
M. Daami-Remadi
National Institute of Agronomic Research of Tunisia,
PRRDA-CE 4042 Chott-Mariem, 4042, Sousse, Tunisia
H. Jabnoun-Khiareddine
High School of Horticulture and Breeding of Chott-Mariem,
4042, Sousse, Tunisia
F. Ayed
High School of Horticulture and Breeding of Chott-Mariem,
4042, Sousse, Tunisia
M. El Mahjoub
High School of Horticulture and Breeding of Chott-Mariem,
4042, Sousse, Tunisia
Plant Pathology Journal
Year: 2006 | Volume: 5 | Issue: 3 | Page No.: 362-367







ABSTRACT
Several fungicides were tested against some isolates belonging to four Fusarium species causing potato tuber dry rot in Tunisia. Incorporated into the culture media PDA, the tested fungicides significantly inhibited the mycelial growth, observed after incubation at 25°C for 4 days, of all Fusarium isolates including those of F. sambucinum resistant to benzimidazoles. A significant interaction (p ≤ 0.05) was observed between both fixed factors where inhibition percentage varied depending on tested pathogens and fungicides. Applied on potato tubers (tuber immersion for 10 min) prior inoculation, certain tested fungicides such as azoxystrobin and fludioxonil significantly reduced by more than 50%, comparatively to the untreated controls, the development of dry rot occasioned by F. graminearum and F. sambucinum observed after 21 days of incubation at 25-27°C. A significant interaction (p ≤ 0.05) was noted between the treatments and the Fusarium species traduced by a variable inhibition percentage depending on tested pathogen and fungicides.
PDF Abstract XML References Citation
How to cite this article
M. Daami-Remadi, H. Jabnoun-Khiareddine, F. Ayed and M. El Mahjoub, 2006. In vitro and in vivo Interaction of Four Fungicides with the Fusarium Species Complex Causing Tuber Dry Rot in Tunisia. Plant Pathology Journal, 5: 362-367.
DOI: 10.3923/ppj.2006.362.367
URL: https://scialert.net/abstract/?doi=ppj.2006.362.367
DOI: 10.3923/ppj.2006.362.367
URL: https://scialert.net/abstract/?doi=ppj.2006.362.367
INTRODUCTION
In Tunisia, a complex of Fusarium species is responsible of potato tuber dry rot. F. solani, F. oxysporum f.sp. tuberosi and at a lesser frequency F. sambucinum and F. graminearum are present as mixed infections on tubers showing dry rot symptoms (Daami-Remadi and El Mahjoub, 1996, 2004; 2006; Priou and El Mahjoub, 1999; Chérif et al., 2001; Daami-Remadi et al., 2006a).
Benzimidazoles and conazoles fungicides were used since 1970 for the control of dry rot and other potato diseases (Leach, 1971; Murdock and Wood, 1972; Tisdale and Lord, 1973; Leach and Nielsen, 1975; Tivoli et al., 1986; Carnegie et al., 1990; Bang, 1992; Kawchuk et al., 1994; Mérida and Loria, 1994; Carnegie et al., 1998; Errampalli and Johnston, 2001). Thiophanate-methyl, carbendazim, iprodione, metalaxyl, procymidone and prochloraz inhibited dry rot development on tubers inoculated by F. sambucinum and other Fusarium species (Choiseul, 1996; Triki et al., 1996; Daami-Remadi and El Mahjoub, 1997; Chérif et al., 2001). However, a recent in vitro screening of some tunisian Fusarium spp. isolates for their resistance to some benzimidazoles showed that F. solani, F. oxysoprum f.sp. tuberosi and F. graminearum isolates are susceptible to these fungicides whereas F. sambucinum isolates are resistant (Daami-Remadi and El Mahjoub, 2006). These chemicals having a single-site mode of action are more likely to lead to development of resistance (Kawchuck et al., 2002). It is to note that since emergence of thiabendazole resistance, chemical combinations of 8-hydroxyquinolin and thiabendazole increased and they are largely used in France as an anti-thiabendazole resistance strategy (Tivoli et al., 1986). Recently, we have shown that in vitro and in vivo inhibition of benzimidazole resistant isolates was reached by several mixtures of fungicides tested individually or in dual combination (Daami-Remadi et al., 2006b). Furthermore, Beresford (1994) reported that fungicides with mode of action different to that of benzimidazoles could optimise dry rot control and minimize the incidence of the mentioned fungicide resistance. Leadbeater and Kirk (1992) found that fenpiclonil, a phenylpyrrole fungicide is more efficient as a pre-planting treatment than thiabendaole or imazalil. Furthermore, fludioxonil, added of mancozeb or difenoconazol, applied at pre-plantation of potato seeds inoculated by F. sambucinum, are shown to be efficient in inhibiting pathogen dissemination around progeny tubers (Bains et al., 2001).
