
Ali Reza Golnaraghi
Department of Plant Protection, College of Agriculture and Natural Resources, Science and Research Campus, Islamic Azad University, P.O. Box 14515-775, Tehran, Iran
Reza Pourrahim
Department of Plant Virology, Plant Pests and Diseases Research Institute, P.O. Box 19395-1454, Tehran, Iran
Shirin Farzadfar
Department of Plant Virology, Plant Pests and Diseases Research Institute, P.O. Box 19395-1454, Tehran, Iran
Ali Ahoonmanesh
Department of Plant Pathology, College of Agriculture,
Esfahan University of Technology, Esfahan, Iran
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 3 | Page No.: 227-234







ABSTRACT
A Tospovirus was isolated by mechanical sap transmission from potato plants showing leaf and stem necrosis symptoms and identified as Tomato yellow fruit ring virus (TYFRV) based on biological, serological and molecular studies. In mechanical transmission trials, a wide range of indicator plants, including members of the Amaranthaceae, Chenopodiaceae, Cucurbitaceae, Fabaceae and Solanaceae, closely similar to that of the not-infecting Nicotiana isolates of the same virus, was infected. The isolates under study strongly reacted with TYFRV antibodies, but not with the specific antibodies of the other tospoviruses tested (Groundnut ringspot virus, Impatiens necrotic spot virus, Iris yellow spot virus, Tomato chlorotic spot virus, Tomato spotted wilt virus and Watermelon silver mottle virus). In reverse transcription-polymerase chain reaction (RT-PCR), using specific primers for TYFRV-N gene, a DNA product with the expected size of approximately 1,200 bp was amplified from infected plants. The sequence analysis of this fragment showed a high identity with the corresponding region of TYFRV and confirmed the presence of this virus in Iranian potato crops. In this study, the transmission of TYFRV through potato tubers, even if at very low extent, was evidenced for the first time.
PDF Abstract XML References
How to cite this article
Ali Reza Golnaraghi, Reza Pourrahim, Shirin Farzadfar and Ali Ahoonmanesh, 2007. Identification and Partial Characterization of a Tospovirus Causing Leaf and Stem Necrosis on Potato. Plant Pathology Journal, 6: 227-234.
DOI: 10.3923/ppj.2007.227.234
URL: https://scialert.net/abstract/?doi=ppj.2007.227.234
DOI: 10.3923/ppj.2007.227.234
URL: https://scialert.net/abstract/?doi=ppj.2007.227.234
INTRODUCTION
Tospoviruses, within the family Bunyaviridae, are considered among the most common plant viruses of substantial agricultural importance, since they infect more than 1,000 plant species (Parella et al., 2003; Whitfield et al., 2005). Losses due to their infection in agricultural crops are significant worldwide (German et al., 1992), as in the case of Tomato spotted wilt virus (TSWV) that causes a total annual loss of 1 billion US$ in groundnut, vegetables, tobacco and other crops (Goldbach and Peters, 1994; Prins and Kormelink, 1998; Moyer et al., 1999).
Tospoviruses have quasi-spherical virions with lipid envelopes and a size of 80-120 nm in diameter, which contain three single-stranded RNA molecules (Cortez et al., 2001; Fauquet et al., 2005). There are at least 12 species of the genera Thrips and Frankliniella reported to transmit tospoviruses in nature (Mound, 2005). Tospovirus species are identified on the basis of their natural mode of transmission, host range, serological relationship of the N protein and their N protein sequence (Cortez et al., 2001; Fauquet et al., 2005). Tospoviruses, as well as their vectors, are worldwide distributed and have an extremely broad host range. These viruses can induce a range of symptoms, i.e., necrosis, chlorosis, ring patterns, mottling, silvering, stunting and local lesions, depending on virus species or strain, host plant, period of infection and environmental conditions (German et al., 1992; Mumford et al., 1996; Mound, 2005). TSWV is the most important Tospovirus species infecting potatoes (Solanum tuberosum) in the world (Nagata et al., 1995; Louro, 1996; Al-Shahwan et al., 1997; Khurana et al., 1997; Lathman and Jones, 1997; Jozsef et al., 2001; Wilson, 2001; Abad et al., 2005). Natural infections of other tospoviruses also have been previously reported in this crop: Groundnut ringspot virus (Granval de Milan et al., 1998), Impatiens necrotic spot virus (INSV) (Perry et al., 2005), Tomato chlorotic spot virus (TCSV) (Nagata et al., 1995; Granval de Milan et al., 1998) and a tospovirus serologically related to Peanut bud necrosis virus (PBNV) (Jefferies, 1998).
