
Amit Kumar Sehgal
Department of Biosciences, Himachal Pradesh University, Summer Hill, Shimla-171005 India
Anand Sagar
Department of Biosciences, Himachal Pradesh University, Summer Hill, Shimla-171005 India
Plant Pathology Journal
Year: 2006 | Volume: 5 | Issue: 3 | Page No.: 315-321







ABSTRACT
Cordyceps militaris is an entomopathogenic and medicinal fungus. Therefore in vitro isolation and influence of nutritional conditions on the mycelial growth of this fungus was carried out in this study. A small bit of tissue from stipe and stroma was used as starting material for in vitro isolation. The present study revealed that this fungus showed optimum growth in Yeastal Potato Dextrose Agar Medium and Glucose Asparagine Solution. The maximum mycelial growth was observed at 25°C and pH 7.5 and 5.5 in solid and liquid medium, respectively. Among 12 carbon and 9 nitrogen sources tested, the best growth was supported by Sucrose and Ammonium phosphate, respectively. All the four mineral sources supported good growth of the fungus, however 2 ppm calcium chloride and manganese sulphate, ppm ferrous sulphate and 5 ppm ammonium molybdate proved superior as compared to higher or lower concentrations of the same mineral source tried. Among the 4 growth regulators, Gibberellic acid supported the maximum mycelial growth at 100 ppm concentration.
PDF Abstract XML References
How to cite this article
Amit Kumar Sehgal and Anand Sagar, 2006. In vitro Isolation and Influence of Nutritional Conditions on the Mycelial Growth of the Entomopathogenic and Medicinal Fungus Cordyceps militaris. Plant Pathology Journal, 5: 315-321.
DOI: 10.3923/ppj.2006.315.321
URL: https://scialert.net/abstract/?doi=ppj.2006.315.321
DOI: 10.3923/ppj.2006.315.321
URL: https://scialert.net/abstract/?doi=ppj.2006.315.321
INTRODUCTION
The genus Cordyceps, a known group of entomopathogenic fungi, forms fruiting bodies in their insect hosts and 750 species in the genus have been recognized (Sung, 1996). Cordyceps militaris is parasitic on larvae and pupae of Lepidoptra. If the Ascospores alight on the integument of susceptible larvae and pupae, germ tubes may penetrate, possibly aided by their ability to hydrolyse chitin (Huber; 1958, McEwen, 1963). After infection, cylindrical hyphal bodies appear in the haemocoel of the pupa. The hyphal bodies increase by budding and the buds are distributed within the insect’s body. After death, mycelial growth follows and the body of the insect becomes transformed into a sclerotium, from which the perithecial stromata later develop (Webster, 1980).
Several Cordyceps species being regarded to have antitumour, antiaging, antidiabetic, immunomodulating, hypoglycemic, aphrodisiac and antimalarial activities (Chen et al., 1997; Isaka and Tanticharoen, 2001; Kiho et al., 1996; Kim et al., 2001; Kittakoop et al., 1999; Kuo et al., 1996; Mizuno, 1998; Zhao et al., 2002; Zhu et al., 1998). The cost of 1 kg of wild collected Cordyceps in the market varies from 30,000 to 60,000 Nepali Rupees in Nepal and about Rs 1 Lac in India. The high prize of this fungus is because of its medicinal benefits and difficulty associated with its harvesting due to its small size (Sharma, 2004).
Medicinal properties, high prize and difficulty associated with its harvesting due to small size of this fungus stress the need for its artificial cultivation and steps in this direction have been taken by in vitro isolation and to study influence of nutritional conditions on the mycelial growth of this fungus.
MATERIALS AND METHODS
Specimens collection and in vitro isolation: The material used in the present studies was Cordyceps militaris, a fascinating Ascomycetous fungus. Fruiting bodies of the fungus have been collected from Glen forests of District Shimla of Himachal Pradesh, India (Fig. 1a-c). Fruiting bodies have been preserved dry as well as wet (Ainsworth, 1971) and the specimens have been deposited in the Herbarium, Department of Biosciences, Himachal Pradesh University Shimla, India.
