
H.U. Ahmed
Field Crop Development Centre, Alberta Agriculture,
Food and Rural Development (AAFRD), Lacombe, AB T4L 1W8, Canada
K.F. Chang
Field Crop Development Centre, Alberta Agriculture,
Food and Rural Development (AAFRD), Lacombe, AB T4L 1W8, Canada
S.F. Hwang
Alberta Research Council, Vegreville, AB T9C 1T4, Canada
B.D. Gossen
Agriculture and Agri-Food Canada, Saskatoon Research Centre, Saskatoon, SK S7N OX2, Canada
R.J Howardand
Crop Diversification Centre South, AAFRD, Brooks, AB TlR 1E6, Canada
T.D. Warkentin
Crop Development Centre, University of Saskatchewan,
Saskatoon, SK S7N 5A8, Canada
Plant Pathology Journal
Year: 2006 | Volume: 5 | Issue: 3 | Page No.: 336-342







ABSTRACT
Nineteen chickpea varieties were evaluated for their reaction to ascochyta blight following detached-leaf and whole-plant inoculation with an aggressive isolate of Ascochyta rabiei. The same varieties were also evaluated under field conditions following inoculations with chickpea residues infected with ascochyta blight. A wide range of variation was observed among the varieties for disease severity, stem lesion length, lesion number and pycnidial density. Positive correlations between stem lesion length, pycnidial density and disease severity indicated that stem lesion length or pycnidial density may be an indicator of disease resistance. Among the chickpea entries tested, two varieties with compound leaves (desi cv. Myles, kabuli cv. CDC Chico) were relatively resistant. Blight severity was consistently higher on unifoliate varieties compared to those with compound leaves, irrespective of evaluation methods. These results suggest that avoiding the cultivation of unifoliate-leaved chickpea varieties in the Albertan agro-ecosystems. Instead, breeders need to focus on developing compound-leaved varieties with improved ascochyta blight resistance for sustainable chickpea production.
PDF Abstract XML References Citation
How to cite this article
H.U. Ahmed, K.F. Chang, S.F. Hwang, B.D. Gossen, R.J Howardand and T.D. Warkentin, 2006. Components of Disease Resistance in Desi and Kabuli Chickpea Varieties Against Ascochyta Blight. Plant Pathology Journal, 5: 336-342.
DOI: 10.3923/ppj.2006.336.342
URL: https://scialert.net/abstract/?doi=ppj.2006.336.342
DOI: 10.3923/ppj.2006.336.342
URL: https://scialert.net/abstract/?doi=ppj.2006.336.342
INTRODUCTION
Ascochyta blight of chickpea (Cicer arietinum L.), caused by Ascochyta rabiei (Pass.) Labr. [teleomorph: Didymella rabiei (Kovacheski) v. Ayx], is a devastating disease in many chickpea growing regions of the world, including western Canada (Nene, 1982; Singh et al., 1984; Nene and Reddy, 1987; Chongo and Gossen, 2003; Chang et al., 2000, 2003, 2004; Ahmed et al., 2005). It affects all above-ground parts of the plant, and can cause total loss of the crop (Singh and Reddy, 1996).
Combinations of cultural practices (e.g., crop rotation and tillage, Gossen and Miller, 2004) and foliar fungicides are used to manage ascochyta blight. However, the repeated application of fungicides that is required to control the disease in susceptible cultivars under conditions favorable for disease development is generally not cost effective (Reddy and Singh, 1993). The preferred method of managing ascochyta blight would be development of cultivars with high levels of resistance.
Many resistant cultivars have been developed and released in several countries (Singh and Reddy, 1983), but breakdown of resistance often occurs quickly. Pathogenic and genetic diversity have been documented within the population of A. rabiei in Canada (Chongo et al., 2004). Information on the response of chickpea varieties to local pathogen isolates is a prerequisite for deploying resistance in commercial fields. Researchers have used a variety of screening and assessment techniques to identify sources of resistance (Vir and Grewal, 1974; Gowen et al., 1989; Riahi et al., 1990; Dolar et al., 1994; Chongo and Gossen, 2001; Lichtenzveig et al., 2002; Chang et al., 2004; Chen et al., 2004), but there is no clear consensus on the optimum methodology at this time.
Two types of chickpea, desi (small, angular, tan-colored seed) and kabuli (large, round, beige-colored seed), with either compound or unifoliate leaf type, are grown in western Canada. The objectives of this study were: i) to evaluate the response of chickpea varieties to ascochyta blight under growth chamber, greenhouse and field conditions and ii) to assess the impact of varieties and leaf types on disease development.