In Iran, several Tospovirus species of different serogroups (Moyer, 1999), including INSV, Iris yellow spot virus (IYSV), PBNV and TSWV, have been reported in several hosts (Moini and Izadpanah, 2001; Golnaraghi et al., 2002a; Ghotbi et al., 2005). In 2002, a distinct Tospovirus species infecting tomato plants in the Varamin area of Tehran province was identified and the name of Tomato yellow fruit ring virus (TYFRV) was for it proposed (Winter et al., 2002, 2006). A widespread occurrence of TYFRV in ornamental and soybean crops was reported in some provinces of Iran, where the virus was also detected in some weed species (Ghotbi et al., 2005; Golnaraghi et al., 2007). It seems that TYFRV isolates from soybean, originally mistook with TSWV, may be subdivided in two main groups: those readily infecting a broad range of Nicotiana species or cultivars and those not-infecting such indicators (Golnaraghi et al., 2002b). Based on ELISA tests, the thrips species Microcephallothrips abdominalis and Thrips tabaci were indicated as potential vectors for TYFRV (Ghotbi et al., 2005); the vectoring role of the latter was then experimentally demonstrated (Golnaraghi et al., 2007).
Potato is an important field crop which is widely cultivated throughout the world. Iran is the fifth largest potato producer in Asia with its 3.6 million tones/year (FAO, 2004). This crop is susceptible to infection by several viruses, which can substantially reduce yield and quality (Hooker, 1990). In recent years, Tospovirus-like symptoms of leaf and stem necrosis have been frequently observed in Iranian potato fields. In the present study, TYFRV was recognized as the causal agent of the disease based on host range studies and serological analysis. The identification of this virus was also confirmed by reverse-transcription polymerase chain reaction (RT-PCR) and sequencing. The information obtained in this study will hopefully aid in improving detection systems and control strategies for such infections in Iran.
MATERIALS AND METHODS
Plant sampling: In a survey done in the summer 2004, a total of 95 potato leaf samples from plants showing leaf and stem necrosis symptoms (Fig. 1a and b) were collected from four fields in Tehran province of Iran.
 | |
| Fig. 1: | Field symptoms associated with TYFRV infections on potato: (a) necrotic spots on leaves (b) leaf and stem necrosis and (c) a field with approximately 100% of infection |
| Table 1: | Tospovirus antibodies used in immunological assays |
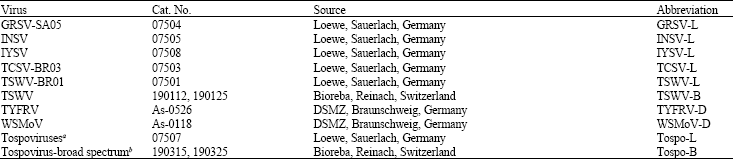 | |
| GRSV, Groundnut ringspot virus; INSV, Impatiens necrotic spot virus; IYSV, Iris yellow spot virus; TCSV, Tomato chlorotic spot virus; TSWV, Tomato spotted wilt virus; TYFRV, Tomato yellow fruit ring virus; WSMoV, Watermelon silver mottle virus. aan artificial mixture of TSWV-BR01, TCSV, GRSV and INSV, brecognizes serogroups I, II and III within the genus Tospovirus | |
Each plant sample was placed in a separate plastic bag, immersed in ice and stored at 4°C until tested by enzyme-linked immunosorbent assay (ELISA) for the presence of different tospovirus species (Table 1).
Enzyme-linked immunosorbent assay (ELISA): Double antibody sandwich ELISA (DAS-ELISA) (Clark and Adams, 1977) was adopted for testing different tospoviruses by using specific polyclonal antibodies. Specific antisera against TYFRV and Watermelon silver mottle virus (WSMoV) were kindly provided by S. Winter (DSMZ, Braunschweig, Germany), whereas those for GRSV, INSV, IYSV, TCSV and TSWV were purchased from Loewe (Sauerlach, Germany) or Bioreba (Reinach, Switzerland) (Table 1). One hour after adding the substrate, the absorbance value (A405 nm) was measured by a microplate reader (Tecan Spectra, Austria). Reactions were considered positive if the readings were equal to or exceeded three times the mean value for negative control.
