in these two transformants (Fig. 7). Two transgenic plantlets were differentiate according to plant codes [B2E (2) and B3E (31)]. Similar observation in all seven transgenic scented geranium with a gene encoding antimicrobial protein Ace-AmPI were confirmed by PCR that these plants contained the npt 11 selectable marker gene (Bi et al., 1999).
Longer duration of geneticin G-418 in selection medium did not help to reduce the outcomes of chimeric tissue instead it decreases the survival of regenerating plants, appeared to effect fertility of the regenerated plants and surprisingly produced pink colour pigmentation in shoots and leaves. However, these pigmented plantlets return back to normal green plantlet when the selection removed after six cycles and maintained in hormone free MS medium.
The presence of hygromycin or basta during five cycles of oil palm embroyogenic callus clumps remained essential as otherwise some of the regenerated plants were not transgenic (Parveez et al., 1996). The fact that transgenic banana cells were selected during a longer period (four to six months) before regeneration is started probably explains the low number of transformants in geneticin G-418 selection medium. However, no possible explanation was formulated for this low transformation frequency. PCR analysis of transgenic papaya showed the presence of the npt 11 selectable marker gene and the linked transgene (the coat protein gene of the papaya ringspot virus) in 95 and 42% of putative transformants analysed, respectively (Fitch et al., 1992). As the latter transgene was located between the npt 11 and gusA transgene, it appeared that complex rearrangements of transgenes occurred (Fitch et al., 1992).
Few extensive PCR analyses have been conducted to investigate transformation frequencies of transgenes, either linked or non linked in kiwifruit and papaya. Examination by PCR of 26 kanamycin resistant and thus, npt 11 containing, regenerated kiwifruit shoots revealed that 17 (65%) also contained the linked soybean β-1,3-glucanase gene (Nakamura et al., 1999), which is lower than obtained in this study. A slightly higher transformation frequency for linked transgenes of 80% can be calculated based on expression analysis of the gusA transgene and PCR analysis of the coat protein gene of the papaya ringspot virus in 12 papaya transformants (Fitch et al., 1992).
Southern blot hybridization analysis: Southern blot analyses were carried out to evaluate further the transfer and insertion of the β-1,3-glucanase gene in the genome of the transformed banana cultivar, Rastali (AAB). Although two lines of evidence, (i) the antibiotic resistant phenotype and (ii) the presence of the introduced genes in putative transformants as shown indeed by PCR demonstrated that the regenerated plants were transgenic, proof of stable integration of transgenes by Southern analysis was required.
Southern blot analysis also yields information on the number of integration sites per transgene, the copy number of the transgenes and whether transgenes are integrated in the same locus (linked co-integration) or not (non-linked co-integration) (Kohli et al., 1998). Results presented here comprise only the β-1,3-glucanase (Eg) hybridizations. No hybridisation was observed to the control plantlets. Hybridisation to high molecular weight DNA indicated that stable integration of the transgenes into the banana genome.
Even two transgenic plantlets (treatment B) obtained from PCR positive analysis, interestingly 100% were found to contain a single gene insertion in both leaves and roots tissue. Hind111 digestion of transforming plasmid pROKLa-Eg (Fig. 8; Lane 1), released an approximately 3.5 kb size fragment containing the CaMV 35S, Eg and Nos terminator (Fig. 8). No hybridization was observed from non-transformed plantlet (Fig. 8; Lane 2). Lane 3 and 4 are the DNA samples from leaves and roots tissue of plantlet code B2E (2), Lane 5 (leaves), Lane 6 and 7 (roots) of plantlet B3E (31). The presence of bands with sizes larger than the Eg (endoglucanase) fragment (3.5 kb) was indicative of integrative of the introduced plasmid.
 | |
| Fig. 8: | Southern blot analysis of the β-1,3-endoglucanse gene integration pattern in banana plantlets. Digested genomic DNA with Hind 111 using PCR amplified β-1,3-endoglucanse gene (830 bp) as probe. Lane1 = Transforming plasmid, pROKla-Eg; Lane 2 = untransformed control plantlets; Lane 3-7 = genomic of putative transformed plantlets |
 | |
| Fig. 9: | Progress of Disease Symptoms (Secondary) caused by Fusarium oxysporum f.sp. cubense (Race 1) on leave of transgenic banana cultivar, Rastali (AAB) plantlets (A). Control plantlet (No spore inoculated), (B) Control plantlet with (spores inoculated), (C) B2E (2), (D) B3E (31) and (E) Transgenic Rastali (AAB) in the glasshouse |
Patterns of integration, inheritance and expression of transgenes in plant upon Agrobacterium-mediated transformation had a lower copy number of transgenes and a more predictable pattern of integration (Smith and Hood, 1995; Klimaszewska et al., 2003). However, Kononov et al. (1997) reported that approximately 75% of the transgenic tobacco plants from Agrobacterium-mediated transformation contained sequence from the binary vector backbone sequences in plant genome. Furthermore, there was not a clear correlation between transgene expression and transgene copy number (Hobbs et al., 1993). While single copies of transgenes may tend to be more stably expressed than multiple gene copies or scrambled inserts, there were additional factors that influenced transgene expression (Iglesias et al., 1997). The results from Southern blot analyses also rule out the possibility of Agrobacterium contamination of tissue resulting in false-positives.