MATERIALS AND METHODS
Plant propagation: Nineteen chickpea varieties (Table 1), including desi and kabuli market classes having compound or unifoliate leaves and primarily obtained from the 2001 Saskatchewan-Alberta regional variety trials, were grown in a soil mixture containing vermiculite, peat moss, loam and sand (1: 1.25: 1: 0.75) in 10 cm-diameter pots under greenhouse conditions (temperature 22±1°C; 16 h light and 8 h dark period and 92 μEs-1m-2 light intensity). Three replicates (pots) were used per variety, with two plants per pot. The test was seeded on January 22, 2004 (Trial I). The trial was repeated, with the same varieties, seeded on May 11, 2004 (Trial II). The plants were fertilized three times with a 0.1% solution of 20-20-20 (N-P-K) during the growing period.
Pathogen inoculum preparation: An aggressive isolate (CMG leaf 5-16) of A. rabiei was obtained from a chickpea field severely infected with ascochyta blight in southern Alberta, Canada in 2003.
| Table 1: | List of chickpea cultivars and lines assessed for resistance to ascochyta blight caused by Ascochyta rabiei |
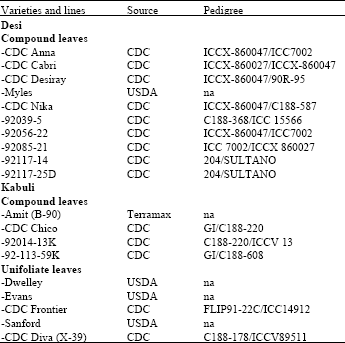 | |
| CDC; Crop Development Centre, University of Saskatchewan, Saskatoon, SK, Canada USDA; United States Department of Agriculture. na; not available | |
For long term storage, the fungus was grown on chickpea-agar medium (20.0 g ground chickpea seed (pre-soaked overnight) and 14.0 g agar L-1) for 7-10 days that yielded profuse pycnidia oozing pycnidiospores. The pycnidiospores were washed with sterile distilled water into a tube for centrifugation, and the pellet was transferred to an Eppendorf (1.5 mL) tube and the culture was stored at -20°C in a freezer. For multiplication of the pathogen, pycnidiospore suspension was prepared with a loop full of the stock culture in sterile water and was spread over the chickpea-agar medium with a glass rod on rotary plate, and the culture was grown for 7-10 days at 20±2°C. During inoculation, the pycnidiospores were transferred to sterile water with a glass slide and filtered through cheesecloth. The concentration of the spore suspension was adjusted to 1x106 spore mL-1 with water and one drop of Tween 80 L-1 as surfactant was added to the suspension before spraying.
Detached-leaf inoculation: Chickpea stems (with leaves attached) were collected from greenhouse-grown plants. Leaves of similar age and size (3-4 leaf positions from the top) were selected from each variety and excised along with 2-3 mm stem on both sides of the nodes with a sharp knife. The leaves were inoculated as previously described (Hwang et al., 2006). The excised leaves were placed inside a small humidity chamber made by lining the bottom of a petri dish with wet paper towel covered with plastic mesh. A cotton plug soaked with nutrient solution was used to cover the cut ends of the stem to maintain leaf hydration. The inoculum of the pathogen was prepared as described before. Five leaflets per compound leaf or five individual unifoliate leaves per replication (experiment replicated three times) were inoculated at a single point with 10 μL of spore suspension containing 1x106 mL-1 pycnidiospore of the pathogen. The leaves were incubated under a 12-h photoperiod at 20°C and >95% relative humidity.
The incubation period (the period between inoculation and first detection of visual symptoms) on each varieties was determined based on daily assessment. Leaf Lesion Diameter (LLD) was assessed three times at 3-day intervals starting 5 days after inoculation, using a digital slide caliper, and the data were used to calculate the area under the disease progress curve (AUDPC). Pycnidial Density (PD) was rated visually using a magnifying glass on a 0-5 scale where: 0 = no pycnidia; 1 = few pycnidia; 3 = moderate numbers of pycnidia and 5 = abundant pycnidia with cirri. The scale to estimate pycnidial density was derived from the one published by Adhikari et al. (2003) to measure pycnidial density in the Mycosphaerella graminicola - Triticum aestivum pathosystem, which produces very similar pycnidia and cirri of pycnidiospores on the disease lesions.
