
Shirin Farzadfar
Department of Plant Protection, College of Agriculture, Tehran University, Karaj, Iran
Ali Ahoonmanesh
Department of Plant Pathology, College of Agriculture, Esfahan University of Technology, Esfahan, Iran
Gholam Hossein Mosahebi
Department of Plant Protection, College of Agriculture, Tehran University, Karaj, Iran
Reza Pourrahim
Department of Plant Virology, Plant Pests and Diseases Research Institute, P.O. Box 19395-1454, Tehran, Iran
Ali Reza Golnaraghi
Department of Plant Protection, Islamic Azad University, P. 0. Box 14515-775, Tehran, Iran
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 1 | Page No.: 22-29







ABSTRACT
A survey was conducted to determine the occurrence and distribution of Cauliflower mosaic virus (CaMV) in several cruciferous crops in Iran. A total of 1451 symptomatic leaf samples were collected in 10 different Iranian provinces during the summers of 2004 and 2005 and then tested by enzyme-linked immunosorbent assay (ELISA) using specific CaMV-polyclonal antisera. Serological diagnosis was confirmed by herbaceous host range study, polymerase chain reaction (PCR) and sequencing. Spherical particles of approximately 50 nm in diameter were observed at electron microscope in symptomatic field samples. CaMV was detected in numerous cruciferous crops, with the highest incidence in cauliflower (Brassica oleracea var. botrytis) plantings. The virus occurred on B. oleracea var. acephala, B. oleracea var. capitata alba, B. oleracea var. italica, B. pekinensis, B. rapa, B. oleracea var. ganglyodes and Raphanus sativus in different provinces of Iran and in three brassica weeds (Rapistrum rugosum, Raphanus raphanistrum and Sisymbrium loeselii). Brevicoryne brassicae and Myzus persicae were the most widespread colonizing aphids of cruciferous crops in the surveyed areas, which also transmitted CaMV experimentally.
PDF Abstract XML References
How to cite this article
Shirin Farzadfar, Ali Ahoonmanesh, Gholam Hossein Mosahebi, Reza Pourrahim and Ali Reza Golnaraghi, 2007. Occurrence and Distribution of Cauliflower mosaic virus on Cruciferous Plants in Iran. Plant Pathology Journal, 6: 22-29.
DOI: 10.3923/ppj.2007.22.29
URL: https://scialert.net/abstract/?doi=ppj.2007.22.29
DOI: 10.3923/ppj.2007.22.29
URL: https://scialert.net/abstract/?doi=ppj.2007.22.29
INTRODUCTION
Brassicaceae are among the most economically important crops in the world. In recent years, their surface has significantly increased in Iran. According to FAO statistics, the cauliflower cultivated area increased from 800 ha in 2000 to 1000 ha in 2003; in the same period the production increased from 20,000 to 25,000 tons (FAO, 2000, 2003). Brassica crops can host a number of viruses, i.e., Beet western yellows virus (BWYV), Cauliflower mosaic virus (CaMV), Radish mosaic virus (RaMV) and Turnip mosaic virus (TuMV), that severely affect their production. Among them, CaMV is particularly feared for its widespread in temperate regions (Sutic et al., 1999).
CaMV, the type member of the genus Caulimovirus, has a circular double-stranded DNA genome of 8 Kbp which contains seven major ORFs and is replicated by the reverse transcription of an RNA intermediate (Haas et al., 2002; Mason et al., 1987). The virus has isometric particles of about 50 nm in diameter (Cheng et al., 1992). CaMV induces a range of systemic symptoms (chlorosis, mosaic, vein clearing and stunting), on many cruciferous plants, in particular on Brassica campestris and B. oleracea, where it is often found in mixture with TuMV (Shepherd, 1981). CaMV affects the plant development, especially in early infections when also the inflorescence is prevented. Low seed yields have been also reported (Sutic et al., 1999). CaMV incidence can easily exceed 70% and subsequent yields may be reduced up to 20-50% (Shepherd, 1981; Sutic et al., 1999). All CaMV isolates are able to infect a wide variety of cruciferous plants, but only a few of them can infect species of the family Solanaceae, including Datura and Nicotiana genera (Gracia and Shepherd, 1985; Lung and Pirone, 1972; Schoelz et al., 1986). CaMV is transmitted in nature in a non-circulative manner (Palacios et al., 2002) by at least 27 species of aphids (Kennedy et al., 1962). However, some CaMV isolates are apparently not aphid transmissible (Lung and Pirone, 1973). No transmission of CaMV by seeds or pollen occurs (Blanc et al., 2001).