Host reaction studies: Symptomatic leaf samples from two ELISA-positive potato plants to TYFRV were macerated in 0.05 M sodium phosphate buffer containing 0.1% 2-mercaptoethanol (pH 7.0) and mechanically inoculated onto Nicotiana rustica plants dusted with carborundum. Systemic infections of these plants with TYFRV were investigated by ELISA. The two virus isolates, named TY-PF12 and TY-PF28, were isolated by three serial single-lesion inoculation passages on cowpea (Vigna unguiculata) leaves. In the host range studies, systemically infected N. rustica leaves were used for inoculating Amarathaceae, Chenopodiaceae, Cucurbitaceae, Brassicaceae, Fabaceae and Solanaceae plant species (Table 2). The inoculated plants were kept under greenhouse conditions and the presence of virus was ascertained by observing eventual local and systemic symptoms induced and by ELISA tests. In host range trials, 2-10 plants of each species or cultivar were inoculated and the tests were repeated twice or more if the results were negative or uncertain. The virus isolates were maintained in freeze-dried infected N. rustica leaf tissue and stored at -70°C.
Serological analyses: For their serological characterization, TY-PF12 and TY-PF28 isolates were further analysed by ELISA, using the available antisera against different tospoviruses (Table 1).
Reverse transcription-polymerase chain reaction (RT-PCR): Total RNA was extracted from 100 mg healthy and infected leaf tissue using the RNA extraction solution (RNX plusTM, CinnaGen Inc., Tehran, Iran), as described by the supplier. RNA was finally precipitated by isopropanol, washed in 75% ethanol, resuspended in diethyl pyrocarbonate (DEPC)-treated water and stored at -70°C until use.
In this procedure, forward (TYf: 5’-TTTTTGTTTTTTGTTGTTTTTGGTTTTTTGTTT TTTGA-3’) and reverse (TYr: 5’-AGAGCAATCGAGGTATAACACATT-3’) primers previously described for amplification of a DNA fragment of approximately 1,200 bp corresponding to TYFRV-N gene (Golnaraghi et al., 2007) were used. These primers were synthesized by MWG-Biotech Co. (Germany). First single-strand cDNA (ss cDNA) was synthesized using 5 μL of template RNA (1.2 μg), 1 μL of the primer TYr (20 pmol μL-1) and 1 μL of RevertAidTMM-MuLV reverse transcriptase (200 u μL-1) (Fermentas, Lithuania) in 20 μL reaction volume at 42°C for 60 min and then at 70°C for 10 min to inactivate the enzyme, according to the manufacturer’s instructions. For the amplification reaction, ss cDNA was used as a template for the specific PCR using Pfu DNA polymerase (purchased from Cinnagen, Iran) and TYFRV specific primers. The PCR program consisted of 94°C for 1 min, 30 cycles of 94°C for 30 sec, 47°C for 1 min, 72°C for 1 min and finally 72°C for 10 min (Golnaraghi et al., 2007). Thermocycling was done in Primus (MWG Biotech Co., Germany). PCR products and DNA ladder (GeneRulerTM 100 bp DNA Ladder Plus, Farmentas) were fractionated by electrophoresis on a 1% agarose gel in the presence of 1 μg mL-1 ethidium bromide (Ausubel et al., 1993).
Cloning and sequencing: To confirm the RT-PCR result, the expected product of approximately 1,200 bp for TY-PF12 isolate was sequenced. The amplified DNA after PCR amplification was cleaned by Nucleospin (Macherey-Nagel, Germany) and cloned into pGEM-T Easy vector (Promega Crop., Madison, WI), according to the manufacturers’ instructions. Sequences from both strands of the cloned DNA in three independent clones were determined by the custom sequencing service of MWG Biotech. Co. (Germany). The nucleotide sequence obtained for N gene of this isolate was compared with other sequences available in the GenBank using Basic Local Alignment Search Tool (BLAST) (Altschul et al., 1997) at the National Center for Biotechnology Information. The nucleotide sequence and deduced amino acid sequence also were aligned and compared with the same region of other tospoviruses available in the GenBank by the Clustal W (Thompson et al., 1994) using Meg Align program ver. 5.00 from the DNASTAR package (DNASTAR, Madison, WI, USA) (Burland, 2000).