Whole-plant inoculation: Three replicate pots of each variety were inoculated with a conidial suspension (1x106 mL-1) of isolate CMG leaf 5-16 60 days after seeding in Trial I and 24 days after seeding in Trial II. The inoculated plants were covered with plastic bags for 48 h to maintain high humidity. The greenhouse temperature was maintained at 20±2°C. In Trial I, Stem Lesion Length (SLL) for the five longest lesions per pot was recorded 7 days after inoculation and whole-plant Disease Severity (DS) (1-9 scale; Singh et al., 1984) was rated 21 days after inoculation. In Trial II, in addition to SLL and DS, Stem Lesion Number (SLN) was assessed on three stems and Pycnidial Density (PD) on at least three disease lesions per replication were rated 3 weeks after inoculation.
Field trial: The ascochyta blight reaction of the same set of chickpea varieties used in the previous studies was assessed in a field trial at the Crop Diversification Centre South, Brooks, Alberta, Canada. The soil type was a brown chernozem clay loam. The trial was laid out in a randomized complete block design with three replications. Each plot consisted of two, 3-m-long rows, with 30 cm between rows and 2 m between blocks. Prior to seeding, the area was treated with granular herbicide (EdgeTM; ethalfuralin at 17 kg ha-1) for weed control. The experiment was seeded at 10 seeds m-1 row using a push seeder on May 13, 2005. After seeding, the plots were irrigated using a riser and impact sprinkler system to ensure adequate soil moisture for seed germination. The trial was inoculated on August 2, 2005 by spreading ascochyta blight-infected chickpea residue collected from the 2004 crop. Ascochyta blight severity was assessed on a (1-9) scale, described previously, on 10 randomly chosen plants in each plot at 95 and 124 days after seeding.
Data analysis: Analysis of variance of AUDPC, LLD, PD, DS, SLL and SLN data were performed following PROC GLM procedure using SAS Statistical Analysis System (SAS Institute, Cary, NC). Means were compared using Fishers’ Protected Least Significant Difference test. In addition, the varieties were pooled into groups based on phenotype and orthogonal contrasts were made between the following groups: compound vs. unifoliate, desi vs. kabuli, desi-compound vs. kabuli-compound and kabuli-compound vs. kabuli-unifoliate, for each response variable. In addition, Pearson Correlation Coefficients were determined to reveal the relationship among the disease parameters obtained over the trials.
| Table 2: | Analysis of variance of area under the disease progress curve, lesion diameter, pycnidial density, disease severity, stem lesion length for 19 chickpea varieties inoculated with Ascochyta rabiei in detached-leaf, whole-plant tests, and in a field trial at Brooks, Alberta |
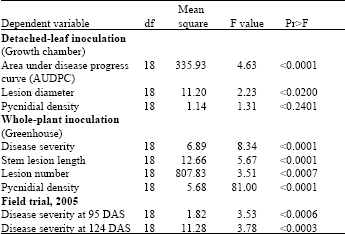 | |
| DAS = Days After Seeding | |
RESULTS
Analysis of variance: Analysis of variance of AUDPC, LLD, PD, DS, SLL and SLN obtained over the trials indicated significant differences among the chickpea varieties, except for PD measured following detached-leaf inoculation (Table 2). The level of significance ranged from p≤0.0001-0.02.
Detached-leaf inoculation: The area under the disease progress curves was the lowest on ‘CDC Desiray’ followed by Amit (B-90), Sanford, Dwelley and Myles, while the highest disease progress was noticed on CDC Diva (Table 3). The leaf lesion diameter ranged from 7-15 mm with CDC Diva having the greatest size and CDC Cabri the smallest. The response of the varieties to pycnidial production was variable (3.0-5.0) on a 0-5 scale. Pycnidial production was observed on CDC Anna, CDC Frontier, CDC Nika, Sanford and 92117-14 3 days after inoculation (data not shown). On the other varieties, the pycnidia were not noticed until at least 5 days after inoculation.
