
T. Amein
Department of Cell and Molecular Biology, Goteborg University, Goteborg, Sweden
Plant Pathology Journal
Year: 2006 | Volume: 5 | Issue: 3 | Page No.: 356-361







ABSTRACT
Using soil bioassays and isolations from field plants to survey soil-borne fungi causing sugar beet diseases in commercial fields in southern Sweden, damping-off pathogens, Aphanomyces cochlioides, Pythium sp. and Rhizoctonia solani, were the main pathogens found. Of the 1260 plants sampled from 28 fields, 8.5% were infected by these pathogens. Of 1120 seedlings grown in greenhouses in soil from these fields, 6.4% were infected by A. cochlioides and 10.7% were infected by Pythium sp. P. sylvaticum (for first time reported as a pathogen of sugar beet in Sweden). P. ultimum and A. cochlioides were also frequently isolated and were pathogenic in all tests. R. solani AG-2-1 was occasionally isolated and Fusarium sp. were frequently isolated, but were in most cases not pathogenic to test plants. Field soil sampling, seedling sampling, isolations and symptom assessments all indicated that Pythium sp. was predominantly the cause of pre-emergence damping-off, but diseased seedlings in later stages were infected mainly by A. cochlioides. In general, historically high-yielding fields in the area had less seedling diseases and damping-off problems than average yielding fields.
PDF Abstract XML References
How to cite this article
T. Amein, 2006. Soil-Borne Pathogens Infecting Sugar Beet in Southern Sweden. Plant Pathology Journal, 5: 356-361.
DOI: 10.3923/ppj.2006.356.361
URL: https://scialert.net/abstract/?doi=ppj.2006.356.361
DOI: 10.3923/ppj.2006.356.361
URL: https://scialert.net/abstract/?doi=ppj.2006.356.361
INTRODUCTION
Soil-borne fungal pathogens are often responsible for poor establishment and stand loss in sugar beet (Beta vulgaris L.) (Buchholtz, 1938; Harveson and Rush, 1998; Kiewnick et al., 2001; Rush and Winter, 1990; Weiland and Sundsbak, 2000). They also reduce yield and sugar content (Duffus and Ruppel, 1993; Harveson and Rush, 2002; Windels and Lamey, 1998). Pathogens within the genera Pythium, Aphanomyces, Rhizoctonia and Fusarium are commonly present in sugar beet fields. A. cochlioides and R. solani (in some areas) are considered to be the most economically significant pathogens (Martyn, 1989; Payne et al., 1994). At least 12 species of Pythium have also been associated with diseases of sugar beet seedlings (O’Sulivan and Kavanah, 1992). Pythium pathogens, alone or in combination with A. cochliodes, are significant causes of seedling loss in North and South America (Papavias and Ayres, 1974) and in Japan (Yamaguchi, 1977). The symptoms of seedling diseases caused by Pythium sp., A. cochlioides and R. solani, are often similar. These species are common soil inhabitants often found together in complexes. In addition to the level and quality of their soil inocula, many other factors, such as climate conditions (especially temperature and moisture), susceptibility of the host-plant variety, sowing time and effectiveness of control measures have a bearing on disease development in the field (Windels, 1988).
Sugar beet has been grown in Sweden for more than 150 years. In southern Sweden, about 60 000 ha are now sown with sugar beets every year (90% of the fore-crops are cereals). Årrhenius (1923) wrote, Root rot of sugar beets, with which are associated the three fungi Pythium de Baryanum, Aphanomyces levis and Phoma betae, has caused considerable damage in Sweden during the last few years. To my knowledge, since 1923 when this was written, not much has happened with the subject of sugar beet diseases and seedling diseases in particular, for which knowledge remains inadequate, despite the long history of cultivation. Some investigations do highlight the problem of soil-borne pathogens, but without identification of Pythium species and Rhizoctonia sp. within the Anastomosis group (Olsson, 2001, 2002; Ewaldz, 1992; Runeson, 1990). Such omission is a drawback, especially for extension specialists, as shown by a disease survey that was carried out during two successive growing seasons. The survey was conducted as part of a larger research project concerning sugar beet yield constraints, in which 14 sugar beet field pairs were investigated, each pair located near the other, with similar conditions of climate, soil type, soil pH and rotations between crops, but with different yields. One field in each pair was on a farm with a history of average-yielding sugar beet production for the area. The other field was a farm with a history of high-yielding sugar beet production (Blomquist, 1998).