Transmission through potato tubers: To study virus transmission through potato tubers as potential inoculum sources, 50 potato plants of cv. Agria which were positive in ELISA to TYFRV were selected in a field in Tehran province, marked by wooden flags and their tubers harvested at the end of the growing season. The potato tubers were evaluated for the presence of symptoms; stored at 4°C in dark conditions for two months, planted individually in 10 cm plastic pots containing pasteurised soil and maintained in an insect-proof greenhouse. In total, 461 seedlings were separately tested by ELISA for the presence of TYFRV.
RESULTS
In the survey carried out in the summer 2004, almost all plants grown in some fields in Tehran province were damaged and showed leaf and stem necrosis symptoms (Fig. 1c). Significant populations of Thrips tabaci were also found in the affected fields. Based on ELISA results, 100% of symptomatic potato leaf samples were positive to TYFRV. None of the samples reacted to the other tospoviruses tested.
Host range: In the host range study, TY-PF12 and TY-PF28 isolates infected Chenopodium amaranticolor, Ch. quinoa, Cucumis sativus, Gomphrena globosa, N. tabacum cvs. Samsun, Phaseolus vulgaris, Solanum nigrum, Vicia faba, Petunia hybrida and Vigna unguiculata plants, on which they induced chlorotic and/or necrotic local lesions. In Datura metel, D. stramonium, Lycopersicon esculentum, Nicotiana benthamiana, N. glutinosa and N. rustica the appearance of systemic symptoms of necrosis followed that of necrotic or chlorotic local lesions. No local or systemic symptoms were observed on Brassica rapa, N. tabacum cv. White Burley, Raphanus sativus, Rapistrum rugosum and Sysimbrium loeselii. When healthy potato plants (Solanum tuberosum cv. Agria) were artificially inoculated with these isolates, necrotic lesions followed by chlorosis and necrosis symptoms developed (Table 2 and Fig. 2), resembling those observed in the field (Fig. 1).
| Table 2: | Symptomatology induced by the potato Tospovirus (TY-PF12) on indicator plantsa |
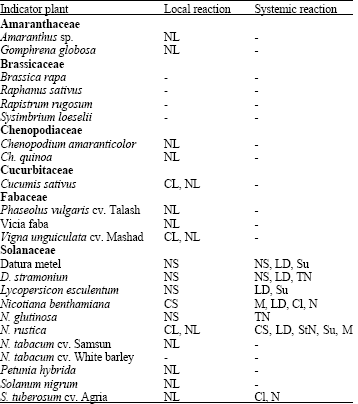 | |
| aThe host reaction obtained for TY-PF28 was similar to those for TY-PF12; Cl: Chlorosis; CL: Chlorotic Lesions; CS: Chlorotic Spots; LD: Leaf Deformation; M: Mosaic; N: Necrosis; NL: Necrotic Lesions; NS: Necrotic Spots; StN: Stem Necrosis; Su: Stunting; TN: Top Necrosis; -: No infection | |
 | |
| Fig. 2: | Symptoms induced by mechanical inoculation of TYFRV (TY-PF12) on Solanum tuberosum cv. Agria: (a) necrotic lesions and (b) systemic necrosis |
| Table 3: | Reaction (A405 nm) of the potato Tospovirus isolates (TY-PF12 and TY-PF28) with different tospovirus antibodies in double-antibody sandwich enzyme-linked immunosorbent assay formatsa |
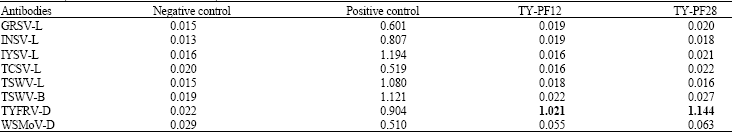 | |
| The highest absorbance value recorded for the two isolates is shown in bold. aThe average of A 405 nm values 60 min after adding substrate is indicated | |
| Table 4: | Tospoviral N protein sequence identities |
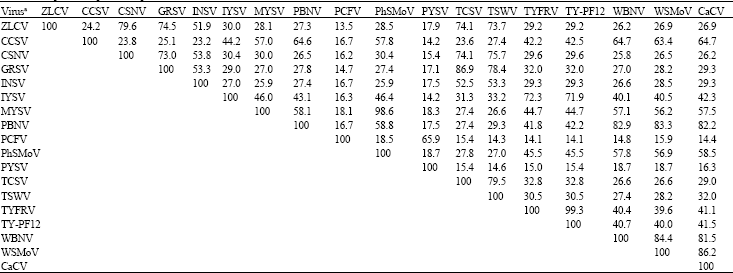 | |