Whole-plant inoculation: Since the analysis of disease severity and stem lesion length noted in both Trials I and II showed a similar pattern of disease reaction to the test varieties, the results of Trial II are presented here. A wide range of variation was detected among the varieties in DS, SLL, SLN and PD (Table 3). Disease severity was the least on Myles (4.3) followed by CDC Chico (4.6), the shortest SLL was observed on CDC Nika (7.6 mm) with the longest on Evans (14.7 mm). The SLN was the least on ‘Myles’ (12) and the highest on line 92056-22 (75). Myles also produced the least pycnidia.
| Table 3: | Reactions of chickpea cultivars and lines inoculated with Ascochyta rabiei following detached-leaf, whole-plant inoculations and in a field trial at Brooks, Alberta in 2005 |
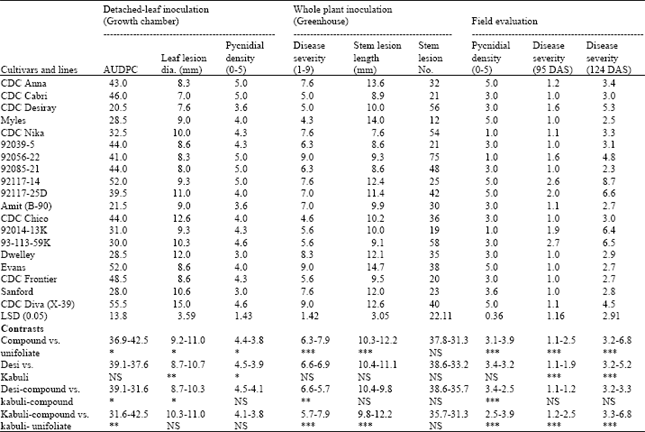 | |
| Data are the mean of three replications. Disease severity was assessed following 1-9 scale and pycnidial density scored following 0-5 scale. LSD = Least significant difference. Chickpea cultivar and lines includes ten desi, four kabuli-compound and five kabuli unifoliate leaf chickpeas (see Table. 1). DAS= Days After Seeding. *, **, ***; indicate level of significance at p≤0.05, 0.01, 0.001, respectively, by orthogonal contrasts (df = 1) between the chickpea phenotype groups. NS = contrast is not significant | |
Field trial: Blight severity was low in the field compared to greenhouse studies, despite a favorable season for ascochyta blight development. However, results indicated significant differences in DS among the chickpea varieties (Table 3). Disease severity at crop maturity ranged from 2.3 to 8.7. The chickpea line 92085-21 had the lowest DS (2.3), followed by Myles (2.5) and Amit. The lines 92014-13K, 93-113-59K, 92117-25D and 92117-14 had disease reaction >5.0 on a 1-9 scale.
Comparison among varieties: Comparison of AUDPC obtained after detached-leaf inoculation indicated significantly higher (16%) disease progress on unifoliate than compound-leaved kabuli, 24% higher on compound-leaved desi than compound-leaved kabuli and 34% higher on unifoliate kabuli than that of the compound-leaved kabuli chickpeas (Table 3). For other contrasts, the differences were not statistically significant.
The LLD was significantly larger on unifoliate, kabuli and compound-leaved kabuli chickpeas compared to desi and compound-leaved kabuli (Table 3). The PD was higher on compound-leaved and desi groups compared to unifoliate and desi in the detached leaf study. It is noteworthy that the desi varieties had higher disease indicators compared to compound-leaved kabuli varieties, except for LLD measurements.
Comparison of blight severity between compound vs. unifoliate leaf types of chickpea indicated unifoliate types had significantly more disease (p≤0.001) in whole-plant trials (Table 3). As a group, kabuli varieties were consistently more susceptible than desi varieties and the group of kabuli varieties with compound-leaves than the group of unifoliate kabuli varieties. Field evaluations showed a similar pattern, although there was variation in disease reactions within each group (compound-leaf, unifoliate, desi and kabuli). Following whole plant inoculation, the SLL on unifoliate varieties was significantly longer (p≤0.001) compared to desi or compound-leaved kabuli chickpea, but other contrasts, e.g., desi vs. kabuli and desi vs. compound-leaved kabuli, were not significantly different (Table 3).
| Table 4: | Pearson correlation coefficients for the chickpea ascochyta blight disease parameters obtained following detached-leaf, whole-plant inoculations and in a field evaluation at Brooks, Alberta, 2005 |
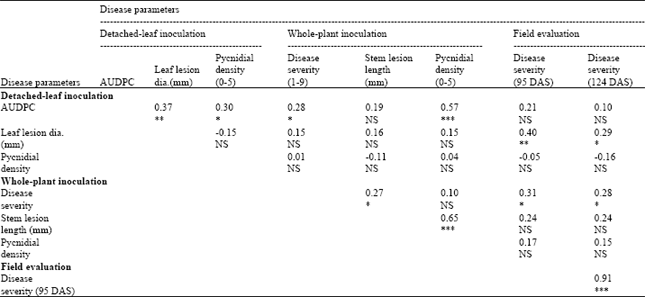 | |
| *, ** and ***; indicates statistically significant at p≤0.05, 0.01 and 0.001, respectively. NS; Not Significant. DAS; Days After Seeding | |
The SLN among the contrasts was not significantly different, although compound-leaved chickpea varieties irrespective of desi or kabuli type produced greater lesion numbers than the unifoliate varieties. The PD was significantly higher on unifoliate than on compound-leaved kabuli, on desi than on compound-leaved kabuli, and on unifoliate kabuli than on compound-leaved kabuli.