Registered fungicides for potato dry rot control are lacking in the tunisian phytosanitary index (Anonymous, 2003). As some fungicides tested in previous studies against some isolates of F. oxysporum f.sp. tuberosi have shown efficacy in reducing potato vascular wilt (Ayed et al., 2006) and are reported to be of reduced risk for the environment (Errampalli, 2004), the present study focused on extend of their in vitro and in vivo efficacy evaluation against the development of the entire Fusarium complex causing potato tuber dry rot in Tunisia.
MATERIALS AND METHODS
Pathogens: F. solani, F. graminearum, F. sambucinum and F. oxysporum f.sp. tuberosi are isolated (on 2001, 2002, 2003 and 2004) from tubers of cv. Spunta showing typical symptoms of dry rot. Isolates of F. sambucinum (FRS1, FRS2, F.3/2.02, F.6.02, F.20.02, F.44.03, F.48.02 and F.17.04) implicated in this study are shown to be resistant to benomyl, carbendazim and thiophanate-methyl and all isolates of F. graminearum (F.10/2.02, F.21.02 and F. 45.03), F. oxysporum f.sp. tuberosi (F.33.02) and F. solani (F.12.03) are susceptible to these fungicides (Daami-Remadi and El Mahjoub, 2006).
Fusarium spp. are grown at 25°C on PDA for one week. They are stored at -20°C in 20% glycerol solution for long term preservation.
Potato cultivars: Tubers cv. Spunta, the most cultivated in Tunisia, are used in this current study. They are obtained, on 2004, from the Technical Centre of Potato of Tunisia. For laboratory experiments, tubers are stored in the darkness at 6°C and brought to room temperature three hours before use.
Fungicides: Active ingredient components of tested fungicides are shown to be efficient against several isolates of F. oxysporum f.sp. tuberosi causing potato vascular wilt in previous studies (Ayed et al., 2006). Main characteristics of the tested fungicides are presented in Table 1.
In vitro activity of tested fungicides against Fusarium spp: Fungicides are dissolved in sterile distilled water before their incorporation (1%v/v), following chosen doses (Table 1), in PDA in surfusion. A culture media added with a same quantity of sterile distilled water serves as untreated control. After solidification, agar discs (of 6 mm in diameter) colonized by the tested pathogen are placed at four equidistant emplacements in the centre of the petri dish (four agar discs per petri dish).
| Table 1: | Characteristics and applied doses of fungicides tested against the four Fusarium species causing potato tuber dry rot |
 | |
Inhibitory activity of fungicides is evaluated on mycelial growth of tested Fusarium spp. estimated via mean colony diameter formed after 4 days of incubation at 25°C.
Statistical analyses (ANOVA) are performed following a completely randomised factorial design where treatments (fungicides and untreated control) and Fusarium isolates are both fixed factors. Means are separated using Fisher’s protected LSD test (p≤0.05).
In vivo activity of tested fungicides against Fusarium spp: Efficacy of fungicides previously tested in vitro was estimated via development of dry rot on inoculated and treated tubers. Tubers (cv. Spunta) are superficially disinfected with a solution of 10% sodium hypochlorite, for 5 min and then rinsed abundantly with sterile distilled water. Container and alveolus plaques used for inoculated tubers incubation, are washed before use, dipped for 24 h in sodium hypochlorite solution then rinsed with sterile distilled water.