| aThe tospovirus species referred to are: CCSV, Calla lily chlorotic spot virus (AY867502); CaCV, Capsicum chlorosis virus (DQ355974); CSNV, Chrysanthemum stem necrosis virus (AF067068); GRSV, Groundnut ringspot virus (AF513219); INSV, Impatiens necrotic spot virus (D00914); IYSV, Iris yellow spot virus (AF001387); MYSV, Melon yellow spot virus (AY673635); PBNV, Peanut bud necrosis virus (AY618564); PCFV, Peanut chlorotic fanspot virus (AF080526); PYSV, Peanut yellow spot virus (AF013994); PhSMoV, Physalis severe mottle virus (AF067151); TCSV, Tomato chlorotic spot virus (AF282982); TSWV, Tomato spotted wilt virus (AY856342); TYFRV, Tomato yellow fruit ring virus (AJ493270); WBNV, Watermelon bud necrosis virus (AF045067); WSMoV, Watermelon silver mottle virus (AY514627); ZLCV, Zucchini lethal chlorosis virus (AF067069) and TY-PF12 (DQ419916) in this study | |
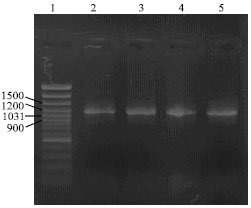 | |
| Fig. 3: | Detection of TYFRV by RT-PCR. Ethidium bromide-stained agarose gel of PCR products. Lane 1, molecular weight DNA marker (GeneRulerTM 100 bp DNA Ladder Plus, Farmentas, Lithuania); lane 2-5, a fragment of approximately 1,200 bp obtained using TYf and TYr primers |
The experimental host range and the kind of symptoms observed were similar for both the isolates under study.
Serological analyses: To study the serological correlation of TY-PF12 and TY-PF28 isolates with other tospovirus species, inoculated N. rustica plants were tested in ELISA against antibodies to different tospoviruses (Table 1). Both isolates strongly reacted in DAS-ELISA when the TYFRV-antiserum (DSMZ, As-0526) was used; at contrary, no significant reactions were observed in ELISA tests using specific antibodies to GRSV, INSV, IYSV, TCSV, TSWV and WSMoV (Table 3).
RT-PCR and sequencing: DNA fragments with the expected size of approximately 1,200 bp were amplified by RT-PCR using the primer pairs TYf and TYr from the total RNAs extracted from N. rustica plants infected with TYFRV (Fig. 3). No products were obtained with RNAs from healthy plants. The nucleotide sequence of the TY-PF12 amplicon was determined (GenBank accession No. DQ419916) and compared with those in the GenBank using the BLAST algorithm. A high identity to TYFRV genome was revealed, thus confirming the results achieved by RT-PCR and that, this sequence corresponded to a tospoviral N gene.
 | |
| Fig. 4: | Nucleotide sequence and deduced amino acid sequence of TY-PF12 N gene. The amino acid sequence was predicted from the complementary sense of the nucleotide sequence shown |
The sequence obtained (Fig. 4) was 1116 nucleotides in length and contained an ORF in the complementary strand starting with an AUG at nucleotide position 1069 and terminating with a UAA codon at position 245. The deduced N protein sequence was determined to be 274 residues long with a predicted molecular weight of about 30.1 KDa.
Computer analyses of the nucleotide sequence obtained for TY-PF12 revealed the highest identities to TYFRV (97.6%) and IYSV (75.9%). This sequence showed the lowest identities to Peanut yellow spot virus-PYSV (9.2%) and Peanut chlorotic fanspot virus-PCFV (3.9%).