The main purpose of this study was (1) to establish which soil-borne seedling pathogens contributed significantly to plant and yield losses in the area surveyed and (2) to estimate differences in pathogens level and occurrence between high and average yielding fields.
MATERIALS AND METHODS
Soil and plant sampling and fungal isolation
Soil samples: Soil samples were collected with a spade, to a depth of 15 cm, from 14 sugar beet fields in July 1999 and in May 2000. (In southern Sweden, sugar beets are sown about the middle of April). Each field was divided into three parts and in each part the sampled area was 20x20 m. From each field, 15 to 20 sub-samples were taken randomly from positions close to plants. After the sub-samples were mixed, the soils were sent to the laboratory by train, (Plant Pathology and Biocontrol Unit, Swedish University of Agricultural Sciences, where the work was carried out). Soils were sieved (2 mm) and tested within a few days, in a greenhouse soil bioassay.
Soil bioassay: In the first season bioassay, four plastic pots (9 cm diameter) were each filled with soil (400-500 g) from each of the 14 fields. Four pots filled with commercial peat mixture were used as control. Five sugar beet seeds were sown at 2 cm depth in each pot. Pots were then placed in a green house with a day temperature of 22-25°C and a night temperature of 16-20°C for a minimum of 4 weeks. Pots were watered daily. Seedlings with damping-off symptoms were removed, rinsed briefly in tap water and tested for the presence of pathogens by culturing on Potato Dextrose Agar (PDA), Corn Meal Agar (CMA) and were incubated in a shallow layer of sterile distilled water in petri dishes at 20-22°C for 24-72 h. Microscopic identification of fungal colonies and pure isolations for pathogenicity tests were done after 2-7 days. Pure fungal isolates were stored on PDA and CMA slants in tubes in a refrigerator.
In year two, because of the large number of pots used in this experiment, for each field the soil samples were mixed with an equal volume of sand and 60 plastic pots (6 cm diameter) were each filled with 300 g of the soil-sand mixture. To avoid the risk of spreading infection among seedlings, in each pot only one sugar beet seed was sown, at a 2 cm depth. Sixty pots per field were placed on greenhouse benches: the temperature range for 30 pots was 20-28°C and for the other 30 pots, 10-16°C, to allow infection by different pathogens. In each greenhouse test, 30 pots filled with a mixture of sand and the commercial peat, were sown as control. Observation and isolation of the pathogens were as described earlier.
Plants collection: In most cases, the variety Loke was planted. Plants were collected from the fields for symptom readings and fungal isolations. For both growing seasons surveyed, plants at different growing stages, from seedlings to maturity, were randomly collected from each field in the same area from which soils were collected. Plants (20 in year 1 and 30 in year 2) were dug up from each field, placed in plastic bags and directly transported to the laboratory. Plants were washed under running tap water and then small root pieces were placed on PDA and CMA containing streptomycin sulphate (50 μg mL-1). The petri dishes with root pieces were incubated at 20-22°C for 3-7 days at a laboratory bench. Diseased seedlings were incubated in a shallow layer of sterile distilled water as previously described.