Development of fusarium bioassay: Progress of the disease symptoms after 4 weeks caused by Fusarium oxysporium f. sp. cubense (race 1) on the leave of the transgenic banana cultivar, Rastali (AAB) plantlets shown in Fig. 9.
Below is the summary of final evaluation obtained for leaf disease symptoms after four weeks inoculation (Fig. 9):
Plant code: B2E (2) contains β-1, 3 glucanase gene (ROKla-Eg): Yellowing most of the older and discolouration of younger leaves appear after 28 days of inoculation.
Plant code: B3E (31) contains β-1, 3 glucanase gene (ROKla-Eg): Slight discolourations appear on one of the younger leaves at the end of 28 days. No yellowing of older leaves could be observed.
Analysis of β-1,3 glucanase protein production in transgenic plantlets: The β-1,3-glucanase enzyme activity was assayed by measuring the rate of reduction of sugar production with laminarin as substrate. There were variation in the concentration of β-1,3-endoglucanase among the transgenic banana cultivar, Rastali (AAB) plantlets including the untransformed plantlets before and after Fusarium oxysporum spores inoculations. However, all transgenic plantlets had significantly higher enzyme activity compared to the untransformed plantlets (Table 4).
The β-1,3-glucanase activity of B2E (2) and B3E (31) was 3 fold higher than untransformed plantlet. The concentration of β-1, 3-endoglucanase in transgenic plantlets ranged from 12.3 to 36.9 nkat mg-1 protein after infections with Fusarium spores.
| Table 4: | β-1,3 glucanase enzyme activity in transgenic Pisang Rastali (AAB) plantlets transformed during pre and post inoculation with Fusarium oxysporum f.sp cubense race 1 (VCG 01217) |
 | |
| Data were analysed using one-way ANOVA and the differences contrasted using Duncan’s multiple range test. Different letter (s) indicate values are significantly different (p<0.05) | |
A correlation between constitutive β-1,3-endoglucanase levels and tolerance has been detected in all transgenic banana cultivar, Rastali (AAB) plantlets.
In addition, transgenic potato plants expressing soybean β-1,3-endoglucanase gene exhibit an increase in the activity of β-1,3 glucanase in transgenic plants. The β-1,3-glucanase (glucan endo-1,3-β-glucosidases, EC 3.2.1.39) able to catalyse endo type hydrolytic cleavage of the 1,3-β-D-glucosidic linkages in β-1, 3-glucans. There is strong evidence that expression of β-1,3-glucanase transgene alone regulated by the strong CaMV 35S RNA promoter can reduce the susceptibility of plants to infection by certain fungi. Transgenic tobacco plants expressing a soybean β-1,3-glucan-elicitor releasing β-1,3-glucanase or the tobacco class II β-1,3-glucanase show reduced symptoms when infected with Alternaria alternate or the oomycetes Phytophthora parasitica var. nicotiane and Peronospora tabacina (Yoshikawa et al., 1993; Lusso and Kuc, 1996).
In addition, similar pattern of β-1,3-endoglucanase levels and resistance has also been detected in leaves of resistant tomato lines (Lycopersicon esculentum) infected with Alternaria solani (Lawrence et al., 1996) and leaf rust resistance in wheat (Anguelova et al., 1999). Nakamura et al. (1999) reported that transgenic kiwifruit with soybean β-1,3-endoglucanase cDNA showed up to six fold higher of β-1,3-endoglucanase activity than control plants which resulted the disease lesion areas in tranformants leaf were smaller than on control plants. A similar phenomenon was reported by Ito et al. (1995) using transformed eggplants with soybean β-1,3-endoglucanase cDNA. The transformed eggplants with higher glucanase activity showed a statistically significant degree of disease resistance to P. capsici. Yoshikawa et al. (1993) reported that transgenic tobacco with soybean β-1, 3-endoglucanase cDNA induced the transcription of a plant defense gene, phenylalanine ammonia lyase, in response to fungal attack to a greater extent than untransformed leaves.