Analyses of the deduced N protein showed the closest relationship of this isolate to the species TYFRV (99.3%) and IYSV (71.9%) and a distant relationship to PYSV (15.4%) and PCFV (14.1%) (Table 4).
Transmission through potato tubers: Symptoms of necrosis and malformation were observed in 7.2% of the tubers collected from TYFRV-infected potatoes. Also, in 11 cases (2.4%), tubers transmitted this virus to potato seedlings, that showed reduced growth and necrosis symptoms.
DISCUSSION
Very little information is available on the tospovirus infections of potato crops in the world. In the present study, the occurrence of a Tospovirus species in potato fields in Tehran province of Iran was demonstrated for the first time based on host range, serology and sequencing data. This virus caused severe leaf and stem necrosis on potatoes and sometimes the death of the whole plant. Electron microscope analysis showed the presence of enveloped spherical particles of ca. 100 nm in diameter in the infected material (data not shown), thus giving the first firm indication that a Tospovirus was involved (Fauquet et al., 2005). The wide experimental host range of two isolates studied (TY-PF12 and TY-PF28) was similar to that described previously for TYFRV (Ghotbi et al., 2005) and closely similar to that of not-infecting Nicotiana isolates from soybean (Golnaraghi et al., 2002b; 2007). In serological studies, a panel of tospovirus-specific antibodies did not react with these isolates, such as it previously occurred for TYFRV (Winter et al., 2006). Additional evidence of the identity of the studied viruses to TYFRV was obtained in RT-PCR, using specific primers for amplifying TYFRV-N gene, since amplicons with the expected size of approximately 1,200 bp (Golnaraghi et al., 2007) were detected in infected plants. The high identity to TYFRV was further confirmed by sequencing of the amplified fragment.
During this survey, the totality of potato plants showing leaf and stem necrosis symptoms were infected with TYFRV. Similar symptoms were frequently observed in the main Iranian potato growing areas, which were often ascribed by farmers to other causal agents, e.g., Alternaria, or to senescence phenomena. Therefore, deeper studies devoted to the determination of the incidence and the distribution of this virus in Iranian potato fields and on its potential importance on different potato cultivars seem justified. Very low levels of virus transmission through potato tubers were also evidenced for the first time in this work, the epidemiological value of which remains anyway of great practical importance and interest since the infected tubers represent the primary foci from which TYFRV inoculum could be further spread by vectors, e.g., Thrips tabaci (Golnaraghi et al., 2007). The rate of virus transmission through potato tubers may be influenced by many factors, including the period of infection and the cultivar, as reported for other potato viruses (Hooker, 1990; Loenbenstein et al., 2001; Wilson, 2001); so, higher levels of such transmission can be expected. Furthermore, cutting tools also will help in getting much higher percentage of transmission to healthy tubers and this, affecting TYFRV epidemiology and wide spreading of the virus.
This research represents a preliminary step towards the characterization of tospoviruses in potato crops in Iran. Other results of the study not reported in this paper revealed considerable differences among the host range of various TYFRV-potato isolates and the lack detection with the primers TYf and TYr. Therefore, further investigation of the biological and molecular properties of this virus and improving the effective tools for virus identification will be necessary for the disease management.
ACKNOWLEDGMENTS
This research was supported by the Plant Virology Department of PPDRI (Tehran) and Science and Research Campus of Islamic Azad University (Tehran). We are grateful to Dr. K. Ohshima (Laboratory of Plant Virology, Faculty of Agriculture, Saga University, Saga, Japan) for designing primers and his kindly comments in this research. The authors also wish to thank Drs. M. Digiaro (Istituto Agronomico Mediterraneo, Valenzano, Italy) and H.R. Mojdehi for their kindly advice and to Dr. S. Winter (DSMZ, Germany) for providing some of the antisera and positive controls used in this study. This research was part of Ph.D thesis of the first author and carried out under a project of Iranian Nanotechnology Initiative entitled use of microarray technique for detection of important viruses of potato.