Correlation among the disease parameters: Pearson correlation among the disease parameters revealed significant and positive relationships of AUDPC with LLD (following detached-leaf inoculation), PD, DS and PD (whole-plant inoculation). The LLD had a significantly positive correlation with field DS for both the scoring dates. A similar association was obtained with DS (whole-plant inoculation) with SLL and field DS. Among the disease parameters, the highest correlation (r = 0.65; p≤0.001) was obtained between SLL and PD measured under whole-plant greenhouse inoculation (Table 4).
DISCUSSION
The isolate of the pathogen used in this study was virulent on all varieties, although the varieties were significantly different in disease reaction. Chen et al. (2004) characterized chickpea for pathogenicity assay of ascochyta blight on different varieties including Dwelley, Evans, Sanford and Myles. When these varieties were inoculated with pathotypes I and II, pathotype II caused more disease severity than pathotype I. We observed similar levels of disease severity on Dwelley, Evans and Sanford in the present study, which may indicate that the isolate we used belongs to pathotype II. However, severity on Myles was lower than on the other cultivars in the present study.
The present study indicated that the varieties CDC Chico and Myles were moderately resistant compared to the other varieties evaluated based on all the tests. We observed a continuous variation in disease reaction among the varieties tested. Therefore, the varieties may be classified as moderately resistant (4-5), moderately susceptible (6-7) and highly susceptible (8-9). In the absence of high levels of resistance, production of a moderately resistant or moderately susceptible cultivar would likely provide a yield advantage over a highly susceptible line (Singh and Reddy, 1996).
Rate-reducing resistance, also described as partial resistance, to ascochyta blight has been described on different chickpea cultivars (Reddy and Singh, 1993). Rate-reducing resistance is the aggregate of several components of the host-pathogen interaction system that leads to a reduced number of infection sites, reduced lesion expansion, a longer interval required for formation of fungal reproductive structures and reduced sporulation (Parlevliet, 1979). Some of the physiological factors underlying these components have been reviewed recently (Jayakumar et al., 2005). This study demonstrated differences in incubation period (data not shown), lesion length and number and pycnidial density.
Incubation period and pathogen reproduction are very important features in epidemic development under field conditions. An epidemic on a cultivar with a short incubation period and production of abundant pycnidia would develop more quickly than on a cultivar with a longer incubation period or production of few pycnidia.
Significant correlations occurred among the disease parameters including positive correlation of AUDPC with LLD (detached-leaf inoculation), PD, DS and PD (whole-plant inoculation), although a large amount of variation remained unexplained in most cases (Table 4). It is interesting to note that the LLD (detached-leaf inoculation) had a significantly positive relationship with field DS for both scoring dates. The DS (whole-plant inoculation) showed a similar relationship with SLL, and field DS. Significantly higher (r = 0.65; p≤0.001) correlation was obtained between SLL and PD measured under whole-plant greenhouse inoculation. These results suggest that the longer the SLL, the higher the PD and DS. Therefore, PD or SLL may be an indicator to measure the resistance of chickpea varieties. Dolar et al. (1994) reported a strong positive correlation between ascochyta blight severity in a whole-plant inoculation study in a greenhouse and inoculation of detached leaves.
Ascochyta blight severity is greatly influenced by environmental conditions (Chen et al., 2004). We observed much higher levels of DS in the greenhouse compared to the field trial, despite environmental conditions in the field which were conducive for ascochyta blight development. Under the uniformly favorable environmental conditions of the detached-leaf inoculation trial, most varieties were susceptible, although there was variation in susceptibility. Unifoliate leaf type chickpea varieties were more susceptible to ascochyta blight than the compound-leaved lines. Similar results were obtained in field trials including 14 chickpea varieties. Gan and McDonald (2002) reported that this difference was consistent regardless of precipitation, and change of population density.