In the last years, CaMV infections have been frequently reported from Iran on different cruciferous plants showing mild to severe symptoms (Farzadfar et al., 2005a). Despite the potential economic importance of such infections, the literature on CaMV in the country was limited to the signalizing of its natural occurrence on canola (B. napus) (Shahraeen et al., 2003). In this study, surveys were carried out during 2004 and 2005 growing seasons in order to identify the natural plant hosts of CaMV in 10 different provinces of Iran, thus representing the first large survey on the occurrence and distribution of this virus in the Iranian fields. The biological properties of different CaMV isolates from different species and provinces were also investigated. Moreover, aphid population analyses were also undertaken as part of this survey. The information presented in this study will hopefully aid in improving control strategies for CaMV infections in Iran.
MATERIALS AND METHODS
Plant sampling: A total of 1048 leaf samples from different brassica plants with typical virus disease symptoms (leaf deformation, mosaic, mottling, necrosis, rugosity, stunting, vein banding, vein clearing and yellowing) were collected from ten provinces of Iran during 2004 and 2005 growing seasons. Symptomatic plants included broccoli (Brassica oleracea var. italica), cauliflower (B. oleracea var. botrytis), Chinese cabbage (B. pekinensis), collard (B. oleracea var. acephala), kohlrabi (B. oleracea var. gongylodes), red cabbage (B. oleracea var. capitata rubra), small radish (Raphanus sativus var. sativus), turnip (B. rapa), white cabbage (B. oleracea var. capitata alba) and white radish (R. sativus) (Table 1). In addition, 403 cruciferous weed samples of charlock (Sinapis arvensis), hedge mustard (Sisymbrium officinale), mustard treacle (Erysimum sp.), persiankaali (B. deflexa), racket salad (Eruca sativa), rugose rapistrum (Rapistrum rugosum), small tumble-mustard (Sisymbrium loeselii) and wild radish (Raphanus raphanistrum) were collected from the surveyed fields. The leaf samples were immediately placed in plastic bags, labeled and kept at 4°C before testing.
| Table 1: | Occurrence and distribution of Cauliflower mosaic virus in different cruciferous crops in the main vegetable-growing provinces of Irana |
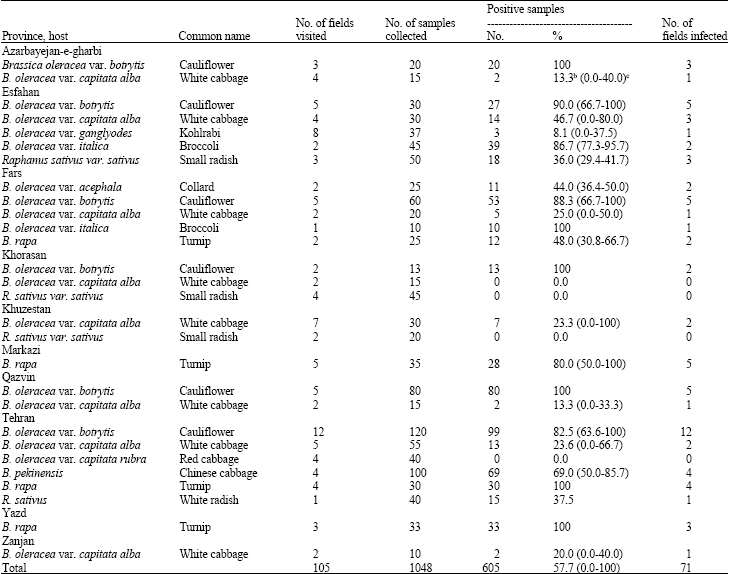 | |
| aIdentification based on serological reactions (ELISA); bAverage of virus infection rate in symptomatic samples; cRange of virus infection rate in symptomatic samples | |
Identification of aphid species in cruciferous fields: In order to identify the colonizing aphid species in the fields surveyed, 10-20 cruciferous plants per field were sampled. The aphids present in the samples were collected manually and preserved in 70% ethanol for counting and identification (kindly by A. Rezwani, Plant Pests and Diseases Research Institute, Iran).
