
Ali Reza Golnaraghi
Department of Plant Protection, College of Agriculture and Natural Resources, Science and Research Campus, Islamic Azad University, P.O. Box 14515-775, Tehran, Iran
Reza Pourrahim
Department of Plant Virology, Plant Pests and Diseases Research Institute,
P.O. Box 19395-1454, Tehran, Iran
Shirin Farzadfar
Department of Plant Virology, Plant Pests and Diseases Research Institute,
P.O. Box 19395-1454, Tehran, Iran
Kazusato Ohshima
Laboratory of Plant Virology, Faculty of Agriculture, Saga University, P.O. Box 840-8502, Saga, Japan
Nooh Shahraeen
Department of Plant Virology, Plant Pests and Diseases Research Institute,
P.O. Box 19395-1454, Tehran, Iran
Ali Ahoonmanesh
Department of Plant Pathology, College of Agriculture,
Esfahan University of Technology, Esfahan, Iran
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 1 | Page No.: 14-21







ABSTRACT
A total of 1,207 symptomatic and 2,677 randomly samples were collected from soybean fields throughout Iran during the growing seasons of 2004-2006 and tested by enzyme-linked immunosorbent assay (ELISA) using specific antibodies to Tomato yellow fruit ring virus (TYFRV), a newly proposed species in the genus Tospovirus. ELISA results were confirmed by electron microscopy, host range studies, serological analysis, polymerase chain reaction and limited to few affected plants, by sequencing of a 1200 bp DNA fragment corresponding to the N gene of TYFRV. The virus was largely detected in the main soybean growing provinces of Iran with an overall incidence of 6.1%, even if its introduction in Iranian soybean fields seems to be only recent. In this study, Thrips tabaci was identified for the first time as a vector of the virus, but no transmission by soybean seeds was evidenced for the same virus. TYFRV was also detected in some weed species and for the first time in Datura metel and D. stramonium, as confirmed by biological and molecular assays.
PDF Abstract XML References
How to cite this article
Ali Reza Golnaraghi, Reza Pourrahim, Shirin Farzadfar, Kazusato Ohshima, Nooh Shahraeen and Ali Ahoonmanesh, 2007. Incidence and Distribution of Tomato yellow fruit ring virus on Soybean in Iran. Plant Pathology Journal, 6: 14-21.
DOI: 10.3923/ppj.2007.14.21
URL: https://scialert.net/abstract/?doi=ppj.2007.14.21
DOI: 10.3923/ppj.2007.14.21
URL: https://scialert.net/abstract/?doi=ppj.2007.14.21
INTRODUCTION
Tospoviruses are considered as economically important pathogens in many crops in the world. Losses due to their infection in agricultural crops are significant worldwide (German et al., 1992), as in the case of Tomato spotted wilt virus (TSWV) that causes a total annual loss of US$ 1 billion in groundnut, vegetables, tobacco and other crops (Goldbach and Peters, 1994; Prins and Kormelink, 1998; Moyer et al., 1999).
Tospoviruses, within the family Bunyaviridae, have quasi-spherical virions with lipid envelopes and a size of 80-120 nm in diameter, which contains three single-stranded RNA molecules (Van Regenmortel et al., 2000; Cortez et al., 2001). There are at least 12 species of the genera Thrips and Frankliniella reported to transmit tospoviruses (Mound, 2005). These viruses can induce a range of symptoms, as necrosis, chlorosis, ring patterns, mottling, silvering, stunting and local lesions, depending on virus species or strain, host plant, time of infection and environmental conditions (German et al., 1992; Mumford et al., 1996). Tospoviruses have an extremely wide host range, comprising more than 1000 species in over 50 families including important ornamental, fruit and vegetable crops (Parella et al., 2003; Whitfield et al., 2005). Reports on natural tospovirus infections of soybean (Glycine max (L.) Merr.) are limited to Groundnut ringspot virus (GRSV) and Peanut bud necrosis virus (PBNV) from South Africa and India, respectively (Thakur et al., 1996; Pietersen and Morris, 2002). However, TSWV and Impatiens necrotic spot virus (INSV) are important emerging viruses in many parts of the world and could represent important limiting factors in soybean production areas (Hartman et al., 1999).
