
Robert T. Lartey
USDA/ARSINPARL, 1500 N Central Ave, Sidney, MT 59270, USA
Plant Pathology Journal
Year: 2006 | Volume: 5 | Issue: 2 | Page No.: 125-142







ABSTRACT
The cropland soil, which is comprised of spermosphere, rhizosphere and bulk soil, is populated by a wide array of microbial inhabitants. These microbial inhabitants include the flora, represented by bacteria, actinomyces, archaea, fungi and algae and the fauna, such as protozoa, nematodes, Acarids and Collembola. These organisms consist of saprophytes, pathogens of plants and antagonists of other soil microbes, including some plant pathogens. The success of crops may be influenced directly by stimulation of their growth and indirectly by variety of interactions among these microbial inhabitants. These interactions may result in negative plant growth by causing a variety of diseases, positive plant growth through mutual benefits or no stimulation on plant growth through neutral effects. In addition, interactions also occur among these cropland microbial populations which also influences the health of crops. The most important of these, with regard to crop health is the antagonistic interactions between beneficial microbial inhabitants and pathogens which could impact biological control efficacy. Therefore, successful development of a biological control system against soil inhabiting plant pathogens requires good knowledge of the composition of the microbial inhabitants and an understanding of the interactive functions of microflora and fauna in the soil.
PDF Abstract XML References
How to cite this article
Robert T. Lartey, 2006. Dynamics of Soil Flora and Fauna in Biological Control of Soil Inhabiting Plant Pathogens. Plant Pathology Journal, 5: 125-142.
DOI: 10.3923/ppj.2006.125.142
URL: https://scialert.net/abstract/?doi=ppj.2006.125.142
DOI: 10.3923/ppj.2006.125.142
URL: https://scialert.net/abstract/?doi=ppj.2006.125.142
INTRODUCTION
The cropland soil is inhabited by a wide array of microbial populations which are composed of flora and fauna. Soil microbial populations have been thought to be comprised of five functional groups. They include autotrophic primary producers such as photosynthetic bacteria and ammonia-oxidizing archaea. There are also decomposers (saprophytes) which include micro- and mesoflora and fauna. The mutualists are composed of microbes such as nitrogen-fixing bacteria and mycorrhizal fungi that form associations with plants. The soil pathogens, pests, their antagonists and propagules of these micro- and mesoorganisms consist of bacteria and fungi, nematodes, Acarid (mites) and Collembola (springtails). Another group of microorganisms needing mentioned are the chemolithotrophs which have been observed in wetlands. These bacteria derive their energy from small and inorganic molecules such as iron, sulphur, hydrogen sulphide, hydrogen and methane compounds. However, for the purpose of this review, which focuses on cropland microbes, only saprophytes, soilborne pathogens, surviving propagules of foliar pathogens and antagonists of other microbial inhabitants of the soil will be discussed. The dynamics of these microbial populations are influenced by several factors such as location and prevailing environmental conditions such as moisture and temperature. Various forms of interaction occur among these organisms which in turn have significant impacts on the success of crops. With regard to plant health, the success of crops is influenced by direct relationships such as disease induction and growth stimulation and indirectly through a variety of interactions among the microbial populations. Under optimal conditions, the pathogenic populations cause various diseases which lead to reduced plant health and yield loss. Antagonists of pathogenic populations reduce inoculum potential and suppress diseases, thereby enhancing plant growth and yield. A key to design of a successful biological control strategy is good knowledge of the dynamics of the cropland microbial population structure and interactions among them. Soilborne plant pathogens, while not exclusive, have been primary targets for application of biological agents to control plant diseases, thus giving less attention to surviving propagules of the foliar pathogens, some of which survive for years in the cropland soil. While most of the attention has been focused on application of the floral agents, several studies have shown that some fauna antagonists are also good potential agents and in some cases, their combinations with floral agents have provided enhanced control of some soilborne pathogens. This is more in line with natural soil conditions where the flora and fauna co-exist. Past reviews have treated biological control by flora and fauna independently. This review attempts to bring to light the importance of both soilborne pathogens and surviving propagules of foliar pathogens as targets for biological control. It also looks at the microflora, microfauna and mesofauna together as potential tools for control of target pathogens. Some overlooked biocontrol targets among the microbial inhabitants of cropland soil and their pathogenic (disease causing) relations to crops are discussed. Furthermore, some unexplored agents such as the algae and their potential as antagonists and for biological control of plant pathogens are also discussed.