Pathogenicity tests: Several hundred fungi, isolated from field-collected seedlings or from soil-assayed seedlings in the greenhouse, were tested for pathogenicity to sugar beet. Mycelial inoculum was grown on PDA in petri dishes at 20-22°C for 7 days. One culture of each isolate was macerated in 20 mL distilled water and then mixed with 400 g of commercial peat mixture, which was used to fill a plastic pot (9 cm diameter). Each such pot was sown with five mono-germ sugar beet seeds. Pots with non-infested soil were used as controls. The sown pots were placed in a greenhouse in which the minimum temperature was 16°C and the maximum, 24°C. The plants were watered every day and when needed, extra light (Philips HPI-T mercury lamps, 400 W) was supplied. Pre-emergence and post-emergence damping-off was recorded daily. Seedlings that survived after 3-4 weeks were removed; roots were washed under tap water; and disease severity was scored. Isolates graded as strong, moderate, or weak pathogens.
Pathogens causing pre-emergence for 70-100% of the test seedlings and/or post-emergence damping-off were considered strong. Those for which 80-100% of the test seedlings emerged, but 50% or more showed clear symptoms (dark brown, water-soaked seedlings, black roots and hypocotyls), were considered moderate. Those for which 100% of the test seedlings emerged, with some showing only weak symptoms (brown to dark brown discoloration of the roots) were considered weak.
Fungal identifications: Strongly and moderately pathogenic isolates were identified using classical methods, viz., colony appearance, growth rate and morphological characters as observed in the light microscope. For identification of R. solani anastomosis groups, the region sequenced was the entire internal transcribed spacer 1 and 2 including 5.8s rRNA gene. the primes used were ITS1 and ITS4 as forward and reverse primers, respectively. Heterothalic isolates of Pythium also were paired on CMA medium.
For identification of the various fungi in the study, the following taxonomic keys were used:
| A. cochlioides | (Scot, 1961) A monograph of the genus Aphanomyces. |
| Pythium sp. | (Dick, 1990) Keys to Pythium and also Van Der Plaats-Niterink (1981). Monograph of the genus Pythium. |
| R. solani | (Sneh et al., 1991) Identification of Rhizoctonia Species. |
| Fusarium sp. | (Both, 1971) The Genus Fusarium and also Nelson et al. (1983); Fusarium species. |
Statistical analyses: For interpretation of data, analysis of variance (ANOVA) was used, with Sources and amounts of variation compared using an F ratio test. To compare treatment means, Least Significant Differences (LSD) test at p≤0.05 were calculated. Means followed by different letters demonstrated a significant statistical difference.
RESULTS
Pathogen isolations and pathogenicity tests: Only A. cochlioides and the various Pythium sp. regularly attacked seedlings in the fields, under the conditions of the soil bioassay. P. sylvaticum, which had not previously been reported on sugar beets in Sweden, was the most common Pythium pathogen, followed by P. ultimum, whereas R. solani was seldom isolated. By doing blast search on the Gen Bank, these isolates showed the most similarity to the sequences of AG2-1 group. Although Fusarium sp. was frequently isolated, most of the isolates were none-pathogenic to sugar beet seedlings.
Soil bioassay: In pooling results from both years surveyed, a total of 10.7% of the 1120 seeds sown in the sampled fields soils were infected by Pythium sp. and 16.4%, by A. cochlioides.
Of 280 seedlings in the year 1 bioassay, two weeks after sowing, 18.9% were infected by Pythium sp. and 17.7%, by A. cochlioides. During the next two weeks, the percentage of seedlings infected by A. cochlioides significantly increased to 44.3% (Fig. 1A).
Temperature had a strong effect on seedlings infected by these two pathogens. In the soil samples tested at 20-28°C, A. cochlioides was isolated from 11.9% of the seedlings, while at the lower greenhouse temperature range of 10-16°C, the pathogen was isolated from only 1.9% of the seedlings. On the other hand, Pythium sp. were isolated from 14.8% of the seedlings at the lower temperature, while at the higher temperature, only 1.2% of the seedlings yielded Pythium sp. (Fig. 1B).