The cultures of C. militaris were raised from the stipe and stroma portion of healthy, sun-dried and fresh specimens. The specimens were first washed with distilled water and then the tissues from the stipe and stroma portion were cut with the help of sterilized blade. The bits of tissue (2-3 mm) were taken by sterilized forceps and dipped in 0.1% Mercuric Chloride solution for 5-10 sec and then washed with sterilized distilled water.
 | |
| Fig. 1: | (a) Fruiting bodies of Cordyceps militaris in its natural habitat. (b) Association of C. militaris with the host insect buried in soil. (c) Fruiting bodies of C. militaris emerging out from the head of the insect host. (d) Petri plate containing pure culture of C. militaris in Yeastal Potato Dextrose Agar Medium |
Now the tissue was placed on sterilized filter paper to remove the excess moisture. These bits of tissue were then transferred aseptically into the petri plates containing Potato Dextrose Agar (PDA) Medium with the help of sterilized forceps. Petri plates were then incubated at ambient temperature for at least 8-10 days and observed regularly for the appearance of culture. The actively growing mycelial colonies were subcultured to obtain pure cultures. Twelve solid and 5 liquid media have been tried during the present studies. All the media were prepared following (Tuite, 1969).
Inoculum preparation: Mycelial discs of 5 mm diameter were taken out with a presterilized borer under aseptic conditions, to be used as inoculum in solid media. In liquid media the inoculum preparation was different. Mycelial disc of 5 mm was transferred to 250 mL flask containing 50 mL of liquid medium and incubated at 25°C for 8-10 days. After 10 days, there appeared a ball of mycelium, which was homogenised in medium by a sterilized rod. The 5 mL of this homogenised mycelium was added to each of different liquid media as an inoculum used for further studies.
Recording of vegetative growth in solid and liquid media: Vegetative growth of mycelium in the solid media was measured by taking the diameter of colony in two directions at right angles. In liquid media, the mycelial mats were filtered through Whatman No. 1 filter paper discs of 7.5 cm diameter. The filter papers were previously oven dried at 80°C until constant weight and weighed after keeping in moisture free desiccator. After filtration, the mycelium along with filter paper was again oven dried as described above for 12 h and finally weighed to give dry weight of the mycelium. Three replicates of each medium were used and average values were taken for comparison of growth in different media. The medium with best mycelial growth was used in further studies i.e., for studying the effect of temperature, pH and light and darkness.
Effect of temperature: For the study of temperature requirement of the fungus in solid and liquid media, inoculated petri plates and flask were incubated at the following temperatures viz. 5, 10, 15, 20, 25, 30, 35 and 40°C in separate incubators on the best suited solid and liquid medium.
Effect of hydrogen ion concentration (pH): To record the effect of different pH on the growth of this fungus, the best solid medium was adjusted at different pH levels, viz. 4.0, 4.5, 5.0, 5.5, 6.0, 6.5, 7.0, 7.5, 8.0, 8.5, 9.0, 9.5, 10.0. The best liquid medium was adjusted at different pH levels viz., 4.0, 4.5, 5.0, 5.5, 6.0, 6.5, 7.0, 7.5, 8.0, 8.5. The pH was adjusted with the help of N/10 NaOH and N/10 HCl. The pH was checked with the help of digital type Phillips pH meter. The inoculated petri plates and flasks were incubated for 10 days at optimum temperature and after that the growth was measured.
Effect of light and darkness: Best selected solid and liquid medium with optimum pH was inoculated and was given light and dark treatment at optimum temperature. Growth was observed after 10 days of incubation.
Effect of different carbon and nitrogen sources: In order to find the best carbon source for the mycelial growth of the fungus, the amount of carbon present in best liquid medium was substituted with twelve carbon sources on the basis of their molecular formulae so as to provide equal amount of carbon, whereas, starch was added at the rate of 5 g L-1 of medium. Similarly the nitrogen present in best liquid medium was also replaced with nine nitrogen sources.
Effect of different mineral sources: Best liquid medium was used for studying the response of various mineral sources on the growth of the fungus. The mineral sources used were calcium chloride, ferrous sulphate, manganese sulphate and ammonium molybdate. Each mineral source was added in amount of 1, 2 and 5 mg L-1 of medium.