THE SOIL ENVIRONMENT
The soil is a living system that represents a finite resource vital to life on earth. It forms the thin unconsolidated mineral and organic matter on earth surface and develops slowly from various parent materials and is modified by time, climate, macro- and micro-organisms, vegetation and topography. It is a complex mixture of minerals, organic compounds and living organisms that interact continuously in response to natural and imposed biological, chemical and physical changes (Allan et al., 1995). Water, which serves as solute for various nutrients for the plant, is a vital component of the soil environment. Besides anchoring the roots, the soil is a dynamic system that serves as natural medium of growth of land plants and is inhabited by a wide range of microorganisms (SSSA, 1984). The microbial inhabitants consist of autotrophs, saprophytes, mutualists, parasites of plants and antagonists of other soil microbes. Growth of plants and the soil microorganisms may be influenced by both direct stimulation and indirectly through a variety of interactions. These interactions may result in negative growth through antagonism, positive growth through mutual benefits or no growth stimulation through neutral effects and can influence biological control efficacy. Successful biological control of soil inhabiting plant pathogens requires complete understanding of the interactive functions of microflora and fauna in the specific soil ecology (Lartey and Conway, 2004).
Cropland soil: There are various ways of classifying the soil (Bruehl, 1987). However, for the purpose of this discussion, which focuses on croplands, the soil can be grouped into the spermosphere, the rhizosphere or the bulk soil. Most cropland soils are dynamic in that, any one of the three soil types has a potential to be converted into one of the other two. In cropland systems with annual crops, the roles could vary from one growing season to another. Thus the soil that may be part of the spermosphere in one growing season may be transformed to a rhizosphere or bulk soil in another season and vice versa. Consequently, surviving soil biota may often find themselves subjected to a changing, newly transformed soil environment.
Spermosphere: The spermosphere has been described as the short-lived, rapidly changing and microbiologically dynamic zone of soil surrounding a germinating seed (Nelson, 2004). Slykhuis (1947) first used the word spermatosphere to describe the distinct environment of the immediate surrounding area of germinating seeds. According to Nelson (2004), the concept of spermosphere was developed by Verona (1958) to describe the zone of elevated microbial activity around germinating seed. The development of the spermosphere is linked to germination of the seed (Nelson, 2004) which commences following a period of dormancy under external or environmental conditions such as water, oxygen and temperature (Raven et al., 1981). Following sowing, water moves rapidly into the seed and hydrates it in what is termed imbibition. Stored enzymes in the seed are activated and new ones are synthesized to digest stored food in the seed. As the seed imbibes water, it swells and builds considerable pressure in the seed. Respiration is initially anaerobic during the early stages of germination. Eventually, the seed coat ruptures and switches to aerobic respiration. The rupture of the seed coat is accompanied by rapid leakage of cellular and vacuolar constituents which are referred to as seed exudates (Nelson, 2004). The spermosphere differs from the rhizosphere in that the former is influenced by seed threshing injuries, maturation, age and other factors (Bruehl, 1987). Although it is widely recognized, spermosphere related research has not kept up with the research on the rhizosphere.
