
Reza Pourrahim
Department of Plant Virology, Plant Pests and Diseases Research Institute, P.O. Box 19395-1454, Tehran, Iran
Shirin Farzadfar
Department of Plant Virology, Plant Pests and Diseases Research Institute, P.O. Box 19395-1454, Tehran, Iran
Ali Reza Golnaraghi
Department of Plant Protection, College of Agriculture and Natural Resources, Science and Research Campus, Islamic Azad University, P.O. Box 14515-775, Tehran, Iran
Ali Ahoonmanesh
Department of Plant Pathology, College of Agriculture, Esfahan University of Technology, Esfahan, Iran
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 3 | Page No.: 254-259







ABSTRACT
The incidence and distribution of Grapevine fanleaf virus (GFLV) were investigated in two viticultural regions of northeastern Iran (Bojnurd and Kashmar) during the 2005 growing season. A total of 3,454 leaf samples were randomly collected from 25 vineyards and virus incidence was subsequently assessed by enzyme-linked immunosorbent assay (ELISA). Serological results were confirmed by reverse transcription-polymerase chain reaction using the specific primers designed to amplify a DNA fragment of approximately 1,500 bp corresponding to the GFLV-coat protein gene. The nucleotide sequence of the amplified DNAs of five selected GFLV Iranian isolates showed the highest identities to the same regions of other published GFLV sequences from GenBank. Based on ELISA, twenty two out of 25 surveyed vineyards had virus infection rates ranging from 4.2 to 15.8%. GFLV incidence in Bojnurd and Kashmar was 10.9 and 6.7%, respectively, with an overall incidence of 8.8%. In the fields surveyed, a widespread occurrence of Xiphinema index was also detected.
PDF Abstract XML References
How to cite this article
Reza Pourrahim, Shirin Farzadfar, Ali Reza Golnaraghi and Ali Ahoonmanesh, 2007. Incidence and Distributions of Grapevine fanleaf virus in North-East of Iran. Plant Pathology Journal, 6: 254-259.
DOI: 10.3923/ppj.2007.254.259
URL: https://scialert.net/abstract/?doi=ppj.2007.254.259
DOI: 10.3923/ppj.2007.254.259
URL: https://scialert.net/abstract/?doi=ppj.2007.254.259
INTRODUCTION
Virus diseases are a worldwide problem of grapevine crops and a major limiting factor for their production (Pearson and Goheen, 1990). Over 55 viruses or strains, belonging to 20 different virus genera, are able to infect this crop (Martelli, 2003). The oldest known virus disease of grapevine is infectious degeneration or fanleaf. The affected plant shows widely opened petiolar sinuses and abnormally gathered primary veins, resulting in the appearance of an open fan of the leaf (Martelli et al., 2001). The economic loss induced by the disease varies depending on grapevine cultivars. Yield losses of up to 80%, progressive decline of the vines, low fruit quality, reduction in vineyard longevity, low proportion of graft take, reduced rooting ability of propagation material and decreased tolerance to adverse climatic conditions have been cited as symptoms for affected sensitive cultivars (Martelli and Savino, 1990).
The causal agent of fanleaf, Grapevine fanleaf virus (GFLV), is a member of the genus Nepovirus within the family Comoviridae (Fauquet et al., 2005). GFLV has polyhedral particles with a size of approximately 30 nm in diameter (Martelli et al., 2001). Its genetic information is divided over two single-stranded positive sense RNA molecules, RNA-1 and RNA-2, coding for two polyproteins (Serghini et al., 1990; Ritzenthaler et al., 1991). This virus is distributed in most vineyards worldwide and can infect almost all Vitis species (Raski et al., 1983; Martelli and Savino, 1990). The host range of GFLV in nature is mainly restricted to Vitis sp., but the virus can occasionally infect weeds (Martelli et al., 2001). Distorting and chromogenic strains of GFLV have been described based on leaf symptoms they induce on grapevines (Martelli and Savino, 1990). GFLV is a mechanical and pollen transmissible virus (Martelli et al., 2001). The virus can be also transmitted by seeds (Lazar et al., 1990), but this has no significant epidemiological importance (Martelli, 1993). GFLV transmission from grape to grape under natural conditions occurs solely through the longidorid nematode Xiphinema index (Martelli et al., 2001). Spread of this virus over long distances can occur by transfer of infected propagation material (Martelli and Savino, 1990).