Since 1996, several species of the genus Tospovirus of different serogroups (Moyer, 1999), including INSV (Moini and Izadpanah, 2001; Ghotbi et al., 2005), Iris yellow spot virus (IYSV) (Ghotbi et al., 2005), PBNV (Golnaraghi et al., 2002) and TSWV (Bananej et al., 1996; Ghotbi et al., 2005) have been reported from Iran. In 2002, a virus disease inducing bright yellow ring patterns on tomato fruits was observed in tomato fields in the Varamin area of Tehran province. The causal agent was then identified as a distinct species in the genus Tospovirus and the name of Tomato yellow fruit ring virus (TYFRV) was for it proposed (Winter et al., 2002; Winter et al., 2006). A widespread occurrence of TYFRV in ornamental plants was reported in Markazi and Tehran provinces of Iran, where the virus was also detected in some weed species. Based on ELISA test, the thrips species Microcephallothrips abdominalis and Thrips tabaci were reported as potential vectors for TYFRV (Ghotbi et al., 2005).
With ca. 90,000 ha of cultivation area, soybean is one of the most important oilseed crops in Iran (FAO, 2005). This crop is susceptible to infection by several viruses, which can substantially reduce yield and quality (Sutic et al., 1999). Preliminary results indicated the prevalence of TYFRV, originally named as TSWW in soybean fields of Golestan province (Golnaraghi et al., 2000). In this study, the incidence and potential importance of the disease in the main soybean growing areas of Iran was determined. Natural weed hosts of the virus, its transmission by soybean seeds and thrips were also investigated. The information presented here will hopefully aid in improving control strategies for TYFRV infections in Iran.
MATERIALS AND METHODS
Soybean sampling: Surveys were conducted from July to October 2004 and 2005. Leaf samples from 1,055 soybean symptomatic plants (10-20 samples per field) were collected from 69 fields of Ardabil, Golestan, Khuzestan, Lorestan and Mazandaran provinces of Iran (Table 1). Symptoms consisted of leaf chlorosis, mosaic, mottling, yellowing, stunting and vein or top necrosis. In summer 2006, samples from other 152 symptomatic soybean plants were taken from three fields in Mazandaran. Additional 2.677 samples were also randomly collected from the 69 fields surveyed (Table 1), according to a zigzag-shaped pattern, with 3-5 m of distance between samples (30-50 samples per field). Each sample was placed in a separate plastic bag and stored at 4°C until testing by enzyme-linked immunosorbent assay (ELISA) for the presence of TYFRV. For the symptomatic samples, ELISA test was extended to other tospoviruses using a panel of specific antibodies (Table 2). Virus incidence in randomly collected samples was determined by ELISA and expressed as percentage of positive samples on total plants tested.
Weeds sampling: A total of 240 symptomatic weed samples were taken from inside or around three fields in Mazandaran (Table 3), one of the main soybean growing province in Iran. These samples were tested by ELISA for TYFRV detection.
| Table 1: | Incidence of TYFRV in Iranian provinces based on ELISA results- |
 | |
| a percent of fields with virus infection, b percent of samples with virus infection | |
| Table 2: | Tospovirus antibodies and ELISA procedures used in serological assays |
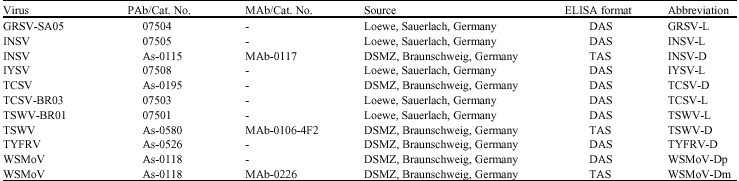 | |
| GRSV, Groundnut ringspot virus; INSV, Impatiens necrotic spot virus; IYSV, Iris yellow spot virus; TCSV, Tomato chlorotic spot virus; TSWV, Tomato spotted wilt virus; TYFRV, Tomato yellow fruit ring virus; WSMoV, Watermelon silver mottle virus; PAb, polyclonal antibody; MAb, Monoclonal antibody | |
| Table 3: | Extent of TYFRV infection in weed samples collected from three fields in Mazandaran province of Iran based on ELISA results |
 | |
Thrips collection: Thrips species were also investigated for their potential role as TYFRV vectors. In this study, 10-15 symptomatic soybean plants, as well as some weed species (Table 3), were sampled from three fields in Mazandaran. Thrips present on the plants were collected and identified at Plant Pests and Diseases Research Institute (Tehran, Iran).