Rhizosphere: It was first described a century ago by Hiltner (1904) as the narrow zone around leguminous roots that stimulates intense bacterial activities. The microorganisms which are influenced by the root exudates have been expanded over time to include bacteria, actinomycetes, fungi, protozoa, nematodes and arthropods. Some terminologies have evolved to describe or elaborate other aspects of the rhizosphere. Clark (1949) used the word rhizoplane to describe the actual root surface in distinction from the soil around the root. Box and Hammond (1990) defined the rhizosphere as the whole soil mass occupied by the root. Balandreau and Knowles (1978) suggested endorhizosphere to describe the root epidermis and the cortex. Lynch (1982) proposed ectorhizosphere as the colonized zone outside the root including the surrounding soil. Curl (1988) stated that the rhizoplane and the rhizosphere are sometimes referred to as the soil-root interface. The establishment of rhizosphere commences with the release of organic compounds (exudates) from the plant roots into the rhizoenvironment. These compounds include root secretions, plant mucilage, mucigel and lysates (Rovira et al., 1979). Thus the most important factors that contribute toward the rhizosphere dynamics are the exudates. These attract variety of organisms which trigger the interaction among these organisms and the dynamics thereof.
In this discussion, the scope of the rhizosphere will be limited to the original description by Hiltner. However, the discussion of rhizosphere effects will not be limited to activities of bacteria around leguminous roots, but also cover the array of micro and mesoorganisms and their interactions with one another with emphasis on biological control. The rhizosphere as the interface between the plant root and its soil environment is probably the most important micro ecological niche that affects diverse aspects of plant life. It is complex and dynamic as a result of interaction among the host plant root and various microorganisms and invertebrate inhabitants which directly or indirectly affect the health of the plant.
Bulk soil: Soil outside the influence of germinating seeds (the spermosphere) or roots (the rhizosphere) forms the bulk soil component of cropland soils. Although it is considered to be the least dynamic with regard to microbial activities, it is the largest of the three categories of cropland soils. Because of its volume, the bulk soil harbors a highly significant population of micro and mesoorganisms including surviving propagules of soil inhabiting (foliar and soilborne) plant pathogens. Several foliar fungal plant pathogens survive as saprophytes in the soil and thus, for that duration, could be considered soil inhabitants. For example, Cercospora beticola is considered a foliar pathogen even though it can survive in the soil for several years (McKay and Pool, 1918). Because it lacks direct contact to crops, very modest research efforts in terms of microbial activities have been directed at the bulk soil per se. Indeed most research efforts that have involved the bulk soil were actually directed at comparisons with either the spermosphere or the rhizosphere (Lupwayi et al., 1998; Gomes et al., 2003; Nielsen and Sørensen, 2003).
SOIL INHABITANTS
A good perceptive view of microbial systems in the soil is an important prerequisite to understanding the impact of soil dynamics on biological control. The resident soil micro- and mesoorganisms could be classified in the flora and the fauna. Soil flora includes bacteria, fungi and algae and tends to be more static. The fauna on the other hand tend to move about once the food source is exhausted. Fauna includes protozoa, nematodes, Collembola and acarids.
The Microflora: The soil microflora have received the most attention with regard to soil microbial interactions and biological control. The cropland flora comprised of bacteria, actinomyces and fungi have been reviewed extensively regard to the spermosphere and the rhizosphere activities and plant health (Curl and Truelove, 1986; Buyer et al., 1999; Whipps, 2001; Nelson, 2004). Since rhizodeposition products stimulate bacteria and fungi, their population and activities tend to be higher in the rhizosphere than in the bulk soil. With regard to the spermosphere, functionality of the community appears to change more than structure with substrate utilization when compared to activities in the control bulk soil (Buyer et al., 1999). There are also the algae in the soil; these have yet to receive significant attention with regard to biological control.
Bacteria: The bacteria are the predominant microorganisms in the soil irrespective of the method of determination (Curl and Truelove, 1986). Ecological differentiation of soil bacteria was first proposed by Winogradsky (1925). He grouped them into autochthonous (indigenous) and zymogenous (fermentation) groups. However, current classification is based on systems in the Bergey’s Manual of Determinative Bacteriology. Estimates of bacterial numbers may vary according to the method of determination (Alexander, 1961). Distribution appears to be influenced by several factors. Alexander (1961) cited Starc (1942) as having observed differences in various horizons of soil profiles in which aerobic bacterial were the most predominant, followed by actinomycetes and the anaerobic bacteria. Rouatt and Katznelson (1961) observed higher bacterial populations in the rhizosphere when compared to the bulk soil of several field crops. According to Bakken (1997) and Alexander (1961) attempted to summarize a great number of classical taxonomical studies of prevailing soil and presented the following range: Arthrobacter 5-60%; Bacillus 7-67%; Pseudomonas 3-15%; Agrobacterium up to 20% Alcaligens 2-12% and Flavobacterium 2-10%.