REFERENCES
- Abad, J.A., J.W. Moyer, G.G. Kennedy, G.A. Holmes and M.A. Cubeta, 2005. Tomato spotted wilt virus on potato in eastern North Carolina. Am. J. Potato Res., 82: 255-261.
Direct Link - Al-Shahwan, I.M., O.A. Abdalla and M.A. Al-Saleh, 1997. Viruses in the northern potato-producing regions of Saudi Arabia. Plant Pathol., 46: 91-94.
CrossRefDirect Link - Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Burland, T.G., 2000. DNASTARs lasergene sequence analysis software. Meth. Mol. Biol., 132: 71-91.
PubMedDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Cortez, I., J. Saaijer, K.S. Wongjkaew, A.M. Pereira, R. Goldbach, D. Peters and R. Kormelink, 2001. Identification and characterization of a novel tospovirus species using a new RT-PCR approach. Arch. Virol., 146: 265-278.
CrossRefDirect Link - Fauquet, C.M., M.A. Mayo, J. Maniloff, U. Desselberger and L.A. Ball, 2005. Virus Taxonomy: Eighth Report of the International Committee on Taxonomy of Viruses. Academic Press, San Diego, CA., USA., ISBN-13: 9780080575483, Pages: 1162.
Direct Link - Ghotbi, T., N. Shahraeen and S. Winter, 2005. Occurrence of tospoviruses in ornamental and weed species in Markazi and Tehran provinces in Iran. Plant Dis., 89: 425-429.
Direct Link - Granval de Milan, N., R. Piccolo and O. Gracia, 1998. Potato (Solanum tuberosum L.) tuber transmission of tospoviruses (TSWV, GRSV and TCSV) in Mendoza, Argentina. Proceedings of the Recent Progress in Tospovirus and Thrips Research. Abstracts of Papers and Poster Presentations at the Fourth International Symposium on Tospoviruses and Thrips in Floral and Vegetable Crops, May 2-6, 1998, Wageningen, The Netherlands, pp: 46-47.
- Golnaraghi, A.R., R. Pourrahim, S. Farzadfar, K. Ohshima, N. Shahraeen and A. Ahoonmanesh, 2007. Incidence and distribution of tomato yellow fruit ring virus on soybean in Iran. Plant Pathol. J., 6: 14-21.
CrossRefDirect Link - Golnaraghi, A.R., R. Pourrahim, N. Shahraeen and S. Farzadfar, 2002. First report of Groundnut bud necrosis virus in Iran. Plant Dis., 86: 561-561.
CrossRefDirect Link - Khurana, S.M.P., S.K. Pandey, R.B. Singh and U.M. Bhale, 1997. Spread and control of the potato stem necrosis. Indian J. Virol., 13: 23-28.
Direct Link - Lathman, L.J. and R.A.C. Jones, 1997. Occurrence of tomato spotted wilt tospovirus in native flora, weeds and horticultural crops. Aust. J. Agric. Res., 48: 359-369.
Direct Link - Mound, L., 2005. Thysanoptera: Diversity and interactions. Ann. Rev. Entomol., 50: 247-269.
PubMedDirect Link - Mumford, R.A., I. Barker and K.R. Wood, 1996. The biology of the tospoviruses. Ann. Applied Biol., 128: 159-183.
Direct Link - Nagata, T., A.C. de Avila, C.J. Barbosa, P.C.T. Melo, F.C. Juliatti and E.W. Kitajima, 1995. Occurrence of different tospoviruses in six states of Brazil. Fitopatol. Brasil., 20: 90-95.
Direct Link - Perry, K.L., L. Miller and L. Williams, 2005. Impatiens necrotic spot virus in greenhouse-grown potatoes in New York State. Plant Dis., 89: 340-340.
Direct Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Whitfield, A.E., D.E. Ulman and T.L. German, 2005. Tospovirus-thrips interactions. Ann. Rev. Phytopathol., 43: 459-489.
Direct Link - Wilson, R.C., 2001. Resistance to infection and translocation of Tomato spotted wilt virus in potatoes. Plant Pathol., 50: 402-410.
Direct Link - Winter, S., N. Shahraeen, M. Koerbler and D.E. Lesemann, 2006. Characterization of Tomato fruit yellow ring virus: A new Tospovirus species infecting tomato in Iran. Plant Pathol., 55: 287-287.
Direct Link