Enzyme-linked immunosorbent assay (ELISA): Double antibody sandwich ELISA (DAS-ELISA) (Clark and Adams, 1977) and triple antibody sandwich ELISA (TAS-ELISA) (Adam et al., 1991) were adopted for testing different tospoviruses by using specific polyclonal and monoclonal antibodies. Specific antisera against INSV, Tomato chlorotic spot virus (TCSV), TSWV, TYFRV and Watermelon silver mottle virus (WSMoV) were kindly provided by S. Winter (DSMZ-Braunschweig, Germany), whereas those for GRSV, INSV, IYSV and TSWV-BR01 were purchased from Loewe (Germany) (Table 2). Alkaline phosphatase-conjugated rabbit anti-mouse (DSMZ) was used as the third antibody in TAS-ELISA method.
DAS-ELISA was also applied for testing the presence of TYFRV in adult thrips collected in soybean fields, according to the procedure described by Cho et al. (1988). Individual thrips was ground in 500 μL microtubes (Treff-Switzerland) using 50 μL of the extraction buffer. In this procedure, healthy and TYFRV-infected leaves of Nicotica rustica plants were used as negative and positive controls, respectively. One hour after adding the substrate, the absorbance value (A405 nm) was measured by a microplate reader (Tecan Spectra, Austria). Reactions were considered positive if the readings were equal to or exceeded three times the mean value for negative control.
Electron microscopy: In this procedure, carbon and formvar-coated grids were floated on 20 μL of crude leaf extracts for 5 min, washed with 40 drops of distilled water and negatively stained with 5 drops of 1% aqueous uranyl acetate (Milne and Lesemann, 1984). Grids were then examined with Phillips TEM-208 electron microscope.
Host reaction studies: Two ELISA-positive soybean samples showing chlorotic and mottled leaves were collected from Mazandaran and analyzed in a host range study. The plant species used were Chenopodium amaranticolor and Ch. quinoa (Chenopodiaceae), Citrulus vulgaris and Cucumis sativus (Cucurbitaceae), Gomphrena globosa (Amarathaceae), Glycine max cv. Hill, Phaseolus vulgaris, Pisum sativum, Vicia faba and Vigna unguiculata (Fabaceae), Datura metel, D. stramonium, Lycopersicon esculentum, Nicotiana benthamiana, N. glutinosa, N. rustica, N. tabacum cv. Samsun, N. tabacum cv. White Burley and Petunia hybrida (Solanaceae). Indicator plants were mechanically inoculated with plant extracts in 0.05 M sodium phosphate buffer (pH 7.0) containing 1% 2-mercaptoethanol. The inoculated plants were kept in a greenhouse with at least 12 h light, 25±5°C and 50-70% relative humidity, for observing eventual local and systemic symptoms induced by the transmitted viruses. ELISA test was performed three weeks after inoculation.
The two isolates, named TY-M23 and TY-M57, were also analyzed by ELISA, using the available antisera directed against different tospoviruses (Table 2).