Actinomycetes: Although actinomycetes actually are classified as bacteria, they have for historical reasons been considered separately. They were originally thought to be fungi because of their filamentous growth characteristics. The actinomycetes are considered to be the second most abundant in the soil after the other bacteria. They are Gram-positive prokaryotes and tend to grow slowly with branching elements. According to Curl and Truelove (1986), most soil actinomycetes fall within the genera Streptomyces, Micromonospora or Nocardia of the Order Actinomycetaceae. The Streptomyces are slow growing, aerobic and produce extensively branched mycelia, with a diameter between 0.5 to 2.0 μm. They are Gram-positive but not acid fast. The aerial mycelia bear long chains of spores which are formed by fragmentation of the filaments. The genus Nocardia also are aerobic but some are acid fast at some stages of growth. The spores are non motile and the chain formation varies among the species. The genus Micromonospora produces septate mycelium with an average of about 0.5 μm diameter. They are aerobic but not acid fast and produce non motile single spores (Lacey, 1973; Locci, 1989). Actinomycetes species within all these groups have been isolated and identified in various countries with the Streptomyces being the dominant species in the soil.
Archaea: Until recently the prokaryotes archaea were referred to as archaebacteria. Since they were thought to be bacteria, they were placed in the same Kingdom of Monera. Based on molecular information, Woese et al. (1990) proposed reclassification of the archaea, which have subsequently been placed in their own domain. The archaea are microscopic, single-celled organisms generally with a single circular DNA and lacking membrane-bound organelles. Various species have been isolated from extremely harsh environments (Madigan and Marrs, 1997). In fact, archaeans habitats include some of the most extreme environments on the planet. They have been found at temperatures well over 100°C, in hot springs (such as the Yellowstone National Park, Wyoming, USA,) and in both extremely alkaline and acid waters. They have been found inside the digestive tracts of cows, in termites and in marine life where they produce methane. They live in the anoxic muds of marshes and at the bottom of the ocean and even in petroleum deposits deep underground. More recently, archaea have been isolated from cropland soil (Bintrim et al., 1997).
Fungi: Fungi are considered to be the next most abundant microflora after the bacteria and actinomyces in the soil. Curl and Truelove (1986) stated that the fungi are actually more difficult to assess than bacteria because of their filamentous nature and that profusely sporulating species are favored by the soil dilution plate isolation procedure. Therefore they expressed a need for caution when fungal populations are compared with those of bacteria. According to Alexander (1961), fungi account for the largest part of total microbial protoplasm in well-aerated cultivated soil. The soil ecosystem harbors almost all major taxonomic groups of fungi including bacterium-engulfing slime molds, root decay, mycorrhizal and root-pathogenic basidiomycetes (Thorn, 1997). He cited several references that indicate variations among the fungal communities with respect to major gradients in latitude and elevation, vegetation, soil antibiotics and season. According to Christensen (1981) for example, Penicillium is the predominant species in temperate to higher latitudes, Aspergillus in grasslands and deserts and Fusarium in grassland soils. Other reports have shown variations of fungal community structures with agricultural practices such as tillage (Lupwayi et al., 1998).