Serological virus identification: Leaf samples were tested for the presence of CaMV by double-antibody sandwich enzyme-linked immunosorbent assay (DAS-ELISA) (Clark and Adams, 1977) using specific antibodies purchased from Loewe (Sauerlach, Germany). Polystyren microtitration plates (Maxisorb, NUNC, Denmark) were coated with 100 μL of IgG diluted 1:200 in carbonate coating buffer (15 mM Na2CO3, 35 mM NaHCO3 and 5 mM NaN3, pH 9.6) and incubated overnight at 4°C. Leaf samples (1:5 wt/vol) were ground in extraction buffer (3 mM KCl, 5 mM NaN3, 8 mM Na2HPO4, 1 mM NaH2PO4 and 0.13 M NaCl (PBS) containing 2% polyvinylpyrrolidone (PVP)-24,000 and 0.05% Tween 20, pH 7.4), then added to each well after washing of the plates for 4 times at 5 min intervals with washing buffer (0.05% Tween 20 in PBS). After incubation overnight at 4°C and washing of the wells, 100 μL of alkaline phosphatase-conjugated IgG diluted in conjugate buffer (2% PVP-24,000, 0.05% Tween 20, 0.2% Bovine Serum Albumin (BSA) and 1 mM MgCl2 in PBS, pH 7.4) were added. The plates were incubated for 3 h at 35°C and washed, then the wells were loaded with 100 μL of substrate buffer (1 mg mL-1 p-nitrophenyl phosphate and 3 mM NaN3 in diethanolamine buffer, pH 9.8) and left to incubate at room temperature. Absorbance values were determined at 405 nm by a microplate reader (Multiskan-334, Lab system, Finland) after 60 min. Samples with absorbance values greater than or equal to 3 times the average of negative samples were considered infected (positive).
Electron microscopy: Carbon and formvar-coated grids were floated on 20 μL of infected leaf extracts for 5 min (Milne and Lesemann, 1984). For trapping, coated grids were floated on one drop of CaMV antiserum (control grids were floated on normal rabbit serum), diluted 1:1000 with 0.05 M Tris buffer (pH 7.2), for 15-30 min at room temperature. After removing the unabsorbed serum by washing with 40 drops of Tris buffer, the grids were immediately floated on crude sap extracts for 30-45 min (Brlansky and Derrik, 1978). Plant debris were then removed by washing with 40 drops of distilled water. The grids were stained with 1% uranyl acetate and examined for the presence of virus particles with Phillips TEM-208 electron microscope.
Host range studies: Totally, 56 CaMV ELISA-positive samples from different hosts and locations were sap inoculated onto a wide herbaceous host range. The indicator plants used (Brassica juncea cv. Hakashina, B. napus cv. Otsubu, B. oleracea var. botrytis, B. rapa cv. Tatsuai and Raphanus sativus cv. Akimasari) were maintained in a greenhouse with at least 12 h light, 25±5°C and 50-70% relative humidity. Leaf samples were ground in 0.1 M Na-phosphate buffer (pH 7.0) containing 0.2% sodium sulfite (1:5 wt/vol) and rubbed onto carborundum-dusted leaves of the indicator plants at two-four leaf stage. In host range trials, at least two plants per species or cultivar were sap inoculated and the tests were repeated at least twice. Plants were kept in greenhouse for 4 to 6 weeks after inoculation, to observe the eventual appearance of virus symptoms and for virus detection by ELISA.
Aphid transmission: The aphid transmissibility of 45 CaMV isolates from different sources was tested using Myzus persicae as vector; the same test was carried out on 15 CaMV isolates using Brevicoryne brassicae. Aphids were starved for 1 h, given a 5 min acquisition access period on young leaves of B. rapa plants which had been previously infected (from 2 weeks) with CaMV isolates and placed on healthy young turnip test plants. Aphids were left to feed for 24 h and then killed by spraying with Confidor (Bayer, Germany). For each isolate at least 10 test plants were assayed, using 5 aphids per plant. The presence of CaMV in the inoculated plants was tested by ELISA.