Reverse transcription-polymerase chain reaction (RT-PCR) and sequencing: The presence of TYFRV in 21 ELISA-positive samples was also tested for a confirmation by RT-PCR. Specific primers TYf (5’-TTTTTGTTTTTTGTTGTTTTTGGTTTTTTGTTTTT TGA-3’) corresponding to nucleotides 1-38 of TYFRV (AJ493270) and TYr (5’-AGAGCAATCGAGGTATAACACATT-3’), complementary to the nucleotides 1156-1179, were used in RT-PCR. These primers, synthesized by MWG-Biotech Co. (Germany), were designed to amplify a DNA fragment of approximately 1200 bp of TYFRV-N gene. Total RNA was extracted from 100 mg of infected leaf tissue using 1 mL of the RNA extraction solution (RNX plusTM, CinnaGen Inc., Tehran, Iran) according to the manufacturer's instructions. Synthesis of first-strand cDNA was performed using 5 μL of template RNA (1.2 μg), 1 μL of the reverse primer (20 pmol μL-1) and 1 μL of RevertAidTMM-MuLV reverse transcriptase (200 u μL-1) (Fermentas, Lithuania) in 20 μL reaction volume at 42°C for 60 min. The mixture was then left at 70°C for 10 min to deactivate the enzyme, as recommended by the manufacturer. Reactions were carried out in 500 μL RNase-DNase free tubes (Treff-Switzerland). PCR amplification was performed in 50 μL reaction volumes containing 5 μL of the first-strand cDNA synthesis product, 1 μL of forward and 1 μL of reverse primer (20 pmol μL-1), 5 μL of 10X Taq reaction buffer (200 mM Tris-HCl, 500 mM KCl, 50 mM MgCl2, pH 8.4), 1.5 μL MgCl2 (50 mM), 1 μL dNTPs (10 mM) and 2.5 units Taq DNA polymerase (CinnaGen, Iran). Thermocycling was done in Mastercycler gradient (Eppendorf, Germany) according the following steps: 94°C for 1 min, 30 cycles of 94°C for 30 sec, 47°C for 1 min and 72°C for 1 min and finally 72°C for 10 min. PCR products were fractionated by electrophoresis through a 1% agarose gel in 1X Tris-borate EDTA buffer (Ausubel et al., 1993).
To confirm the detection achieved by RT-PCR, the expected product of approximately 1200 bp for TY-M23 isolate was sequenced. To this end, the DNA fragments were cleaned from the amplification reaction mixture using Nucleospin kit (Macherey-Nagel, Germany) and cloned into pGEM-T Easy vector (Promega Crop., Madison, WI) according to the manufacturer’s instructions. Sequences from both strands of the purified PCR product were obtained by the custom sequencing service of MWG Biotech. Co. (Germany). The nucleotide sequences obtained were compared with other sequences available in the GenBank using Basic Local Alignment Search Tool (BLAST) (Altschul et al., 1997) at the National Center for Biotechnology Information.
Thrips transmission: Thrips tabaci was tested for its ability to transmit the virus. Healthy insects used for this study were obtained from laboratory-reared colonies of thrips collected from onion plants in Tehran. Young tobacco seedlings (N. rustica) were freely exposed to a colony of thrips collected from infected soybean plants in Mazandaran. In a second test, larval thrips were allowed to feed for 4 days on tobacco plants previously inoculated with TYFRV and then transferred to healthy tobacco seedlings at a density of 10 insects per plant. After 15 day inoculation feedings, plants were sprayed with the insecticide Imidacloprid (Bayer, Germany) and kept in a greenhouse for symptom observation for 4 weeks. All plants were also tested by retro-inoculation on Vigna unguiculata and ELISA using TYFRV-specific antibodies.
Seed assays: To study virus transmission through seed, 50 soybean plants (G. max cv. Hill) which were positive in ELISA to TYFRV were selected in a field in Mazandaran, marked by wooden flags and their seeds harvested at the end of the growing season. Seeds were planted in groups of five in 10-cm plastic pots containing pasteurised soil and maintained in the greenhouse. In total, 1,540 seedlings were individually tested by ELISA for the presence of TYFRV at four-trifoliate leaf stage.
RESULTS
Detection of TYFRV in soybean samples by ELISA: Totally, 30.9% (326/1055) of the symptomatic soybean samples collected from 49 fields surveyed in 2004-2005 growing seasons and 93.4% (142/152) of symptomatic samples collected from 3 fields in 2006, were positive in ELISA to TYFRV-specific antibodies. None of the samples reacted to the other tospoviruses tested. During the field surveys, the main symptoms associated with TYFRV-infection included chlorosis, necrosis, mosaic and mottling, with the two latter symptoms prevailing in soybeans.
ELISA positive reactions were also obtained for 163 out of 2,677 samples (6.1%) randomly collected, distributed in 45 of 69 fields (65.2%), in all provinces surveyed. The highest virus incidence was recorded in Golestan (7.7%), followed by Mazandaran (7.6%), Lorestan (6.1%), Ardabil (3.6%) and Khuzestan (3.2%) provinces, with peaks of infection that in some locations or fields of Mazandaran province reached 28.6% (Table 1).