Effect of different growth regulators: Effect of various growth regulators Gibberelic acid, NAA (Naphthalene acetic acid), Kinetin and IBA (Indole butyric acid) all at 20,50,100 mg L-1 concentration were also evaluated for mycelial growth of fungus in best liquid medium.
Statistical analysis of the data: Data was statistically analysed for Standard Deviation (SD) and Critical Difference (CD). Critical Difference was calculated by Analysis of Variance (ANOVA) one-way classification model.
RESULTS
Mycelial characteristics: Mycelium was initially creamish white, becoming densely matted. As soon as the colony matures the colour of mycelia changes from creamish white to light brown. At approximately 14 days of incubation, the mycelium begins to form small nodules at the centre on the surface of the medium, appearing light brown while peripheral mycelia remain creamish white. As the medium is completely consumed, the mycelia become increasingly mud-like (Fig. 1d). When the mycelial growth was observed at periodic intervals, maximum growth of this fungus was attained at 10 days of incubation. After that there is no significant increase in mycelial growth. Keeping this information in mind in subsequent experiments, the final data related to mycelial growth was recorded only after 10 days of incubation.
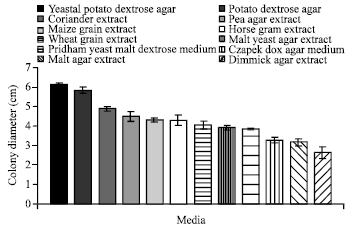 | |
| Fig. 2: | Effect of different solid media on the growth of C. militaris |
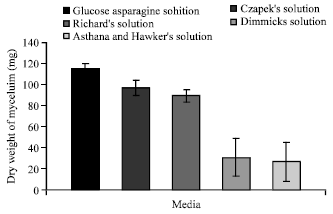 | |
| Fig. 3: | Effect of different liquid media on the growth of C. militaris |
The cultures were deposited in the Herbarium, Department of Biosciences, Himachal Pradesh University, Shimla India.
Growth of mycelium on different solid and liquid media: It is evident from (Fig. 2) that out of twelve solid media tested for mycelial growth, the maximum colony diameter (6.12 cm) was recorded on Yeastal Potato Dextrose Agar Medium followed by Potato Dextrose Agar Medium (5.83 cm). The minimum colony diameter (2.63 cm) was observed in Dimmick Agar Extract. Since the best growth of the fungus was recorded on Yeastal Potato Dextrose Agar Medium, it has been used as basal solid medium in subsequent studies and for the maintenance of cultures.
Out of five liquid media used to record the mycelial growth of the fungus. It is clear from the (Fig. 3) that the maximum dry weight (115.23 mg) of mycelium was recorded in glucose asparagine solution followed by that in Czapek’s solution (96.78 mg). Asthana and Hawker’s solution showed the least growth of mycelium (27 mg). Since Glucose Asparagine Solution showed the best growth of mycelium, hence it was used as the basal liquid medium for the subsequent studies.
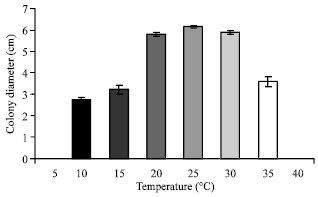 | |
| Fig. 4: | Effect of different temperature on the growth of C. militaris in basal solid medium |
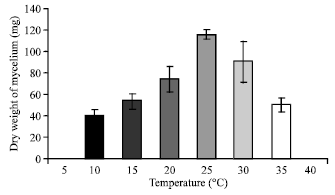 | |
| Fig. 5: | Effect of different temperature on the growth of C. militaris in basal liquid medium |
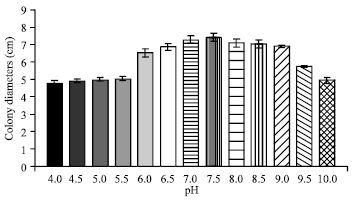 | |
| Fig. 6: | Effect of different pH on the growth of C. militaris in basal solid medium |
Effect of temperature: It can be seen from (Fig. 4 and 5) that the maximum colony diameter (6.15 cm) and maximum dry weight (115.23 mg) of mycelium was recorded at 25°C in basal solid and liquid medium respectively. The growth ceases completely at 5 and 40°C in both solid and liquid medium. Since the optimum temperature is 25°C, hence the subsequent studies were conducted at this temperature only.