Polymerase chain reaction (PCR): The presence of CaMV in 87 samples was also tested by PCR. Each leaf sample was ground in extraction buffer (250 mM NaCl, 0.25% sodium dodecyl sulphate, 30 mM EDTA and 100 mM Tris, pH 8) at a ratio of 1:4 (wt/vol). The extract was incubated with 10 μL of proteinase K (10 mg mL-1) per ml of tissue extract at 55°C for 60 min. The extract was diluted with an equal volume of distilled water and incubated at 95 to 100°C for 10 min. After cooling, the preparation was centrifuged at 13,000 g for 5 min and the supernatant was used as template for PCR analyses (Agama et al., 2002). Healthy turnip extracts were used as controls.
PCR amplifications were conducted in a 50 μL reaction volume containing 10 μL of DNA template, 1 μL each of the upstream and downstream primers (Agama et al., 2002) (20 pmol μL-1), 5 μL of 10X reaction buffer (200 mM Tris-HCl, 500 mM KCl, pH 8.4), 1.5 μL MgCl2 (50 mM), 1 μL dNTPs (10 mM) and 2.5 units Taq DNA polymerase (CinnaGen Inc., Tehran, Iran). These primers were synthesized by MWG Biotech. Co. (Germany). The PCR program consisted of a 5 cycle pre-amplification step of 30 ses at 92°C, 90 sec at 35°C and 40 sec at 72°C, followed by a single elongation step of 1 min at 95°C and 5 min at 72°C. Then, a 35 cycle amplification step of 35 sec at 94°C, 30 sec at 45°C and 40 sec at 72°C was performed (Agama et al., 2002). PCR reactions were conducted in a Primus (MWG Biotech. Co., Germany) thermal cycler.
PCR products and DNA ladder (GeneRulerTM 100 bp DNA Ladder Plus, Farmentas, Lithuania) were fractionated by electrophoresis on a 1% agarose gel in the presence of 1 μg mL-1 ethidium bromide using 1X Tris-Borate EDTA (TBE) buffer (89 mM Tris, 89 mM boric acid, 2 mM Na2EDTA, pH 8.3) (Sambrook et al., 1989). Gels were visualized and photographed with UV-illumination (Imago, The Netherlands).
Sequencing: To confirm the detection achieved by PCR, the expected PCR product (ca. 720 bp) for two isolates
were sequenced. The DNA fragments were cleaned from the amplification reaction mixture using Nucleospin kit (Macherey-Nagel, Germany) and cloned into pGEM-T Easy vector (Promega Crop., Madison, WI) according to the manufacturers’ instructions. Sequences from both strands of the cloned DNA were determined by the custom sequencing service of MWG Biotech. Co. (Germany). The nucleotide sequences obtained were compared with the other sequences available in the GenBank using Basic Local Alignment Search Tool (BLAST) (Altschul et al., 1997) at the National Center for Biotechnology Information.
RESULTS
Plant sampling: During the cultivation seasons of 2004 and 2005, surveys were conducted in 105 commercial fields distributed over 10 provinces of Iran (Fig. 1), where cruciferous plants were grown under different environmental conditions and/or cultural practices.
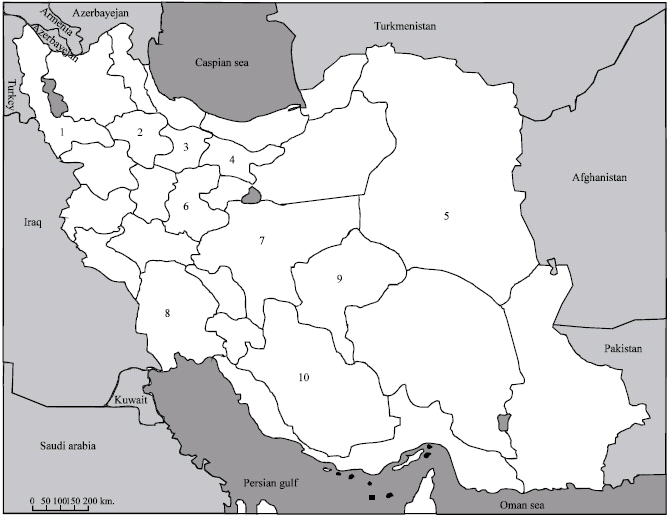 | |
| Fig. 1: | Map of Iran showing the location of provinces (1 to 10) where cruciferous crops were surveyed during the growing seasons 2004 and 2005; 1: Azarbayejan-e-gharbi, 2: Zanjan, 3: Qazvin, 4: Tehran, 5: Khorasan, 6: Markazi, 7: Esfahan, 8: Khuzestan, 9: Yazd and 10: Fars. Courtesy of Farzadfar et al. (2002) |
 | |