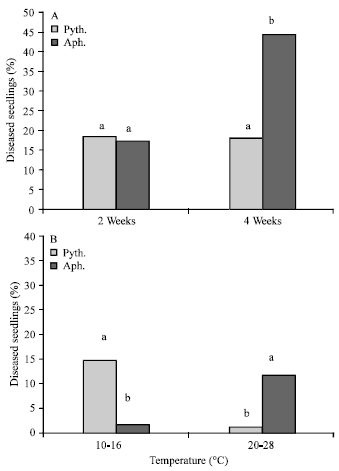 | |
| Fig. 1: | Percent infection of sugar beet seedlings by Pythium sp. and by A. cochlioides in the green house soil bioassay, 2 and 4 weeks after sowing, respectively and under two different temperature conditions (A) 10-16°C and (B) 20-28°C |
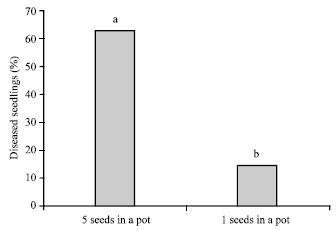 | |
| Fig. 2: | Percent infection of sugar beet seedlings by Pythium sp. and by A. cochlioides in the green house soil bioassay when five and one seeds, respectively, were sown in each pot |
The number of seedlings per pot for soil bioassays in greenhouse tests also had a strong effect on the percentage of seedlings infected.
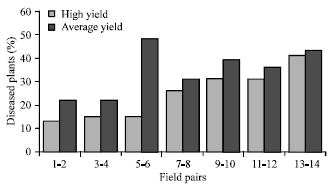 | |
| Fig. 3: | Percent infections of A. cochlioides and Pythium sp. in seedlings grown in green houses and seedlings and mature plants sampled from the average and the high-yielding pairs fields surveyed (80 seedlings in the greenhouse assay and 90 seedlings and mature plants from field-collection were tested for each field) |
When 5 sugar beet seeds were sown in each pot, infection spread among the plants, resulting in 63.2% of the seedlings being infected by Pythium sp. and A. cochlioides, whereas only 14.9% of the seedlings became infected when just 1 seed was sown per pot (Fig. 2).
Pathogens isolations from sampled field plants: During the two growing seasons surveyed, a total number of 1260 field plants from the seedling stage to the maturity were sampled from 28 fields and were then tested for presence of pathogenic fungi. Of these plants, 8.5% were infected by the damping-off pathogens, Pythium sp. and/or A. cochlioides. Pythium was isolated only from seedlings.
Occurrence of damping-off pathogens in high and average fields: The average occurrence of the damping-off pathogens (Aphanomyces and Pythium) was significantly less in high-yielding fields than in average-yielding fields (Fig. 3). Of a total number of 1190 seedlings grown in soil or sampled from average-yielding fields, 20.2% were infected by damping-off pathogens, whereas an average of 14.4% of the same number of seedlings sampled from the high-yielding fields were infected.
DISCUSSION
The dominating seedling pathogens encountered in this survey were zoosporic fungi within the species Pythium and Aphanomyces. These findings correspond well with other investigations in Western Europe. Payne et al. (1994) found A. cochlioides in 39% and Pythium sp. in 31% of 341 British sugar beet fields. Aphanomyces commonly causes damping-off at the 2-8 leaf stage (2-5 weeks old plants) (Jacobsen et al., 2000; Windels, 1988), while P. ultimum may colonize the seed pericarps of sugar beet within 4 h (Osburn et al., 1989). In our survey, A. cochlioides was mainly responsible for later infections and for causing post-emergence damping-off, while Pythium sp. attacked plants before or soon after seedling emergence (Fig. 1A). These two pathogens often are the main cause of poor stand establishment, due to pre-emergence and post-emergence damping-off, but they may also cause severe reduction of sugar beet yields (Baker and Rush, 1988; Rush, 1987). In 1999, about 51% of over 293,000 ha of land sown to sugar beet in the states of Minnesota and North Dakota (United States) was infested with A. cochlioides (Beale et al., 2002). In the state of Montana, stand losses from Aphanomyces of 10-50% are common (Jacobsen et al., 2000) and Pythium has been reported to kill more than 90% of the growing seedlings (Hendrix and Campbell, 1973).