Effect of hydrogen-ion concentration (pH): It is clear from (Fig. 6) that maximum growth of the mycelium (7.41 cm) was achieved at pH 7.5 followed by 7.0 (7.26 cm), 8.0 (7.08 cm) and 8.5 (7.03 cm) in descending order.
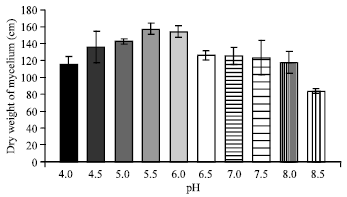 | |
| Fig. 7: | Effect of different pH on the growth of C. militaris in basal liquid medium |
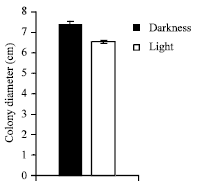 | |
| Fig. 8: | Effect of light and darkness on the growth of C. militaris in basal solid medium |
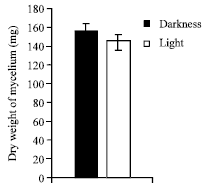 | |
| Fig. 9: | Effect of light and darkness on the growth of C. militaris in basal liquid medium |
Since, slightly alkaline pH of 7.5 was recorded to be optimum for the best growth of mycelium in basal solid medium.
It is evident from (Fig. 7) that the maximum dry weight (157.32 mg) of mycelium was recorded at pH 5.5 followed by pH 6.0 (154.15 mg) and pH 5.0 (142.85 mg) in descending order. Hence 5.5 is the optimum pH for the growth of C. militaris in basal liquid medium.
Effect of light and darkness: Basal solid and liquid medium were incubated at optimum temperature and optimum pH in light and darkness, growth was observed after 10 days of incubation.
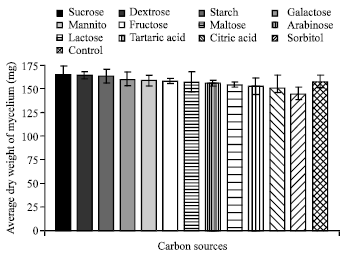 | |
| Fig. 10: | Effect of various carbon sources on the growth of C. militaris |
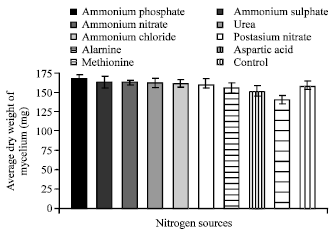 | |
| Fig. 11: | Effect of various nitrogen sources on the growth of C. militaris |
It is clear from (Fig. 8 and 9) that, in both basal solid and liquid medium the better mycelial growth was observed in darkness in comparison to light.
Effect of different carbon and nitrogen sources: Out of twelve carbon sources tested for mycelial growth of C. militaris. Maximum growth was observed in sucrose (165.02 mg) followed by dextrose (164.35 mg) whereas minimum growth was observed in sorbitol (145.02 mg) (Fig. 10). It is also clear from Fig. 11 that out of 9 nitrogen sources evaluated for maximum mycelial growth of fungus, ammonium phosphate (167.53 mg) was found to be best nitrogen sources followed by ammonium sulphate (163.22 mg). In methionine least growth (140.02 mg) was observed.
Effect of different mineral sources: It is evident from the data presented in Fig. 12 that mineral sources tested supported more mycelial growth of C. militaris as compared to control.
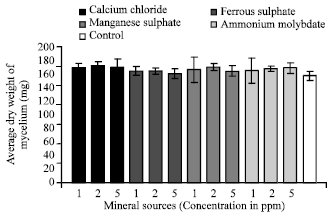 | |
| Fig. 12: | Effect of different mineral sources on the growth of C. militaris |
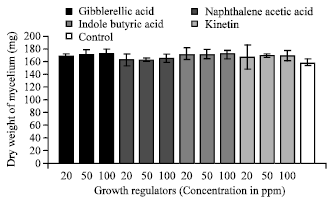 | |
| Fig. 13: | Effect of different growth regulators on the growth of C. militaris |
It was also revealed that 2 ppm concentration of calcium chloride and manganese sulphate was optimum requirement of the fungus, whereas it was 1 and 5 ppm for ferrous sulphate and ammonium molybdate respectively. An increase or decrease in the concentration beyond the optimum one resulted in decreased growth.