Fungicides are suspended in water according to tested doses and tuber treatment was realized by dipping tubers, during 10 min, in a fungicidal suspension prior inoculation. Inoculation technique consists of deposing an agar disc (6 mm diameter) colonized by pathogen at occasioned wounds (6 mm diameter and depth). Tuber incubation is realized at 25-27°C for 21 days at high relative humidity. Every elementary treatment is repeated twenty times (ten tubers x two wounds). After incubation period, tubers were cut longitudinally via sites of inoculation. Parameters of dry rot induced (maximal width (w) and depth (d)) are noted. The pathogen penetration within tubers is calculated following formula proposed by Lapwood et al. (1984) where:
Penetration (mm) = (w/2 + (d -6))/2 |
Statistical analyses (ANOVA) are performed following a completely randomised factorial design where treatments (fungicides and untreated control) and Fusarium isolates are both fixed factors. Means are separated using Fisher’s protected LSD test (p≤0.05).
RESULTS
Effects of fungicides on mycelial growth of Fusarium spp: The effect of some fungicides, incorporated in the culture media, are tested against in vitro development of Fusarium spp.
| Table 2: | Effect of some fungicides incorporated into culture media PDA, on mycelial growth of several isolates, belonging to four Fusarium species, as measured by the mean colony diameter (cm) noted after 4 days of incubation at 25°C |
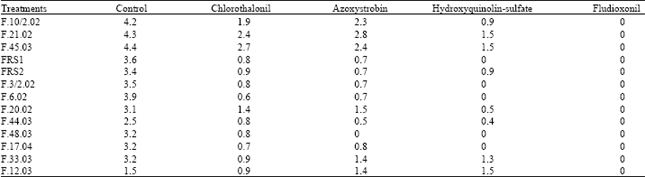 | |
| LSD (Treatments x Isolates of Fusarium sp.) = 0.26 cm (p≤0.05). F.10/2.02, F.21.02 and F.45.03: isolates of F. graminearum, FRS1, FRS2, F.3/2.02, F.6.02, F.20.02, F.44.03, F.48.03 and F.17.04: isolates of F. sambucinum, F.33.03: F. oxysporum f.sp. tuberosi and F.12.03: F. solani | |
| Table 3: | Effect of some fungicides on dry rot development occasioned by Fusarium species as measured by the mean pathogen penetration (mm) into inoculated tubers noted after 21 days of incubation at 25-27°C |
 | |
| LSD (Treatments x Fusarium sp.) = 3.4 mm (p≤0.05), F.45.03: F. graminearum, F.17.04: F. sambucinum, F.33.03: F. oxysporum f.sp. tuberosi and F.12.03: F. solani | |
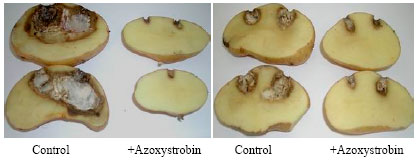 | |
| Fig. 1: | Effect of azoxystrobin on incidence of dry rot occasioned by F. graminearum (left) and F. solani (right), in comparison to inoculated untreated controls, on potato tubers cv. Spunta noted after 21 days of incubation at 25-27°C |
Table 2 showed that mean colony diameter, formed after 4 days of incubation at 25°C, varied upon tested Fusarium isolates and treatments revealing existence of a significant interaction (at p≤0.05) between both fixed factors.
All tested Fusarium isolates (three of F. graminearum, eight of F. sambucinum, one of F. oxysporum f.sp. tuberosi and one of F. solani) showed differential susceptibility to fungicides which significantly reduced pathogen mycelial growth comparatively to the untreated controls.