Detection of TYFRV in weed samples: Some of the weed species, including Amaranthus spp., Datura metel, D. stramonium, Euphorbia spp. and Lactuca spp. were positive to TYFRV in ELISA (Table 3). The presence of this virus in D. metel, D. stramonium and Lactuca spp. was also confirmed by host range studies and PCR amplification technique (data not shown).
Detection of TYFRV in individual thrips: Thrips of the species Frankliniella intonsa, Odontothrips confosus and Thrips tabaci were identified in the fields surveyed, with T. tabaci as predominant species. Furthermore, Exothrips sp., Haplothrips sp. and T. tabaci were also observed on the surrounding weeds.
| Table 4: | TYFRV detection in adult thrips collected from three soybean fields in Mazandaran by ELISA |
 | |
| a average of A405 nm values. In this study, the absorbance values obtained for positive and negative controls were 1.466 and 0.043, respectively | |
| Table 5: | Absorbance values (A405 nm) of the soybean isolates TY-M23 and TY-M57 in DAS- and TAS-ELISA using different polyclonal and monoclonal antibodies against different tospovirus species a |
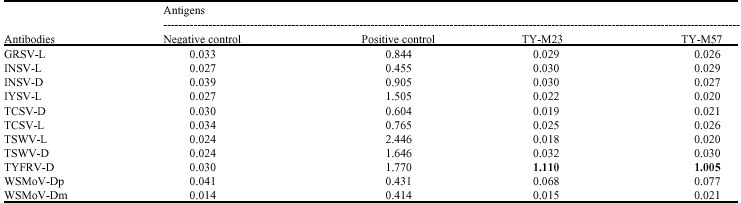 | |
| The highest absorbance value recorded for each of the two isolates (TY-M23 and TY-M57) is shown in bold; a Average of A405 nm values after sixty minutes by adding of substrate | |
 | |
| Fig. 1: | Symptoms induced by mechanical inoculation of TYFRV on Glycine max cv. Hill: (a) chlorotic and necrotic lesions on cotyledonous leaves; (b) mottling; (c) chlorosis |
DAS-ELISA carried out on adults of the different thrips species by using TYFRV-specific antibodies detected the presence of the virus in T. tabaci individuals (Table 4).
Host range and electron microscopy: Chlorotic or necrotic local lesions were induced on Chenopodium amaranticolor, Ch. quinoa, Cucumis sativus, Gomphrena globosa, Phaseolus vulgaris, Vicia faba and Vigna unguiculata plants inoculated with infected soybean extracts. Furthermore, Petunia hybrida showed the typical necrotic local lesions of tospoviruses. Systemic necrotic symptoms were observed in Datura metel, D. stramonium, Lycopersicon esculentum, Nicotiana benthamiana, N. glutinosa and N. rustica for the isolates under study. No symptom was observed on Citrulus vulgaris, N. tabacum cvs. Samsun and White barley and Pisum sativum.
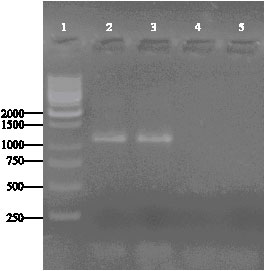 | |
| Fig. 2: | Detection of TYFRV by RT-PCR. Ethidium bromide-stained agarose gel of PCR products. Lane 1, molecular weight DNA marker (GeneRulerTM 1 kbp DNA Ladder, Farmentas, Lithuania); lane 2 and 3, a fragment of approximately 1200 bp obtained using TYf and TYr primers; lane 4 and 5, negative controls |
In this study, when healthy soybean seedlings (Glycine max cv. Hill) were inoculated with the above isolates, they showed chlorotic lesions followed by mottling, chlorosis and necrosis (Fig. 1), similar to those observed on the field-infected soybean plants. Virus particles of approximately 100 nm were observed in leaf extracts of TYFRV ELISA-positive samples. The results obtained from host range studies and electron microscopy observations for the soybean virus were similar to those previously reported for TYFRV (Ghotbi et al., 2005).
Serological analyses: In DAS-ELISA, TYFRV was clearly detected in crude extracts of plants infected with TY-M23 and TY-M57 isolates. The two isolates reacted with none of specific antibodies to GRSV, INSV, IYSV, TCSV, TSWV and WSMoV in DAS- or TAS-ELISA tests (Table 5), confirming previous reports (Winter et al., 2006).