Effect of different growth regulators: Out of four growth regulators tested at three concentrations. All the concentrations of different growth regulators supported more growth of the fungus as compared to control. However gibberellic acid at 100 ppm concentration supported maximum mycelial growth than the lower concentration tested (Fig. 13).
DISCUSSION
In the present study in vitro isolation of Cordyceps militaris was done on Potato Dextrose Agar Medium initially. Similarly C. militaris, C. sinensis and C. unilateralis were grown on PDA successfully (Kim and Yun, 2005; Kim et al., 2003; Wongsa et al., 2005). Out of 12 solid and 5 liquid media tried for evaluation of maximum mycelial growth. Yeastal potato dextrose agar medium and glucose asparagine solution were found to be best solid and liquid medium, respectively. The exact reason for the maximum growth of mycelium in yeastal potato dextrose agar medium and glucose asparagine solution is not known, but it appears that the reason for such growth is probably the presence of Yeast and Asparagine, which as nitrogen source can enhance the growth of mycelium. Similarly maximum mycelial growth of C. militaris and C. sinensis was recorded in media containing nitrogen sources in comparison with media containing carbon sources (Kim et al., 2003; Kim and Yun, 2005). The maximum growth of C. militaris was recorded at 25°C. According to Kim et al. (2003) and Xu and Yun (2003) the most of the entomopathogenic fungi have similar temperature optima, ranging from 20-25°C. The maximum mycelial growth of C. militaris was observed at pH 7.5 and 5.5 in solid and liquid medium respectively. Sung et al. (1995) also observed that Cordyceps nutans and Phytocordyceps ninchukiospora grow best on pH 6.0 and Paecilomyces tenuipes on pH 7.0 and Cordyceps scarabaeicola on pH 9.0. The optimum pH for mycelial growth of entomopathogenic fungi is presumably dependent on the strain even though most of the fungi grow well at pH 5.0-pH 6.0 in their submerged cultures (Bae et al., 2000; Kim et al., 2002, 2003; Park et al., 2001). Mycelium was found to grow better under the condition of darkness in comparison to light.
The results of present study also revealed that sucrose and ammonium phosphate were the best carbon and nitrogen sources, respectively. Kim and Yun (2005) also reported the Sucrose and corn steep powder as carbon and nitrogen sources respectively for maximum mycelial growth of C. militaris and C. sinensis. Bae et al. (2000) observed a high level of mycelial growth of P. japonica in media containing maltose. Out of four mineral sources tested it was also revealed that 2 ppm concentration of calcium chloride and manganese sulphate was optimum requirement of the fungus, whereas it was 1 and 5 ppm for Ferrous sulphate and Ammonium molybdate respectively. Kim and Yun (2005) also reported CaCl2 as best mineral source for maximum mycelia growth of C. militaris and C. sinensis. Jonathan and Fasidi (2001) observed a high level of mycelial growth by Psathyerella atroumbonata in media containing calcium and magnesium. Chardonnet et al. (1999) found that external Ca2+ can play an indirect role in fungal growth by altering internal Ca2+, which controls the cytoplasmic Ca2+ gradiant and the activity of fungal enzymes involved in cell wall expansion. Flammulina velutipes showed best mycelial growth at 1 ppm FeSO4, 2 ppm ZnSO4 and MnSO4 and 10 ppm ammonium molybdate (Sharma et al., 2005). Among the four growth regulators Gibberellic acid supported maximum mycelial growth at 100 ppm concentration. Similarly, Sharma et al. (2005) also observed the maximum mycelial growth of Flammulina velutipes.
ACKNOWLEDGMENTS
The authors are thankful to the Prof. K.G. Mukerji, Department of Botany, University of Delhi, New Delhi, India for the authentication of specimens and its cultures. We also express thanks to Chairman Department of Biosciences, Himachal Pradesh University, Shimla India for providing necessary facilities for carrying out the present study.