Aphanomyces and Pythium pathogens often occur together in sugar beet fields, but the dominating pathogens can belong to one or the other of these genera, depending on environmental factors such as soil moisture and temperature. In our greenhouse test, using host bait plants, A. cochlioides infections were significantly more frequent at higher temperatures (20-28°C) than at lower temperatures (10-16°C): that is, 11.9 and 1.9%, respectively (Fig. 1B). This pathogen is known to cause damping-off in warm (20-30°C), wet soils (Whitney and Duffus, 1986; Windels and Lamey, 1998), but at soil temperatures less than 15°C, infection seldom occurs (Windels and Lamey, 1998). On the other hand, Pythium sp. were isolated more frequently at lower greenhouse temperatures (10-16°C) (Fig. 1B), which agrees with other reports (O’Sullivan and Kavanagh, 1992; Schmidt et al., 2004). Since at least 12 species of Pythium have been associated with diseases of sugar beet seedlings (O’Sullivan and Kavanagh, 1992), the variation in favorable temperatures might be high among various species. In our soil samples, P. sylvaticum was the most frequent and widely distributed species attacking sugar beet seedlings, similar to reports from Ireland (O’Sullivan and Kavanagh, 1992). The second most frequently isolated species was P. ultimum. This study happens to be the first to report P. sylvaticum as a sugar beet pathogen in Sweden, confirmed by Persson (2003) who isolated and identified this pathogen in the same fields afterwards.
The simple bating methods used in these studies were suitable for arranging soils relative to each other, but they gave less reliable values concerning amounts of inocula, or the inoculum potentials for Pythium sp. and A. cochlioides. In addition to being very sensitive to temperature and watering, these methods were highly dependent on the number of plants sown per pot. A much higher percentage of infected baiting plants were obtained when 5 seeds per pot were sown than when 1 seed per pot was used (Fig. 2). This might be explained by movement and by spreading of the zoospores of these pathogens in the water, in that infection of nearby seedlings in pots with 5 seeds was observed. Zoospore dispersal occurs through transport in water currents over soil surfaces and also through autonomous movement over mm-or cm-distances (Newhook et al., 1981). Also, planting more seeds per pot increases the odds of roots encountering pathogen inoculum in soil.
We also isolated R. solani and, frequently, Fusarium sp. from sugar beet seedlings. R. solan is often a strong pathogen in sugar beets (O’Sullivan and Kavanagh, 1991) and in certain areas is the economically most important. In Southern Europe, R. solani is regarded a severe beet damaging pathogen. In the United States, R. solani also causes significant economic loses and occurs in more than 25% of the beet production area (Kiewnick et al., 2001). The degree of damage can vary greatly from field to field (as much as 50%), depending on cropping history and environment (Whitney and Duffus, 1986). O’Sullivan and Kavanagh (1991) also reported R. ceralis as a damping-off pathogen of beet seedlings, but few other reports on such infections are available. In our samples, R. solani was isolated less frequently than Pythium sp. and A. cochlioides.
Various Fusarium species were isolated with high frequency; however, most of these isolates were not pathogenic to sugar beet seedlings in the greenhouse. For this reason, the sugar beet pathogens within this genus are considered economically less important than Pythium sp. and A. cochlioides in Southern Sweden, at least in the seedling stage. In the United States, F. oxysporum has often been reported as a pathogen of sugar beet (Harveson and Rush, 1997) and has caused major sugar beet yield losses in Texas in the 1990s (Harveson and Rush, 1998).