RT-PCR and sequencing: An expected DNA fragment of approximately 1200 bp was amplified by RT-PCR from total nucleic acids extracted from the infected samples using primers TYr and TYf (Fig. 2). No amplicons were observed in similar preparations from healthy plants. The nucleotide sequence of the TY-M23 amplified fragment was determined (GenBank accession No. EF157855). BLAST search results revealed a high identity to TYFRV genome, thus confirming the results achieved by RT-PCR.
Thrips transmission: After three weeks of exposition to a colony of T. tabaci previously grown on TYFRV-infected soybeans, some tobacco plants (2/12, 16.7%) showed leaf chlorosis and diffuse necrosis symptoms. In another test, 7 out of 20 tobacco seedlings (35.0%) exhibited the same symptoms 21 days after the inoculation with larval thrips. TYFRV infection of these plants were confirmed by ELISA. The same virus was also sap-transmitted on V. unguiculata, inducing chlorotic and necrotic local lesions.
Seed assays: According to ELISA results, none of the seeds from infected soybean plants transmitted TYFRV to germinated seedlings.
DISCUSSION
In this study, the incidence and distribution of TYFRV were studied in different soybean fields of Iran. Based on the symptom observation in field and in greenhouse, early infections of soybean plants with this virus result in more severe chlorotic and necrotic symptoms, if not the death of the entire plant. The lack of TYFRV detection in some soybean plants with tospovirus-like symptoms can be explained with the possible confusion with similar symptoms induced by other soybean viruses (Hartman et al., 1999). The ELISA results also showed that the main soybean cultivars grown in Iran (G. max cvs. Clark, Hill and Williams) are susceptible to TYFRV (unpublished data). This virus was detected in randomly collected samples from 45 different fields, representing all provinces surveyed, with considerable differences of incidence among the fields (from 2.1 to 28.8%). This difference could be due to many factors, as the presence or abundance of virus reservoirs. TYFRV has a wide experimental and natural host range, including field crop plants, ornamentals and weed species (Ghotbi et al., 2005; this study). Based on ELISA, TYFRV was abundant in Golestan and Mazandaran provinces (Table 1), which are characterized by moderate climatic conditions and where many potential hosts are cultivated (Anonymous, 2004). ELISA results also indicated high levels of TYFRV-infections in smaller fields, in particular at Qa’emshahr area of Mazandaran province. On the other hand soybean in Iran is usually sown in early summer and harvested in autumn. In some regions like Mazandaran, soybean is cultivated in mid spring and early summer and matures in early and late autumn, respectively. Therefore, the spring sown crop may act as inoculum source for the second crop in the same region.
TYFRV was serologically detected in some weed species of Amaranthus sp., Euphorbia sp. and Lactuca sp., confirming earlier reports (Ghotbi et al., 2005). The natural occurrence of TYFRV on Datura metel, D. stramonium and Lactuca sp., based on host range studies and PCR, is for the first time reported in this study. Weeds were commonly found in many soybean fields throughout Iran (personal observations), but weed samples collection was restricted to three fields in Mazandaran province of Iran. An extension of this survey to other areas of this country could reveal the presence of additional natural hosts for TYFRV.
In this study, high populations of Thrips tabaci were found in some of the affected soybean fields. Some adults collected in these fields gave positive reaction in ELISA against TYFRV antibodies, confirming previous reports (Ghotbi et al., 2005). This virus was also transmitted by T. tabaci under experimental conditions from infected soybean to young tobacco plants, giving the first evidence of the natural transmission of TYFRV by thrips vectors. However, the possible role of other thrips species in transmission of the same virus should be further investigated.
During the last decade from the first description of a Tospovirus disease in Iran (Bananej et al., 1996), such infections have been frequently reported on tomato (Winter et al., 2002; Winter et al., 2006), potato (Pourrahim et al., 2001), peanut (Golnaraghi et al., 2001; Golnaraghi et al., 2002), tobacco (Moini and Izadpanah, 2001) and ornamental crops (Moeini and Izadpanah, 2000; Ghotbi et al., 2005), as well as on weed species (Ghotbi et al., 2005) in several provinces. This study indicates that TYFRV, a new emerging Tospovirus on soybeans in the world, is widespread across most of the soybean-growing areas of Iran. In the survey done in 2006, over 90% of symptomatic plants tested positive to TYFRV. The data strongly indicate the possible rise in the incidence of tospovirus infections in Iran and its potential economic importance, especially in soybean crops. Therefore, more studies towards determining effective control strategies for this disease seem justified. Also, further investigation on the biological and molecular properties of Iranian TYFRV isolates seems necessary in order to investigate more accurately the differences in the biological behaviour observed in some TYFRV isolates and the lack detection of amplicons for some of the isolates under study with the primers TYr and TYf (data not shown).