Grapevine (Vitis vinifera L.) is an important horticultural crop in Iran with the cultivation area of 313,315 ha and 2,795,925 tons of production (Anonymous, 2004). It is produced mainly in three distinct regions of Iran, namely northwest (including Azarbayejan-e-gharbi, Azarbayejan-e-sharqi, Hamedan, Qazvin and Zanjan provinces), northeast (Khorasan) and south (Fars). Khorasan is the first viticultural province in Iran for production (17%) and second for covered (16%) (Anonymous, 2004). Fanleaf symptoms were described for the first time in Iran in 1970 (Vuittenez, 1970). The causal agent of the disease was then identified as GFLV by Ghorbani (1988). This virus and its ectoparasite vector (X. index) was subsequently detected in vineyards of Ourmia, in Azarbayejan-e-gharbi province (Parvizy, 1989; Pourrahim et al., 2000). Also, Bermuda grass (Cynodon dactylon L.) was reported from Fars province as a potential reservoir host for GFLV (Izadpanah et al., 2003). The incidence and distribution of this virus were previously determined in northwestern and southern regions of Iran (Zaki-Aghl and Izadpanah, 2003; Rakhshandehroo et al., 2005).
In despite of the economical importance of grapevine in northeast Iran, no data is available on the incidence of GFLV and its impact on production. In the present study, 25 vineyards in Khorasan province were surveyed for the presence of this virus and of its soil-borne vector, X. index. The presence of GFLV in the collected leaf samples was assessed by enzyme-linked immunosorbent assay (ELISA), reverse transcription-polymerase chain reaction (RT-PCR) and, limited to few affected plants, by sequencing of a 1,500 bp DNA fragment corresponding to the coat protein gene.
MATERIALS AND METHODS
Field sampling: Surveys were conducted in vineyards of northeast region of Iran. Overall, 25 fields in two locations in Khorasan province, Bojnurd and Kashmar, were surveyed and 3,454 grapevine leaf samples of regional cultivars Asgari, Asgari-e-peykani, Kolahdari and Kaj-angur were collected during the early spring of 2005 (Table 1). Approximately 50-70 samples per hectare were randomly collected according an M-shaped pattern. In each location, the fields were randomly selected using a predetermined distance criterion, ranging from 2 to 5 km between fields. Samples were transported to the laboratory and kept at 4°C until tested by ELISA. Moreover, 59 soil samples from the vineyards surveyed in Bojnurd and 65 from Kashmar were collected. In each field, four to six samples were collected from the upper 30-70 cm of soil around the vine roots. The nematodes were extracted by the sieving method, hand picked from the resulting suspension and identified using a dissecting microscope (kindly by Sh. Barooti, PPDRI and Islamic Azad University, Tehran, Iran).
Serological assays: The initial screening for GFLV was carried out by double-antibody sandwich ELISA (DAS-ELISA) according to the general protocol described by Clark and Adams (1977).
| Table 1: | Results of ELISA tests run on grapevine samples randomly taken from 25 fields in Khorasan province of Iran using Grapevine fanleaf virus-polyclonal antibodies |
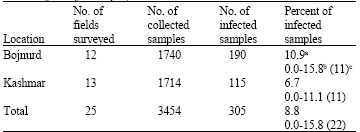 | |
| aAverage of percent virus incidence, bRange of percent virus incidence, cNumber of fields with virus infection | |
Serological reagents for the testing were purchased from Bioreba (Reinach, Switzerland). In ELISA tests, all buffers were prepared according to the manufacturer's instructions. Grapevine leaf samples were homogenized (1:5 Wt/Vol) in Tris extraction buffer using a sterile mortar and pestle and 100 μL volumes of the extracts were incubated overnight at 4°C in microtiter plate wells (Nunc Maxisorb, Denmark) previously treated with 100 μL of 1:1000 dilution of GFLV-IgG in carbonate coating buffer. Microtiter plate wells were then rinsed with washing buffer and incubated with 100 μL of the alkaline phosphatase-conjugated IgG diluted 1:1000 in conjugate buffer (3 h at 37°C). Wells were washed and incubated in 100 μL of substrate for 1 h at room temperature and the absorbance was determined at 405 nm using a microtiter plate reader (Multyscan-334, Finland). All samples were assayed in two repeats and results were judged to be positive if the mean absorbance was greater than or equal to three times the average reading of negative (healthy) controls. The presence of GFLV in a portion of samples was confirmed by RT-PCR.