Algae: Algae are not as abundant as bacteria, actinomycetes or fungi, but are common in habitats with adequate moisture and light (Alexander, 1961). With regard to cropland soil and interaction with crops and microbial inhabitants, the algae have received very little attention. In one of the few interesting studies, Rouatt et al. (1960) found lower numbers in the rhizosphere than in the bulk soil. The soil algae belong generally to four groups, namely the blue-green (Cyanophyceae), green (Chlorophyceae), yellow-green (Xanthophyceae) and diatoms (Bacillariaceae). Species within these four groups have been found to be dominant in North America and in soils of arid regions of North Africa, the Middle East and Asia (Evans and Johansen, 1999). They are characteristically photoautotrophic and would therefore be anticipated to flourish under high light-energy and in the presence of CO2. However, some species within the blue-green algae, green algae and diatoms are also capable of growing heterotrophically (Curl and Truelove, 1986). According to the authors (Curl and Truelove, 1986), highest populations occur in the top 10 cm of the soil, although isolates have been obtained from between 50 and 100 cm deep.
The micro and mesofauna: The soil fauna consists of the microfauna, the mesofauna and the macrofauna. Unlike the microflora, the microfauna and the mesofauna have received only modest attention with regard to control of soil-inhabiting plant pathogens. Arbitrary grouping of the microfauna primarily refers to animals with body widths of < 100 μm (Swift et al., 1979), thus encompassing the large populations of free-living protozoa and nematodes. The mesofauna (largely microarthropods) generally measure 1-2 mm. These invertebrates, because of their small body size and feeding habits, are most likely to respond to root and seed exudates and biotic phenomena in the spermosphere and the rhizosphere. Mesoscopic members of the Arthropoda commonly referred to as microarthropods, like other small soil animals, vary in population with season, soil chemical and physical factors, vegetative cover, tillage practices and food availability (Loring et al., 1981; Petersen, 1982; Andrén and Lagerlof, 1983; Mallow et al., 1985). The macrofauna, exemplified by large arthropods and the annelids, are important in organic matter fragmentation and decomposition but are largely inhabitants of surface litter and do not fit prominently into the interactive scheme within microniches of the spermosphere and the rhizosphere. This discussion will be limited to activities of the micro and mesofauna and include protozoa, nematodes, Collembola and Acarina.
Protozoa: Most protozoa are represented by flagellates and small amoebae, their distribution being determined by water films on soil particles and pore size (Bamforth, 1988). Both uncultivated and cultivated soils harbor relatively abundant and heterogeneous populations of protozoa. A conservative estimate for a fertile agricultural soil might be 2x103 to 17x105 g-1 dry wt of soil, the dominant group being the naked amoebae (Heal, 1971; Elliott and Coleman, 1977). Peak populations of amoebae may be expected to follow peaks in numbers and biomass of bacteria, especially after rainfall (Clarholm, 1981). High populations in plant rhizospheres in terms of R/S (rhizosphere to nonrhizosphere) ratios may range from 2:1 for total protozoa to 20:1 for some species. Generally, bacteria and yeasts are preferred food sources for protozoa but they can also feed on some fungi. Thus, certain species of mycophagous amoebae in agroecosystems can destroy a sufficient quantity of pathogenic fungal propagules to induce significant biological control of specific root diseases (Old and Chakraborty, 1986).
Nematodes: Estimates of population density of free-living nematodes vary widely with season and kinds of plant cover, ranging from 1x104 to 18x105 m-2 for agroecosystems (Stinner and Crossley Jr., 1982). Summary estimates in terms of biomass (Hendrix et al., 1986) showed non plant parasitic nematodes to be more abundant in conventional tillage systems than in no-tillage agroecosystems; highest values were for bacterivores rather than fungivores or herbivores. Total nematode biomass is generally high in temperate grasslands (Petersen, 1982). Studies in Poland (Golebiowska and Ryszkowski, 1977), Sweden (Sohlenius and Bostrom, 1984) and the USA (Parmelee and Alston, 1986) showed that numerical dominance of bacteria in agroecosystems is accompanied generally by higher populations of bacteriovorus nematodes. Along with bacteria-feeding species, populations of mycophagous species of Aphelenchoides, Aphelenchus, Ditylenchus and Neotylenchus are relatively abundant in various cropping systems (Nicholas, 1984). Species of nematodes that feed upon fungi are found in several genera in cropland soils. Some food preference has been noted; for example, Ditylenchus destructor fed and reproduced on 64 of 115 fungal species tested in laboratory cultures (Faulkner and Darling, 1961).