ACKNOWLEDGMENTS
This research was supported by Science and Research Campus of Islamic Azad University (Tehran) and Plant Virology Department of Plant Pests and Diseases Research Institute (Tehran). The authors wish to thank Dr. S. Winter for providing some of the antisera and positive controls used in this study and to Drs. M. Digiaro (Istituto Agronomico Mediterraneo, Valenzano, Italy) and H.R. Mojdehi for their kindly advice. This research was part of PhD thesis of the first author.
REFERENCES
- Adam, G., D.E. Lesemann and H.J. Vetten, 1991. Monoclonal antibodies against Tomato spotted wilt virus characterization and application. Ann. Applied Biol., 118: 87-104.
Direct Link - Cho, J.J., R.F.L. Mau, R.T. Hamasaki and D. Gonsalves, 1988. Detection of tomato spotted wilt virus in individual thrips by enzyme linked immunosorbent assay. Phytopathology, 78: 1348-1352.
Direct Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Cortez, I., J. Saaijer, K.S. Wongjkaew, A.M. Pereira, R. Goldbach, D. Peters and R. Kormelink, 2001. Identification and characterization of a novel tospovirus species using a new RT-PCR approach. Arch. Virol., 146: 265-278.
CrossRefDirect Link - German, T.L., D.E. Ulman and J.W. Moyer, 1992. Tospoviruses: Diagnosis, molecular biology, phylogeny and vector relationships. Ann. Rev. Phytopathol., 30: 315-348.
CrossRefPubMedDirect Link - Ghotbi, T., N. Shahraeen and S. Winter, 2005. Occurrence of tospoviruses in ornamental and weed species in Markazi and Tehran provinces in Iran. Plant Dis., 89: 425-429.
Direct Link - Golnaraghi, A.R., N. Shahraeen, R. Pourrahim, S. Ghorbani and S. Farzadfar, 2001. First report of a Tospovirus infection of peanuts in Iran. Plant Dis., 85: 1286-1286.
CrossRefDirect Link - Golnaraghi, A.R., R. Pourrahim, N. Shahraeen and S. Farzadfar, 2002. First report of Groundnut bud necrosis virus in Iran. Plant Dis., 86: 561-561.
CrossRefDirect Link - Hartman, G.L., J.B. Sinclair and J.C. Rupe, 1999. Compendium of Soybean Diseases. 4th Edn., The American Phytopathological Society, Minnesota, USA.
Direct Link - Mumford, R.A., I. Barker and K.R. Wood, 1996. The biology of the tospoviruses. Ann. Applied Biol., 128: 159-183.
Direct Link - Pietersen, G. and J. Morris, 2002. Natural occurrence of Groundnut ringspot virus on soybean in South Africa. Plant Dis., 86: 1271-1271.
Direct Link - Thakur, M.P., D.V.R. Reddy, A.S. Reddy, A.S. Ratna, M.A. Nasiri and K.C. Agrawal, 1996. Identification of bud blight of soybean (Glycine max (L.) Merr.) through ELISA and infectivity assay. Ind. J. Virol., 12: 79-82.
Direct Link - Van Regenmortel, M.H.V., C.M. Fauquet, D.H.L. Bishop, E.B. Carstens and M.K. Estes et al., 2000. Virus Taxonomy: Classification and Nomenclature of Viruses: Seventh Report of the International Committee on Taxonomy of Viruses. 1st Edn., Academic Press, New York, ISBN-10: 0123702003.
Direct Link - Winter, S., N. Shahraeen, M. Koerbler and D.E. Lesemann, 2006. Characterization of Tomato fruit yellow ring virus: A new Tospovirus species infecting tomato in Iran. Plant Pathol., 55: 287-287.
Direct Link - Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link