In conclusion, we produced transgenic banana cultivar, Rastali (AAB) expressing the soybean β-1,3-endoglucanase gene and showed their increased tolerance to Fusarium wilt race 1 disease by inoculation of the plantlets. Our next step will be focused on the evaluation of the transgenic plants tolerance against various fungal pathogens under greenhouse and filed conditions.
ACKNOWLEDGMENTS
The generous gifts of a pROKla-Eg (soybean β-1,3-glucanase) plasmid by Dr. Yoji Takeuchi from Hokkaido University, Japan are gratefully acknowledged. This research was supported by the Ministry of Science, Technology and Environmental funded this research through IRPA PR Research Grant (01-02-04-0000-PR0010/04)
REFERENCES
- Anfoka, G. and H. Buchenauer, 1997. Systemic acquired resistance in tomato against Phytophthora infestans by pre-inoculation with tobacco necrosis virus. Physiol. Mol. Plant Pathol., 50: 85-101.
CrossRefDirect Link - Anguelova, V.S., A. Van der Westhuizen and Z.A. Pretorius, 1999. Intercellular proteins and β-1, 3-glucanase activity associated with leaf rust resistance in wheat. Physiol. Plant., 106: 393-401.
Direct Link - Antunez-de-Mayolo, G., S.N. Maximova, S. Pishak and M.J. Guiltinan, 2003. Moxalactam as a counter-selection antibiotic for Agrobacterium mediated transformation and its positive effects on Theobroma cacao somatic embryogenesis. Plant Sci., 164: 607-615.
Direct Link - Bean, S.J., P.S. Gooding, P.M. Mullineaux and D.R. Davies, 1997. A simple system for pea transformation. Plant Cell Rep., 16: 513-519.
CrossRefDirect Link - Bi, Y.M., B.P.A. Cammue, P.H. Goodwin, R.S. Krishnan and P.K. Saxena, 1999. Resistance of Botrytis cinerea iscented geranium transformed with a gene encoding the antimicrobial protein Ace-AmP1. Plant Cell Rep., 18: 835-840.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cheng, M., J.E. Fry, S.Z. Pang, H.P. Zhou and C.M. Hironaka et al., 1997. Genetic transformation of wheat mediated by Agrobacterium tumefaciens. Plant Physiol., 115: 971-980.
Direct Link - Ganapathi, T.R., N.N.S. Higgs, P.J.B. Kurthi, C.J. Arntzen, G.D. May and J.M.V. Eck, 2001. Agrobacterium-mediated transformation of embryogenic cell suspension of the banana cultivar Rasthali AAB. Plant Cell Rep., 20: 157-162.
Direct Link - Hiei, Y., T. Komari and T. Kubo, 1997. Transformation of rice mediated by Agrobacterium tumefaciens. Plant Mol. Biol., 35: 205-218.
CrossRefDirect Link - Hobbs, S.L.A., T.D. Warkentin and C.M.O. DeLong, 1993. Transgene copy number can be positively or negatively associated with transgene expression. Plant Mol. Biol., 21: 17-26.
CrossRefDirect Link - Holland, L., J.E. Gemmell, J.A. Charity and C. Walter, 1997. Foreign gene transfer into Pinus radiata cotyledons by Agrobacterium tumefaciens. New Zealand J. For. Sci., 27: 289-304.
Direct Link - Janna, O.A., M. Marziah and G.K.A. Parveez, 2001. Potential selective agents for orchid transformation. Asia Pac. J. Mol. Biol. Biotechnol., 8: 85-94.
Direct Link - Khanna, H., B. Dong, J. Kleidon and J. Dale, 2004. Centrifugation Assisted Agrobacterium tumefaciens-mediated Transformation (CAAT) of embryogenic cell suspension of banana (Musa spp. Cavendish AAA and Lady finger AAB). Mol. Breed., 14: 239-252.
CrossRefDirect Link - Kohli, A., M. Leech, P. Vain, D.A. Laurie and P. Christou, 1998. Transgene organization in rice engineered through direct DNA transfer supports a two phase integration mechanism mediated by the establishment of integration hot spots. Proc. Nat. Acad. Sci. USA., 95: 7203-7208.
PubMedDirect Link - Kononov, M.E., B. Bassuner and S.B. Gelvin, 1997. Integration of T-DNA binary vector �backbone� sequences into the tobacco genome: Evidence for multiple complex patterns of integration. Plant J., 10: 165-174.
CrossRefDirect Link - Kuvshinov, V., K. Koivu, A. Kanerva and E. Pehu, 1999. Agrobacterium tumefaciens-mediated transformation of greenhouse-grown Brassica rapa ssp. oleifera. Plant Cell Rep., 18: 773-777.