There was a clear tendency for the historically high-yielding fields to have less damping-off problems compared to the average-yielding fields in the same area (Fig. 3). Presumably, these differences were mainly due to the influence of factors affecting soil inoculum and disease outbreaks, such as crop rotation, soil texture, soil-pH, time of sowing and cultivar selection. Other soil factors, including soil-type, clay content, organic materials and drainage probably also influence soil-borne pathogens of sugar beet.
ACKNOWLEDGMENT
We thank the Swedish Farmers’ Foundation for Agricultural Research (SLF) for funding this study.
REFERENCES
- Beale, J.W., C.E. Windels and L.L. Kinkel, 2002. Spatial distribution of Aphanomyces cochlioides and root rot in sugar beet fields. Plant Dis., 86: 547-551.
Direct Link - Booth, C., 1971. The Genus Fusarium. Commenwealth Mycological Institute, Kew, Surrey, England, ISBN: 9780851980461, Pages: 237.
Direct Link - Harveson, R.M. and C.M. Rush, 1998. Characterization of Fusarium root rot isolates from sugar beet by crowth and virulence at different temperatures and irrigation regimes. Plant Dis., 82: 1039-1042.
Direct Link - Harveson, R.M. and C.M. Rush, 2002. The influence of irrigation frequency and cultivar blends on the severity of multiple root diseases in sugar beets. Plant Dis., 86: 901-908.
Direct Link - Hendrix, F.F. and W.A. Campbell, 1973. Pythium as plant pathogens. Ann. Rev. Phytopathol., 11: 77-98.
CrossRefDirect Link - Kiewnick, S., B.J. Jacobsen, A. Braun-Kiewnick, J.L.A. Eckhoff and J.W. Bergman, 2001. Integrated control of Rhizoctonia crown and root rot of sugar beet with fungicides and antagonistic bacteria. Plant Dis., 85: 718-722.
CrossRefDirect Link - Martyn, R.D., 1989. Etiology of a root rot disease of sugar beet in Texas. Plant Dis., 11: 879-884.
Direct Link - Nelson, P.E., T.A. Toussoun and W.F.O. Marasas, 1983. Fusarium Species: An Illustrated Manual for Identification. 1st Edn., Pennsylvania State University Press, University Park, University Park, PA., USA., ISBN-13: 978-0271003498, Pages: 226.
Direct Link - Osburn, R.M., M.N. Schroth, J.G. Hancock and M. Hendson, 1989. Dynamic of sugar beet seed colonization by Pythium ultimum and Pseudomonas Species: Effects on rot and damping-off. Phytopathology, 79: 709-716.
Direct Link - O`Sulivan, E. and J.A. Kavanagh, 1992. Characteristics and pathogenicity of Pythium sp. associated with damping-off of sugar beet in Ireland. Plant Pathol., 41: 582-590.
CrossRefDirect Link - Payne, P.A., M.J.C. Asher and C.D. Kershaw, 1994. The incidence of Pythium sp. and Aphanomyces cochliodes associated with the sugar-beet growing soils of Britian. Plant Pathol., 43: 300-308.
Direct Link - Rush, C.M. and S.R. Winter, 1990. Influence of previous crops on Rhizoctonia root crown rot of sugar beet. Plant Dis., 74: 421-425.
CrossRefDirect Link - Schmidt, C.S., F. Agostini, C. Leifert, K. Killham and C.E. Mullins, 2004. Influence of soil temperature and matric potential on sugar beet seedling colonization and suppression of Pythium damping-off by the antagonistic bacteria Pseudomonas fluorescens and Bacillus subtilis. Phytopathology, 94: 351-363.
Direct Link - Weiland, J.J. and J.L. Sundsbak, 2000. Differentiation and detection of sugar beet fungal pathogens using PCR amplification of actin coding sequences and the ITS region of the rRNA gene. Plant Dis., 84: 475-482.
Direct Link - Windels, C.E. and H.A. Lamey, 1998. Identification and Control of Seedling Diseases, Root Rot and Rhizomania on Sugar Beet. BU-7192-S. North Dakota State University, Fargo, pp: 1142.
Direct Link