REFERENCES
- Bae, J.T., J. Sinha, J.P. Park, C.H. Song and J.W. Yun, 2000. Optimization of submerged culture conditions for exo-biopolymer production by Paecilomyces japonica. J. Microbiol. Biotechnol., 10: 482-487.
Direct Link - Chardonnet, C.O., C.E. Sams and W.S. Conway, 1999. Calcium effect on the mycelial cell walls of Botrytis cinerea. Phytochemistry, 52: 967-973.
Direct Link - Chen, Y.J., M.S. Shiao, S.S. Lee and S.Y. Wang, 1997. Effect of Cordyceps sinensis on the proliferation and differentiation of human leukemic U937 cells. Life Sci., 60: 2349-2359.
CrossRefDirect Link - Isaka, M. and M. Tanticharoen, 2001. Structure of cordypyridones A-D, antimalarial n-hydroxy and n-methoxy-2-pyridones from the insect pathogenic fungus Cordyceps nipponica. J. Org. Chem., 66: 4803-4808.
CrossRefDirect Link - Jonathan, S.G. and I.O. Fasidi, 2001. Effect of carbon, nitrogen and mineral sources on growth of Psathyerella atroumbonata (Pegler), a Nigerian edible mushroom. Food Chem., 72: 479-483.
CrossRef - Kim, S.W., H.J. Hwang, C.P. Xu, Y.S. Na, S.K. Song and J.W. Yun, 2002. Influence of nutritional conditions on the mycelial growth and exopolysaccharide production in Paecilomyces sinclairii. Lett. Applied Microbiol., 34: 389-393.
Direct Link - Kim, S.W., H.J. Hwang, C.P. Xu, J.M. Sung, J.W. Choi and J.W. Yun, 2003. Optimization of submerged culture process for the production of mycelial biomass and exo-polysaccharides by Cordyceps militaris C738. J. Applied Microbiol., 94: 120-126.
Direct Link - Kim, H.O. and J.W. Yun, 2005. A comparative study on the production of exopolysaccharides between two entomopathogenic fungi Cordyceps militaris and Cordyceps sinensis in submerged mycelial cultures. J. Applied Microbiol., 99: 728-738.
Direct Link - Kittakoop, P., J. Punya, P. Kongsaeree, Y. Lertwerawat, A. Jintasirikul, M. Tanticharoen and Y. Thebtaranonth, 1999. Bioactive naphthoquinones from Cordyceps unilateralis. Phytochemistry, 52: 453-457.
CrossRefDirect Link - Park, J.P., S.W. Kim, H.J. Hwang and J.W. Yun, 2001. Optimization of submerged culture conditions for the mycelial growth and exo-biopolymer production by Cordyceps militaris. Lett. Applied Microbiol., 33: 76-81.
CrossRefDirect Link - Sharma, S., 2004. Trade of Cordyceps sinensis from high altitudes of the Indian Himalaya: Conservation and biotechnological priorities. Curr. Sci., 86: 1614-1619.
Direct Link - Wongsa, P., K. Tasanatai, P. Watts and N. Hywel-Jones, 2005. Isolation and in vitro cultivation of the insect pathogenic fungus Cordyceps unilateralis. Mycol. Res., 109: 936-940.
CrossRefDirect Link - Xu, C.P. and J.W. Yun, 2003. Optimization of submerged-culture conditions for mycelial growth and exo-bipolymer production by Auricularia polytricha (wood ears fungus) using the methods of uniform design and regression analysis. Biotechnol. Applied Biochem., 38: 193-199.
Direct Link - Zhao, C.S., W.T. Yin, J.Y. Wang, Y. Zhang, H. Yu, R. Cooper, C. Smidt and J.S. Zhu, 2002. Cordyceps Cs-4 improves glucose metabolism and increase insulin sensitivity in normal rats. J. Altern. Complement. Med., 8: 309-314.
PubMedDirect Link - Zhu, J.S., G.M Halpern and K. Jones, 1998. The scientific rediscovery of an ancient Chinese herbal medicine: Cordyceps sinensis part I. J. Altern. Complementary Med., 4: 289-303.
CrossRefDirect Link