RNA extraction: The procedure of Rowhani et al. (1993) was used for total RNA extraction. Briefly, small pieces of leaves of ELISA-positive samples were homogenized (1:10 Wt/Vol) in extraction buffer and centrifuged at 1,000 g for 3-4 min. The upper phase was then centrifuged at 16,800 g for 20 min. The pellet was resuspended in 2 mL of resuspension buffer; incubated for 10 min at 60°C in 250 μL of 10% SDS solution; centrifuged at 16,800 g for 15 min after adding 800 μL of 5 M potassium acetate and incubated at 4°C overnight. The RNA was finally precipitated by centrifugation at 16,800 g for 20 min in the presence of one-tenth of 3 M sodium acetate (pH 5.4) and an equal volume of isopropanol, then washed in 70% ethanol and resuspended in 50 μL of diethyl pyrocarbonate (DEPC)-treated water.
RT-PCR amplification: In addition to serological detection, 46 samples that reacted positive in ELISA were tested by RT-PCR technique. In this procedure, oligonucleotide primers (RT:5’-AAAAATTTGCATAACAGTAAAAAG-3’, C: 5’-CAAGGCAAGTGTGTCCAAA-3’ and V: 5’-TGATGCTTATAATCGGATAACTA-3’) designed to amplify a DNA fragment of approximately 1,500 bp corresponding to 3'UTR and GFLV-coat protein gene (Naraghi-Arani et al., 2001) were used. These primers were synthesized by MWG-Biotech Co. (Germany).
RT-PCR was performed in a two-step format using the extracted total RNA. Reverse transcription reaction was done in 20 μL volumes containing 4 μL of template RNA, 0.2 μL of the reverse primer RT (100 pmol μL-1) and 1 μL of RevertAidTM M-MuLV reverse transcriptase (Fermentas, Lithuania). This reaction was carried out using a top-heating thermal cycler (Mastercycler-5330, Eppendorf, Germany) at 42°C for 60 min and stopped by incubation at 70°C for 10 min, as suggested by the manufacturer. For PCR amplification in 50 μL reaction volumes, 0.2 μL of the primers C and V (100 pmol/μL), 5 μL of 10X Taq reaction buffer, 1.5 μL MgCl2 (50 mM), 1 μL dNTPs (10 mM) and 2.5 units Taq DNA polymerase (CinnaGen, Iran) were added to each 5 μL of first-strand cDNA reaction mixture. The PCR program consisted of a 2 min heating step at 95°C, followed by ten cycles of 30 sec at 95°C, 1 min at 63°C and 2 min at 72°C. Then, a 25-cycle amplification step of 30 sec at 95°C, 1 min at 63°C and 2 min at 72°C was performed.
PCR products and DNA ladder (GeneRulerTM 100 bp DNA Ladder Plus, Fermentas, Lithuania) were analyzed by electrophoresis through 1% agarose gels in the presence of 1 μg mL-1 ethidium bromide using 1X Tris-Borate EDTA buffer (Sambrook et al., 1989). Gels were visualized and photographed with UV-illuminator (Imago, The Netherlands).
Sequencing: To confirm the detection achieved by RT-PCR, the DNA amplicons of three GFLV-isolates from Bojnurd (KS-26, KS-31 and KS-35) and two from Kashmar (KS-24 and KS-25), were cleaned by Nucleospin (Macherey-Nagel, Germany) and cloned into pGEM-T Easy vector (Promega Corp, Madison, WI), according to the manufacturers’ instructions. Sequences from both strands of the cloned DNA in three independent clones were determined by the custom sequencing service of MWG Biotech. Co. (Germany). The five sequences obtained were compared with the other sequences available in the GenBank using Basic Local Alignment Search Tool (BLAST) (Altschul et al., 1997) at the National Center for Biotechnology Information. These sequences also were aligned and compared with the corresponding regions of other GFLV sequences available in the GenBank, including AF304013, AF304014, AF304015, AY017338, X16907 and X60775 by the Clustal W (Thompson et al., 1994) using MegAlign program ver. 5.00 from the DNASTAR package (DNASTAR, Madison, WI, USA) (Burland, 2000).