CrossRefDirect Link - Lawrence, C.B., M.H.A.J. Joosten and S. Tuzun, 1996. Differential induction of pathogenesis-related proteins in tomato by Alternaria solani and the association of a basic chitinase isozyme with resistance. Physiol. Mol. Plant Pathol., 48: 361-377.
CrossRefDirect Link - Lawrence, P.K. and K.R. Koundal, 2001. Agrobacterium tumefaciens-mediated transformation of pigeon pea (Cajanus cajan (L.) Mill sp.) and molecular analysis of regenerated plants. Curr. Sci., 80: 1428-1432.
Direct Link - Lusso, M. and J. Kuc, 1996. The effect of sense and antisense epression of the PR-N gene for β-1,3-glucanase on disease resistance of tobacco to fungi and viruses. Physiol. Mol. Plant Pathol., 49: 267-283.
Direct Link - Maximova, S.N., A.M. Dandekar and M.J. Guiltinan, 1998. Investigation of Agrobacterium-mediated transformation of apple using green fluorescent protein: High transient expression and low stable transformation suggest that factors other than T-DNA transfer are rate-limiting. Plant. Mol. Biol., 37: 549-559.
PubMedDirect Link - May, G.D., R. Afza, H.S. Mason, A. Wiecko, F.J. Novak and C.J. Arntzen, 1995. Generation of transgenic banana (Musa acuminata) plants via Agrobacterium-mediated transformation. Bio/Technol., 13: 486-492.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nadolska-Orczyk, A. and W. Orczyk, 2000. Study of the factors influencing Agrobacterium-mediated transformation of pea (Pisum sativum L.). Mol. Breed., 6: 185-194.
CrossRefDirect Link - Nakamura, Y., H. Sawada, S. Kobayashi, I. Nakajima and M. Yoshikawa, 1999. Expression of soybean b-1, 3-endoglucanase cDNA and effect on disease tolerance in kiwifruit plants. Plant Cell Rep., 18: 527-532.
CrossRefDirect Link - Park, S.U. and P.J. Facchini, 2000. Agrobacterium-mediated genetic transformation of California poppy, Eschsholzia california Cham., via somatic embryogenesis. Plant Cell Rep., 19: 1006-1012.
Direct Link - Parveez, G.K.A., M.K.U. Chowdhury and N.M. Saleh, 1996. Determination of minimal inhibitory concentration of selection agents for oil palm Elaeis guineensis Jacq. transformation. Asia-Pac. J. Mol. Biol. Biotechnol., 4: 219-228.
Direct Link - Pasakinskiene, I. and V. Paplauskiene, 1999. Floral meristems as a source of enhanced yield and quality of DNA in grasses. Plant Cell Rep., 18: 490-492.
CrossRefDirect Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Smith, R.H. and E.E. Hood, 1995. Agrobacterium tumefaciens transformation of monocotyledons. Crop Sci., 35: 301-309.
Direct Link - Sreeramanan, S., M. Maziah, M. Sariah, M.P. Puad and R. Xavier, 2006. Bioassay method for testing Fusarium wilt disease tolerance in transgenic banana. Sci. Hortic., 108: 378-389.
CrossRefDirect Link - Tonon, C., A. Andreu, M.E. Aued, M. Van Damme, M. Huarte and G.R. Daleo, 1998. Defence reactions in two potato cultivars following infection with two races of Phytophthora infestans. Potato Res., 41: 319-325.
CrossRefDirect Link - Weide, R., M. Koornneef and P. Zabel, 1989. A simple, nondestructive spraying assay for the detection of an active kanamycin resistance gene in transgenic tomato plants. Theor. Applied Genet., 78: 169-172.
CrossRefDirect Link - Yang, J.S., T.A. Yu, Y.H. Cheng and S.D. Yeh, 1996. Transgenic papaya plants from Agrobacterium-mediated transformation of petioles of in vitro propagated multishoots. Plant Cell Rep., 15: 459-464.
CrossRefDirect Link - Yoshikawa, M., M. Tsuda and Y. Takeuchi, 1993. Resistance to fungal diseases intransgenic tobacco plants expressing the phytoalexin elicitor-releasing factor β-1,3-endoglucanse from soybean. Naturwissenchaften, 80: 417-420.
Direct Link - Yu, T.A., S.D. Yeh and J.S. Yang, 2003. Comparison of the effects of kanamycin and geneticin on regeneration of papaya from root tissue. Plant Cell, Tissue Org. Cult., 74: 169-178.
CrossRefDirect Link - Hoekema, A., P. Hirsch, P.J.J. Hooykas and R.A. Schilperoort, 1983. A binary vector strategy based on separation of vir and T-region of the Agrobacterium tumefaciens Ti plasmid. Nature, 303: 179-180.
CrossRefDirect Link