Fludioxonil showed higher efficacy and totally inhibited the mycelial growth of all tested Fusarium isolates, even those of F. sambucinum which are shown to be resistant to benzimidazoles in previous studies (Daami-Remadi and El Mahjoub, 2006). Table 2 revealed that chlorothalonil, azoxystrobin and hydroxyquinolin-sulfate have inhibited, by more than 70%, the mycelial growth of the majority of tested F. sambucinum isolates comparatively to untreated controls. Their interaction with F. solani, F. graminearum and F. oxysporum f.sp. tuberosi was different; noted inhibition varied from 0 to 79% depending on Fusarium species and different isolates within the same specie. In fact, F. graminearum inhibition varied from 37% to 55% among tested isolates in the case of chlorothalonil. However, lesser growth reduction was noted in F. solani (F.12.03) where the maximum reached inhibition was of about 33%.
Effects of fungicides on Fusarium sp. agressivity on potato tubers: Table 3 showed that mean pathogen penetration noted after 21 days of incubation at 25-27°C varied upon Fusarium sp. used for tuber inoculation and different tested treatments; a significant interaction was noted between both fixed factors (p≤0.05).
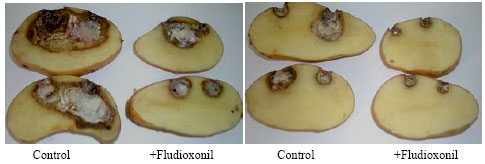 | |
| Fig. 2: | Effect of fludioxonil on incidence of dry rot induced by F. graminearum (left) and F. sambucinum (right), in comparison to inoculated untreated controls, on potato tubers cv. Spunta observed after 21 days of incubation at 25-27°C |
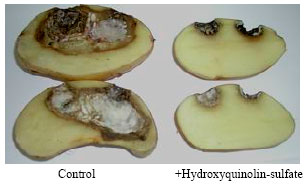 | |
| Fig. 3: | Effect of hydroxyquinolin-sulfate on incidence of dry rot induced by F. graminearum, in comparison to the inoculated untreated control, on potato tubers cv. Spunta, observed after 21 days of incubation at 25-27°C |
In vivo development of F. graminearum (F.45.03), the most aggressive Fusarium species at these incubation conditions, was inhibited by more than 50% by azoxystrobin (Fig. 1), hydroxyquinolin-sulfate (Fig. 3) and fludioxonil (Fig. 2). However, the interaction of the other Fusarium species with these fungicides was slightly different (Fig. 1 and 2); the development of F. sambucinum (F.17.04) and F. oxysporum f.sp. tuberosi (F.33.03) regressed by 40 to 60% by an azoxystrobin treatment.
DISCUSSION
Potato tuber dry rot is a post harvest disease with an increasingly importance in Tunisia. The complex of Fusarium species involved in disease development, its agressivity on tubers and potato plants also as wilting agents (Daami-Remadi and El Mahjoub, 2004) and its survival in fields justified necessity of a tuber treatment in addition to respect of prophylactic methods. The appearance of benzimidazole resistant isolates of F. sambucinum incited us to search for other alternatives.
The present study revealed efficacy of fludioxonil, azoxystrobin and hydroxyquinolin-sulfate against major tested Fusarium isolates. These fungicides have never been tested for F. graminearum, F. sambucinum, F. solani control in Tunisia; they are previously tested only on F. oxysporum f.sp. tuberosi isolates and have shown to be efficient in reducing the Fusarium wilt incidence on inoculated treated plants. Our finding concerning efficacy of fludioxonil joins other studies showing induced mycelial growth inhibition of certain Deuteromycetes fungi achieved by this fungicide (Rosslenbroich and Stuebler, 2000). Fludioxonil, as fenpiclonil, belongs to the phenylpyrrole group and was also shown to have an inhibitory activity against Botrytis cinerea, Monilia sp. and Sclerotinia sp. (Gullino et al., 2000). Leadbeater and Kirk (1992) found that fenpiclonil, is more efficient as a pre-planting treatment than thiabendaole or imazalil. Furthermore, strains showing reduced sensitivity to carbendazim, diethofencarb or vinchlozolin did not show cross resistance with fludioxonil (Forster and Staub, 1996) and resistance to phenylpyrroles has never been observed in the field (Baroffio et al., 2003). In the current study, fludioxonil showed higher efficacy by totally inhibiting the mycelial growth of all tested Fusarium spp. isolates, including those of F. sambucinum resistant to benzimidazoles. When applied at 2 ppm, fludioxonil has completely inhibited the in vitro development of F. sambucinum isolates resistant to thiabendazole.