RESULTS
Incidence of GFLV: Based on ELISA results for randomly collected grapevine leaf samples, the incidence of GFLV was estimated as 8.8% in Khorasan province, ranging from 0 to 15.8% in the fields visited. This virus was detected in 305 out of 3,454 samples collected from 22 different vineyards. Vines in Bojnurd area showed a higher degree of GFLV-infection (10.9%) than that in Kashmar (6.7%). In other words, 190 out of 305 samples infected with GFLV were found in Bojnurd. Both symptom incidence and serological assay results revealed a variation in virus incidence among the different vineyards surveyed. Viral disease symptoms observed in association with GFLV-infection were mottling, mosaic, opening of petiolar sinus, leaf deformation, shortening of internodes, the presence of double nodes and zigzag growth of shoots (Fig. 1). In particular, opening of petiolar sinus was the most common symptoms observed in Khorasan province. GFLV-like symptoms were found in 82 out of 305 plants (26.9%) with positive reaction in ELISA to the virus. However, in many cases GFLV-infected vines did not show any clear-cut symptoms (223 samples, 73.1% of the total infection). In this survey, the presence of X. index (Fig. 2) was detected in 36 and 24 soil-samples collected from Bojnurd and Kashmar, respectively, representing almost all vineyards surveyed in the two areas (23/25 fields infested, 92.0%). The average of nematode numbers in the soil samples was 46.0, ranging from 0 to 221.
RT-PCR amplification and nucleotide sequence: The presence of GFLV in ELISA positive samples was confirmed by RT-PCR using specific primers. A PCR product with the expected size of approximately 1,500 bp (Fig. 3) was obtained for each of the RNA extracts investigated (Naraghi-Arani et al., 2001). No differences were observed between the sizes of PCR products of grapevine leaf extracts by using the same primers (data not shown). The PCR product of five GFLV isolates (KS-24, KS-25, KS-26, KS-31 and KS-35) was sequenced. The 1,433 nt long sequences obtained for these isolates were submitted to the GenBank, where the accession numbers DQ513332 to DQ513336 were respectively assigned. Nucleotide sequence analysis of the amplified fragments with BLAST revealed a very high identity to the available sequences of GFLV, confirming the result achieved by RT-PCR.
 | |
| Fig. 1: | Symptoms associated with natural infections by GFLV on grapevine. (A) widely open petiolar sinuses with abnormally gathered primary veins resulting in fan leaf symptoms; (B) double node; (C) poor fruit set; (D) mottling and mosaic and (E) denticulation of leaf margins |
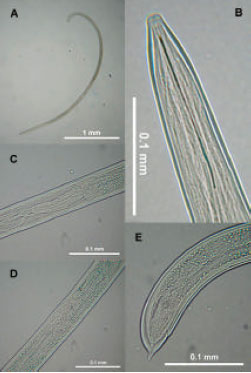 | |
| Fig. 2: | Xiphinema index. (A) female nematode (general view), (B) head and stylet, (C) terminal bulb, (D) reproductive system and (E) tail |
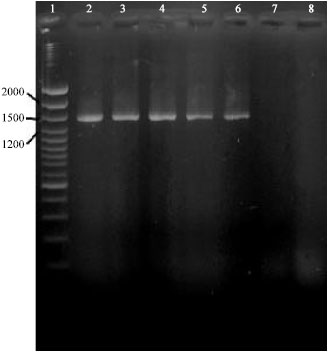 | |
| Fig. 3: | Detection of GFLV by RT-PCR. Ethidium bromide-stained agarose gel of PCR products. Lane 1, molecular weight marker (GeneRulerTM 100 bp DNA Ladder Plus, Farmentas, Lithuania); lane 2-6, a fragment of approximately 1,500 bp obtained using GFLV-specific primers; lane 7-8, healthy plant |
Computer analysis of these sequences using the Clustal W program showed high nucleotide identities ranged from 98.7 to 100%. The data indicated closest relation of our sequences to AF304014-USA (83.7-83.9%) and lowest to X16907-France (86.6-86.8%).