| Fig. 2: | Symptoms associated with natural infections by CaMV in Iran. A) banding mosaic and vein-clearing on cauliflower (Brassica oleracea var. botrytis); B) mosaic on small radish (Raphanus sativus var. sativus); C) leaf deformation, rugosity, stunting and vein clearing on Chinese cabbage (B. pekinensis); D) mosaic and vein banding on broccoli (Brassica oleracea var. italica); E) necrotic spots and vein yellowing on turnip (B. rapa); F) deformation, vein clearing, yellowing and stunting on rugose rapistrum (Rapistrum rugosum); G) deformation and vein yellowing on white cabbage (B. oleracea var. capitata alba) and H) deformation and stunting on small tumble-mustard (Sisymbrium loeselii) |
| Table 2: | Occurrence of Cauliflower mosaic virus in different cruciferous crops in Irana |
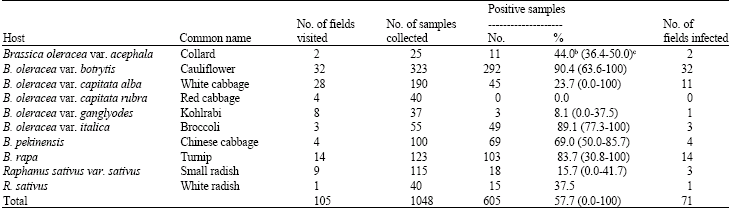 | |
| aIdentification based on serological reactions (ELISA), bAverage of virus infection rate in symptomatic samples; cRange of virus infection rate in symptomatic samples | |
Based on ELISA assays, CaMV was detected in 71 distinct fields, representing all 10 provinces surveyed. Incidence of CaMV in the symptomatic brassica plants varied in the different regions and among fields of the same regions. Of 1048 symptomatic samples collected, 605 (57.7%) reacted positively with antibodies to CaMV. Most of the cruciferous species surveyed, including broccoli, cauliflower, Chinese cabbage, collard, kohlrabi, small radish, turnip, white cabbage and white radish were infected. However, the highest incidence of CaMV was recorded in cauliflower samples (292/323 plants, corresponding to 90.4% of infection) (Table 1 and 2). CaMV infections were not confined to cruciferous crops. This virus was also detected on cruciferous weeds of Raphanus raphanistrum (7/31, 22.6%), Rapistrum rugosum (19/57, 33.3%) and Sisymbrium loeselii (11/41, 26.8%). Associated with CaMV infections were the symptoms of vein clearing, green vein banding, leaf deformation, necrosis, rugosity and stunting (Fig. 2).
Identification of aphid species in cruciferous fields: Brevicoryne brassicae and Myzus persicae (Aphididae) were identified as colonizing aphid species on brassica plants in the fields surveyed. B. brassicae was the prevailing aphid species in cruciferous fields, representing about 58.7% of the aphids trapped.
Host range and electron microscopy: In this study the electron microscopy and the mechanical transmission on different plant species were used to confirm the serological diagnosis. Spherical virus particles of approximately 50 nm in diameter were observed in ELISA-positive samples, confirming previous observations by Pirone et al. (1961). Sap inoculation of CaMV ELISA-positive samples induced symptoms of vein clearing, mosaic, rugosity and stunting preceded by chlorotic and necrotic local lesions on B. rapa cv. Tatsuai, mosaic on B. napus cv. Otsubu, vein clearing and mosaic on B. juncea cv. Hakashina, green vein banding on B. oleracea var. botrytis and Raphanus sativus cv. Akimasari. The host range and symptoms observed in this study agreed with those of previous reports (Brunt et al., 1995; Shepherd, 1981).
Aphid transmission: The CaMV isolates used in this study were transmitted by Myzus persicae and Brevicoryne brassicae. After about 16 days from aphid-inoculation, young B. rapa test plants showed systemic infections by CaMV, as confirmed by ELISA.
PCR amplification and nucleotide sequence: A DNA fragment of approximately 720 bp was obtained using CaMV-specific primers in PCR amplification (Agama et al., 2002) (Fig. 3).