Collembola (Springtails): The Collembola are the most abundant and important of the orders of primitive insects in the Apterygota. They are well represented in agricultural soils to 15 cm depth and may migrate to much greater depths, depending upon soil type, moisture and cropping practices. Collembola and other invertebrates that feed on the microflora are not uniformly distributed in the soil but tend to congregate around living, dead or dying roots and organic matter where food sources are most abundant. The rhizosphere factor is important in relation to microarthropod potential to challenge plant pathogens at the infection site. Laboratory tests showed that Collembola are attracted to cotton roots, particularly in slowly drying soil (Wiggins et al., 1979). In conventional agroecosystems, the biomass of soil-dwelling mites was estimated to be 118 mg and Collembola 17 mg dry wt. m-2 (Hendrix et al., 1986); these figures were higher in no-tillage systems. Extensive field assessments in Alabama (Wiggins et al., 1979) provided evidence that two collembolan species, Proisotoma minuta and Onychiurus encarpatus, were predominant in cotton rhizosphere; furthermore, they were significantly more numerous in the rhizosphere than in root-free soil regardless of long-term fertilization regimes. A direct relationship was also demonstrated between increasing cotton-root density and collembolan populations. Collembolan activities in the rhizosphere of other crops have been studied by direct observation techniques. Using minirhizotron in Michigan, Snider et al. (1990), found that Folsomia candida was much more numerous in the rhizosphere of soybean and sugar beet than in the rhizosphere of corn agroecosystems. Thus, it is obvious that microflora populations and rhizodeposition products promote the activities of small soil fauna with the potential to alter the behavior of plant pathogens.
Acarina (Mites): The soil mites and Collembola are often the most abundant forms of microarthropods that remain permanently in the soil (Odum, 1959). According to Curl and Truelove (1986) the mites are more common in the soil than Collembola but are not the most prominent in most soil. Of the seven orders of Acarina, the Crystostigmata, Mesostigmata, Prostigmata and Astigmata are the most common in the soil (Bardgett and Griffiths, 1997). Wallwork in 1976, cited by Curl and Truelove (1986), stated that the predominant soil dwelling cryptostigmatid mites are composed of wood tissue feeding macrophytophages, leaf tissue feeding phyllophages, fungi feeding mycophages, bacteria feeding bacteriophages or algae feeding phycophages. In an example of a population study, it was estimated that the number of Acarina were relatively higher than the Collembola in European grassland soil (Kevan, 1965).
INTERACTION: BASIS FOR BIOLOGICAL CONTROL
The soil inhabitants include the seed and root infecting pathogens such as the damping off causing Rhizoctonia solani and surviving propagules of foliar pathogens such as C. beticola and Streptomyces scabie which may survive sometimes for several years in the soil. The basis for biocontrol of soil inhabiting plant pathogens is the interaction among the antagonists and pathogenic inhabitants. Interactions among organisms as basis for developing a biological control system was recently reviewed by Lartey and Conway (2004). By definition, the term antagonism falls within the realm of growth inhibition relationship and is the foundation for biological control and features competition, antibiosis, mycoparasitism, ammensalism and predation. The first three types of relations, namely competition, antibiosis and mycoparasitism occur commonly between flora and growth of one or both interacting organisms are adversely affected. Both interacting organisms are adversely affected by competition, thus resulting in reduced growth of both organisms. This may be due to competition for limited resources. Under antibiosis, organic products of one microorganism are detrimental to growth or metabolic activities of another organism. Under parasitism one organism establishes a long term relationship with a host from which it derives nutrients and frequently, death does not occur immediately. In some cases, the parasitism does not result in the death of the host. These relations are common when the biological agent is a member of the microflora. On the other hand, predation results in the death of the host within a short period of time. Under amensalism, one interacting partner is adversely affected while the other is not affected.