DISCUSSION
Prior to this study, no data was available on the occurrence, incidence and distribution of GFLV infecting grapevine plants in north-east of Iran. Based on ELISA results, the detection of GFLV in almost all vineyards surveyed was shown, such as the presence of its soil-borne vector, X. index. The incidence of GFLV at Bojnurd in Khorasan province was quite variable and higher than that of Kashmar, probably due to the more favorable conditions given by the higher humidity of soils in Bojnurd. Almost all the typical fanleaf symptoms, similar to those previously described (Martelli et al., 2001), i.e., mosaic, opened petiolar sinuses, short internodes and double nodes, were found in Iranian vineyards in association with GFLV-infections.
GFLV is widespread in different grapevine growing regions of Iran (Zaki-Aghl and Izadpanah, 2003; Rakhshandehroo et al., 2005; this study). This virus is transmitted from grape to grape by mechanical inoculations, nematode vectors, pollen or seeds (Lazar et al., 1990; Martelli et al., 2001); however, the infected propagative materials are considered the major and most efficient means of dissemination of this virus (Martelli, 1993). Therefore, it seems that the widespread occurrence of GFLV in the fields surveyed may have resulted from the absence of an efficient certification program for plant materials in Iran. In this respect, it seems justifiable to place increasing emphasis on optimizing quality control checks. The present study also reports cloning and sequencing of Iranian GFLV isolates for the first time. The information obtained in this study will hopefully aid in improving detection systems and control strategies for such infections in Iran. However, further investigations onto biological and molecular properties of GFLV isolates from vineyards of other parts of Iran should be carried out in this respect.
ACKNOWLEDGMENT
This research was supported by the Plant Virology Department of PPDRI (Tehran). We are grateful to Drs. M. Digiaro (Istituto Agronomico Mediterraneo, Valenzano, Italy) and H.R. Mojdehi for their kindly advice and to Sh. Barooti (PPDRI and Laboratory Complex, Science and Research Campus, Islamic Azad University, Tehran, Iran) for identification of nematodes. This research was carried out under project No. 107-0-11-81-064 of PPDRI.
REFERENCES
- Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Burland, T.G., 2000. DNASTAR's lasergene sequence analysis software. Methods Mol. Biol., 132: 71-91.
CrossRefPubMedDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Izadpanah, K., M. Zaki-Aghl, Y.P. Zhang, S.D. Daubert and A. Rowhani, 2003. Bermuda grass as a potential reservoir host for Grapevine fanleaf virus. Plant Dis., 87: 1179-1182.
Direct Link - Naraghi-Arani, P., S. Daubert and A. Rowhani, 2001. Quasispecies nature of the genome of Grapevine fanleaf virus. J. Gen. Virol., 82: 1791-1795.
Direct Link - Rakhshandehroo, F., R. Pourrahim, H. Zamani Zadeh, S. Rezaee and M. Mohammadi, 2005. Incidence and distribution of viruses infecting Iranian vineyards. J. Phytopathol., 153: 480-484.
Direct Link - Raski, D.J., A.C. Goheen, L.A. Lider and C.D. Meredith, 1983. Strategies against grapevine fanleaf virus and its nematode vector. Plant Dis., 67: 335-340.
CrossRefDirect Link - Ritzenthaler, C., M. Viry, M. Pinck, R. Margis, M. Fuchs and L. Pinck, 1991. Complete nucleotide sequence and genetic organization of Grapevine fanleaf nepovirus RNA1. J. Gen. Virol., 72: 2357-2365.
PubMedDirect Link - Rowhani, A., C. Chay, D.A. Golino and B.W. Falk, 1993. Development of a polymerase chain reaction technique for the detection of grapevine fanleaf virus in grapevine tissue. Phytopathology, 83: 749-753.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Serghini, M.A., M. Fuchs, M. Pinck, J. Reinbolt, B. Walter and L. Pinck, 1990. RNA2 of grapevine fanleaf virus: Sequence analysis and coat protein cistron location. J. Gen. Virol., 71: 1433-1441.
CrossRefDirect Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link