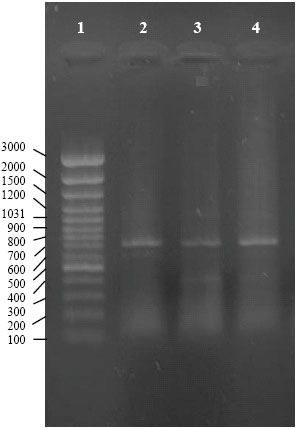 | |
| Fig. 3: | Detection of CaMV by PCR. Ethidium bromide-stained agarose gel of PCR products. Lane 1, molecular weight DNA marker (GeneRulerTM 100 bp DNA Ladder Plus, Farmentas, Lithuania); lane 2-4, a fragment of approximately 720 bp obtained using CaMV specific primers |
No amplicons were observed in similar preparations from healthy plants. The nucleotide sequence analysis of the amplified fragments of two selected virus isolates (GenBank accession nos. DQ119036 and DQ119037) with BLAST revealed a very high identity to the available sequences of CaMV, confirming the result achieved by PCR.
DISCUSSION
In the present study, the natural hosts and the distribution of CaMV were studied in different vegetable brassica fields of Iran. As expected, ELISA and PCR provided more accurate and consistent results than observation of field symptoms. Based on ELISA, this virus was present in most of fields surveyed. CaMV was detected in almost 58% of the 1048 symptomatic brassica plants tested using the specific antibodies. The highest incidence of CaMV infections was usually associated with cauliflower, broccoli and turnip plantings. In some symptomatic samples CaMV was not detected. It seems, therefore, that other virus species likely infect brassica plants in Iran, as previously reported (Farzadfar et al., 2005b; Farzadfar et al., 2005c; Shahraeen et al., 2003).
Results of this investigation indicate that CaMV is widespread in most of the vegetable-growing areas of Iran; however, the incidence of CaMV-infections considerably differed among the fields surveyed (Table 1). This variation could be due to many ecological factors such as to the presence or abundance of virus reservoirs. In Iran, vegetable cruciferous crops are normally sown in spring and autumn and mature in summer and winter, respectively, thus, the spring sown crops may act as inoculum source for the second crop in the same region. Also, in some locations, cruciferous plants were grown for seed production in a period of two years.
This study indicates that Brassica oleracea var. acephala, B. oleracea var. botrytis, B. oleracea var. capitata alba, B. oleracea var. italica, B. pekinensis, B. rapa and Raphanus sativus are natural hosts of CaMV in Iran. This virus was also detected on wild radish (Raphanus raphanistrum), as previously reported (Coutts and Jones, 2000). The natural occurrence of CaMV on Rapistrum rugosum and Sisymbrium loeselii has been reported for the first time in this study. The presence of CaMV in weeds was confirmed by mechanical transmission studies and PCR. The weed hosts analyzed in this study were the dominant weeds in the cruciferous fields surveyed. This survey was limited to 105 fields in 10 provinces of Iran; an extension of it to other areas of the country might reveal the presence of other new natural hosts for CaMV. In this study the presence of CaMV in volunteer canola plants was also reported (data not shown), supporting the previous report of natural occurrence of CaMV on canola plants (Brassica napus) in different Iranian provinces (Shahraeen et al., 2003). This crop is sown in autumn and harvested during spring and summer. In recent years, the total cultivated area of canola has increased from 200 ha in 1989 to 72,000 ha in 2003 (Anonymous, 2003). It seems likely that this change in cropping pattern may influence aphid population dynamics (Woodford, 1988) and subsequently, virus disease incidence in the same location, as previously described (Loenbenstein et al., 2001). Therefore, it appears that canola should be considered as an important virus bridging host for vegetable cruciferous plants in Iran.
Brevicoryne brassicae and Myzus persicae were frequently found colonizing brassica plants in the fields surveyed. Moreover, high populations of M. persica and B. brassicae were found in some fields where high incidence of CaMV (up to 100%) was recorded. The experimental evidence of CaMV transmission by the two species of aphids to young turnip plants had been previously reported (Brunt et al., 1995; Shepherd, 1981). Furthermore, the important role of non-colonizing winged aphids in spreading of CaMV should be further investigated. A more detailed surveys of insect populations and transmission studies would be useful in trying to identify the epidemiology of CaMV in Iran.