Similar results were obtained by Bains et al. (2001) when tubers are treated by 50 ppm of imazalil. These authors also found that fludioxonil, additionned by mancozeb or difenoconazol, applied as pre-plantation treatment of tuber seeds inoculated by F. sambucinum, was efficient. In the same way, fludioxonil, tested against Penicillium expansum, a post-harvest apple pathogen, has inhibited the mycelial growth of isolates sensitive and thiabendzole resistant (Errampalli and Crnko, 2004; Errampalli et al., 2004).
Chlorothalonil, azoxystrobin and hydroxyquinolin-sulfate have inhibited by more than 70% the mycelial growth of the majority of F. sambucinum isolates comparatively to the untreated control. Their interaction with the isolates of F. solani, F. graminearum and F. oxysporum f.sp. tuberosi was different; noted inhibition varied from 0 to about 80% depending of Fusarium species and different isolates within same specie. This result joins in part that of Gullino et al. (2000) and D’Mello et al. (2001) who reported that azoxystrobin showed higher efficacy and selectivity for the control of the Fusarium wilt of several plants. Activity of azoxystrobin against benzimidazole resistant isolates of F. sambucinum joins, in part, findings of Schutte et al. (2003) concerning the application of this fungicide for the control of benomyl-resistant Guignardia citricarpa.
The development of F. graminearum, the most aggressive pathogen at inoculation and incubation conditions, and at a lesser degree the other Fusarium species was inhibited in vivo by more than 50% by azoxystrobin, hydroxyquinolin-sulfate and fludioxonil treatments. This is the first report in Tunisia of reduced dry rot development on inoculated potato tubers due to the fludioxonil, azoxystrobin and hydroxyquinolin-sulfate inhibitory activity. Furthermore, as these fungicides are shown to inhibit mycelial growth of benzimidazole resistant isolates of F. sambucinum, they may be implicated in an anti-resistance strategy for dry rot control.
ACKNOWLEDGMENTS
Authors thank the Technical Potato Centre of Tunisia (CTPT) for their financial contribution. Many thanks for Aymen Youssef for his excellent technical assistance.
REFERENCES
- Ayed, F., M. Daami-Remadi, H. Jabnoun-Khiareddine, K. Hibar and M. El-Mahjoub, 2006. Evaluation of fungicides for control of Fusarium wilt of potato. Plant Pathol. J., 5: 239-243.
CrossRefDirect Link - Bang, U., 1992. Influence of seed tuber infestation, chemical seed treatment and pre-harvest climate on incidence of gangrene and dry rot of potato (Solanum tuberosum L.). Potato Res., 35: 3-15.
CrossRefDirect Link - Barofflo, C.A., W. Siegfried and U.W. Hilber, 2003. Long-term monitoring for resistance of Botryotinia fuckeliana to anilinopyrimidine, phenylpyrrole and hydroxyanilide fungicides in Switzerland. Plant Dis., 87: 662-666.
Direct Link - Carnegie, S.F., A.D. Ruthven, D.A. Lindsay and T.D. Hall, 1990. Effects of fungicides applied to seed potato tubers at harvest or after grading on fungal storage diseases and plant development. Ann. Applied Biol., 116: 61-72.
Direct Link - Carnegie, S.F., A.M. Cameron, D.A. Lindsay, E. Sharp and I.M. Nevison, 1998. The effect of treating seed potato tubers with benzimidazoles, imidazole and phenylpyrrole fungicides on the control of rot and skin blemish diseases. Ann. Applied Biol., 133: 343-363.