The compound-leaved type has a morphological advantage over the unifoliate type because it is unlikely that all the leaflets would be infected and infection can spread without interruption in unifoliate leaves. Furthermore, the unifoliate type of leaf is sessile, while the compound leaf has a prominent petiole. Therefore, it is easy for the pathogen to infect the stem of unifoliate plants, thereby cutting off water and nutrients to other parts of the plant. Thus, under similar environmental and disease conditions, the compound-leaved chickpeas can withstand infection more efficiently compared to the unifoliate type.
Present study confirms and complements the findings of Gan and McDonald (2002) by adding information on other components of ascochyta blight resistance including stem lesion length, leaf lesion diameter and pycnidial density under controlled inoculation studies. These findings indicate that compound-leaved chickpea varieties are relatively more resistant than unifoliate types. Sanford, a unifoliate type of chickpea, is predominantly grown in southern Alberta (Ahmed et al., 2005) and was found to be highly susceptible to the isolate of A. rabiei used in our study. Farmers need to be encouraged to avoid growing unifoliate and highly susceptible chickpea cultivars in order to increase their profit margins. Breeding for resistance to ascochyta blight should be targeted at kabuli-type chickpeas with compound leaves.
Furthermore, integration of foliar fungicides with moderately resistant or moderately susceptible cultivars would likely give more cost-effective disease control compared to applying fungicides onto highly susceptible varieties.
ACKNOWLEDGMENTS
The authors are grateful to Robyne Bowness, Dustin Burke, Lindsay Benoit and Trina Dubitz for their assistance in the greenhouse and field trials. This research was funded by the Alberta Agricultural Research Institute, Alberta Crop Industry Development Fund Ltd., and Agriculture and Food Council.
REFERENCES
- Adhikari, T.B., J.M. Anderson and S.B. Goodwin, 2003. Identification and molecular mapping of a gene in wheat conferring resistance to Mycosphaerella graminicola. Phytopathology, 93: 1158-1164.
Direct Link - Chen, W., C.J. Coyne, T.L. Peever and F.J. Muehlbauer, 2004. Characterization of chickpea differentials for pathogenicity assay of ascochyta blight and identification of chickpea accessions resistant to Didymella rabiei. Plant Pathol., 53: 759-769.
Direct Link - Chongo, G. and B.D. Gossen, 2001. Effect of plant age on resistance to Ascochyta rabiei in chickpea. Can. J. Plant Pathol., 23: 358-363.
CrossRefDirect Link - Chongo, G., B.D. Gossen, L. Buchwaldt, T. Adhikari and S.R. Rimmer, 2004. Genetic diversity of Ascochyta rabiei in Canada. Plant Dis., 88: 4-10.
Direct Link - Dolar, F.S., A. Tenuta and V.J. Higgins, 1994. Detached leaf assay for screening chickpea for resistance to ascochyta blight. Can. J. Plant Pathol., 16: 215-220.
Direct Link - Gowen S.R., M. Orton, B. Thurley and A. White, 1989. Variation in pathogenicity of Ascochyta rabiei on chickpeas. Trop. Pest Manage., 35: 180-186.
Direct Link - Jayakumar, P., B.D. Gossen, Y.T. Gan, T.D. Warkentin and S. Banniza, 2005. Ascochyta blight of chickpea: Infection and host resistance mechanisms. Can. J. Plant Pathol., 27: 499-509.
Direct Link - Lichtenzveig, J., D. Shtienberg, H.B. Zhang, D.J. Bonfil and S. Abbo, 2002. Biometric analyses of the inheritance of resistance to Didymella rabiei in chickpea. Phytopathology, 92: 417-423.
Direct Link - Nene, Y.L., 1982. A review of ascochyta blight of chickpea. Trop. Pest Manage., 28: 61-70.
CrossRefDirect Link - Parlevliet, J.E., 1979. Component of resistance that reduce the rate of epidemic development. Ann. Rev. Phytopathol., 17: 203-222.
CrossRefDirect Link - Reddy, M.V. and K.B. Singh, 1993. Rate-reducing resistance to ascochyta blight in chickpea. Plant Dis., 77: 231-233.
Direct Link - Singh, K.B. and M.V. Reddy, 1983. Inheritance of resistance to ascochyta blight in chickpea. Crop Sci., 23: 9-10.
Direct Link - Singh, K.B., M.V. Reddy and Y.L. Nene, 1984. International testing of chickpeas for resistance to ascochyta blight. Plant Dis., 68: 782-784.
Direct Link - Singh, K.B. and M.V. Reddy, 1996. Improving chickpea yield by incorporating resistance to ascochyta blight. Theoret. Applied Genet., 92: 509-515.
CrossRefDirect Link