The results obtained in this investigation clearly demonstrate that CaMV is widely distributed in different geographical areas of Iran and provide evidence that this virus causes serious damages in cruciferous crops. The outbreaks of CaMV-infections can be minimized by several protective measures, as using healthy seedling transplants, eradicating old crops that are important infection foci, avoiding planting brassica crops sequentially in close proximity, rotation with non-host crops, choosing planting periods unfavorable to aphid population development, removing all weeds and volunteer plants in and around crops, removing symptomatic plants within the crop and having a brassica free growing period. However, considering the potential damage caused by CaMV, further studies towards determining more effective control strategies seem justified.
ACKNOWLEDGMENT
This research was supported by the Plant Virology Department of PPDRI (Tehran) and Plant Protection Department of Tehran University. We are grateful to Science and Research Campus of Islamic Azad University for its helps. We also wish to thank S. Sajedi for identification of weed species, to Dr. A. Rezvani for identification of aphids and to Drs. M. Digiaro (Istituto Agronomico Mediterraneo, Valenzano, Italy) and H.R. Mojdehi for their kindly advice. This research was part of PhD thesis of the first author and carried out under projects No. 2-009-100000-11-0000-84077 and 100-11-81050 of PPDRI.
REFERENCES
- Brlansky, R.H. and K.S. Derrik, 1979. Detection of seed borne plant viruses using serologically specific electron microscopy. Phytopathology, 69: 96-100.
CrossRefDirect Link - Brunt, A., K. Crabtree, M. Dallwitz, A. Gibbs and L. Watson, 1996. Viruses of Plants. CABI International, Wallingford, ISBN: 978-0-85198-794-1, Pages: 1488.
Direct Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Coutts, B.A. and R.A.C. Jones, 2000. Viruses infecting canola (Brassica napus) in South West australia Incidence distribution, spread and infection reservoir in wild radish (Raphanus raphanistrum). Aust. J. Agric. Res., 51: 925-936.
Direct Link - Farzadfar, Sh., R. Pourrahim, A.R. Golnaraghi and A. Ahoonmanesh, 2005. Occurrence of Cauliflower mosaic virus in different cruciferous plants in Iran. Plant Pathol., 54: 810-810.
CrossRefDirect Link - Farzadfar, Sh., K. Ohshima, R. Pourrahim, A.R. Golnaraghi, S. Jalali and A. Ahoonmanesh, 2005. Reservoir weed hosts for Turnip mosaic virus in Iran. Plant Dis., 89: 339-339.
CrossRefDirect Link - Gracia, O. and R.J. Shepherd, 1985. Cauliflower mosaic virus in the nucleus of Nicotiana. Virology, 146: 141-145.
Direct Link - Haas, M., B. Marin, A. Gelderich, P. Yot and M. Keller, 2002. Cauliflower mosaic virus: Still in the news. Mol. Plant Pathol., 3: 419-429.
CrossRefDirect Link - Lung, M.C.Y. and T.P. Pirone, 1972. Datura stramonium a local lesion host for certain isolates of cauliflower mosaic virus. Phytopathology, 62: 1473-1474.
CrossRefDirect Link - Mason, W.S., J.M. Taylor and R. Hull, 1987. Retroid virus genome replication. Adv. Virus Res., 32: 35-96.
PubMedDirect Link - Palacios, I., M. Drucker, S. Blanc, S. Leite and A. Moreno, 2002. Cauliflower mosaic virus is preferentially acquired from the phloem by its aphid vectors. J. Genet. Virol., 83: 3163-3171.
Direct Link - Schoelz, J.E., R.J. Shepherd and R.D. Richins, 1986. Properties of an unusual strain of cauliflower mosaic virus. Phytopathology, 76: 451-454.
Direct Link - Shahraeen, N., S.H. Farzadfar and D.E. Lesemann, 2003. Incidence of viruses infecting winter oilseed rape (Brassica napus ssp. oleifera) in Iran. J. Phytopathol., 151: 614-616.
Direct Link - Cheng, R.H., N.H. Olson and T.S. Baker, 1992. Cauliflower mosaic virus: A 420 subunit (T = 7), multilayer structure. Virology, 186: 655-668.
CrossRef - Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link