Direct Link - Daami-Remadi, M. and M. El Mahjoub, 2006. Presence in Tunisia of Fusarium sambucinum isolates resistant to Benzimidazoles: In vitro growth and aggressiveness on potato tubers. Biotechnol. Agron. Soc. Environ., 10: 7-16.
Direct Link - Daami-Remadi, M., H. Jabnoun-Khiareddine, F. Ayed and M. El-Mahjoub, 2006. Effect of temperature on aggressivity of Tunisian Fusarium species causing potato (Solanum tuberosum L.) tuber dry rot. J. Agron., 5: 350-355.
CrossRefDirect Link - Daami-Remadi, M., F. Ayed, H. Jabnoun-Khiareddine, K. Hibar and M. El Mahjoub, 2006. In vitro, in vivo and in situ evaluation of fungicides tested individually or in combination for the control of the fusarium dry rot of potato. Int. J. Agric. Res., 1: 564-572.
CrossRefDirect Link - D`Mello, J.P.F., A.M.C. Macdonald and R. Rinna, 2001. Effects of azoxystrobin on mycotoxin production in a carbendazim-resistant strain of Fusarium sporotrichioides. Phytoparasitica, 29: 431-440.
CrossRefDirect Link - Errampalli, D. and H.W. Johnston, 2001. Control of tuber-borne black scurf Rhizoctonia solani and common scab Streptomyces scabies of potatoes with a combination of sodium hypochlorite and thiophanate-methyl preplanting seed tuber treatment. Can. J. Plant. Pathol., 23: 68-77.
Direct Link - Errampalli, D., 2004. Effect of fludioxonil on germination and growth of Penicillium expansum and decay in apple cvs. Empire and Gala. Crop Prot., 23: 811-817.
CrossRefDirect Link - Forster, B. and T. Staub, 1996. Basis for use strategies of anilinopyrimidine and phenylpyrrole fungicides against Botrytis cinerea. Crop Prot., 15: 529-537.
CrossRefDirect Link - Gullino, M.L., P. Leroux and C.M. Smith, 2000. Uses and challenges of novel compounds for plant disease control. Crop Prot., 19: 1-11.
CrossRefDirect Link - Kawchuk, L.M., J.D. Holley, D.R. Lynch and R.M. Clear, 1994. Resistance to thiabendazole and thiophanate-methyl in canadian isolates of Fusarium sambucinum and Helminthosporium solani. Am. Potato J., 71: 185-192.
CrossRef - Leach, S.S. and L.W. Nielsen, 1975. Elimination of fusarial contamination on seed potatoes. Am. Potato J., 52: 211-218.
CrossRefDirect Link - Leadbeater, A.J. and W.W. Kirk, 1992. Control of tuber borne diseases of potatoes with fenpiclonil. Brighton Crop Prot. Conf.-Pests Dis., 2: 657-662.
Direct Link - Priou, S. and M. El Mahjoub, 1999. Bacterial and fungal diseases in the major potato-growing areas of Tunisia. EPPO Bull., 29: 167-171.
CrossRefDirect Link - Rosslenbroich, H.J. and D. Stuebler, 2000. Botrytis cinerea-history of chemical control and novel fungicides for its management. Crop. Prot., 19: 557-561.
Direct Link - Schutte, G.C., R.I. Mansfield, H. Smith and K.V. Beeton, 2003. Application of azoxystrobin for control of benomyl-resistant Guignardia citricarpa on valencia oranges in South Africa. Plant Dis., 87: 784-788.
Direct Link - Tisdale, M.J. and K.A. Lord, 1973. Uptake and distribution of thiabendazole by seed potatoes. Pesticide Sci., 4: 121-130.
CrossRefDirect Link - Tivoli, B., A. Deltour, D. Molet, P. Bedin and B. Jouan, 1986. Isolation of thiabendazole-resistant strains of Fusarium roseum var. sambucinum from potato tubers. Agronomie, 6: 219-224.
CrossRefDirect Link