SOIL MICROFLORA AS AGENTS FOR BIOLOGICAL CONTROL
Biological control by the soil flora: The potential of the soil flora as biological agents has been recognized since the early days of biological control (Garrett, 1965) and continues to the present. An interesting trait of the flora is the continuously altering of their roles as pathogens or biological agents and extensive interactions within and outside of the floral group. It is thus not unusual to control a bacterium with another one and even non-pathogenic isolates of a pathogenic species are sometimes applied to control a pathogenic isolate. Control of soil inhabiting plant pathogens with the soil bacteria has often been considered in association with either the spermosphere or the rhizosphere and less for the bulk soil, where in the rare cases, studies have focused on comparison to the spermosphere or the rhizosphere bacteria. There have been extensive reports of control of bacterial pathogens with other bacteria. For example, Pseudomonas fluorescens was applied to control Erwinia carotovora and soft rot of potato (Cronin et al., 1999). Bacterial agents have also been applied to control other plant pathogens such as fungi and nematodes. The bacterium Pseudomonas putida was applied to control Fusarium wilt on radish (De Boer et al., 2003) and Paenibacillus sp. 300 to control Fusarium wilt of Cucumber. P. putida was applied weekly to soil during 1997-1998 irrigation seasons and suppressed populations of Phytophthora parasiticum in two citrus orchards over three growing seasons (Steddom et al., 2002). Shaukat and Siddiqui (2003) observed significant inhibition of the nematode Meloidogyne javanica by antagonistic Rhizobia. Sulfate reducing bacteria Desulfo-vibrio sp. were also been used to control the rice nematode Hirschmanniella oryzue in Senegal (Jacq and Fortuner, 1979).
Several species of Actinomycetes have been identified and documented to control a variety of soilborne plant pathogens. In a recent study in Turkey, 50 different actinomycetes were assessed and 34% showed antibacterial activity against at least one of the tested six phytopathogenic bacteria including Agrobacteria tumefaciens, Erwinia amylovora and Pseudomonas viridiflova (Oskay et al., 2004). In a two year greenhouse experiment, Streptomyces griseovirides and the fungus Gliocladium catenulatum were most effective in reducing plant mortality from incidence of Pythium aphanidermatum in cucumbers (Punja and Yip, 2003). Increases in plant height resulted from the effects of reducing disease by the two agents. In another experiment, Singh et al. (1999) observed suppression of Fusarium wilt of cucumber with a chytinolitic Streptomyces spp. in potting soil in growth chamber experiment.
Currently, I am not aware of any research effort to control bacterial pathogens directly with fungal antagonists. However cercosporin, a phytotoxin that is produced by the sugar beet pathogen C. beticola and other Cercospora species, has been shown to inhibit the growth of Escherichia coli (Fig. 1) (Lartey and Caesar-TonThat, unpublished).
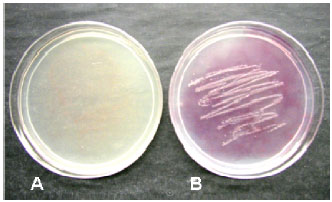 | |
| Fig. 1: | Inhibition of Escherichia coli by cercosporin from Cercospora beticola. 100 μL of cercosporin solution (25 μg/ml) was applied to potato dextrose agar (PDA). The plates were then streak inoculated with E. coli and half were incubated in darkness and the other half under light at 37°C overnight. Growth of E. coli was inhibited under light (A) while growth inhibition was not observed in the dark (B). |
Thus there is considerable potential for fungi to antagonize and suppress bacterial pathogens. While a number of fungi have been evaluated as biological agents to control plant pathogens, the Trichoderma spp. are certainly the clear leaders among the fungal antagonists which, according to Whipps and Lumsden (2001), may be a reflection of their ease of culture and the size of their potential hosts. Trichoderma harzianum, like other species within the genus, has been used to control various pathogens on a number of crops. Some examples are presented in the following when application of T. harzianum suppressed R. solani and damping-off on radish, damping-off of snap beans, collar-rot of coffee, seed rot and stem and root lesions on beans and also resulted 86% reduction of R. solani inoculum potential in naturally infested soil (Henis et al., 1978; Chet and Baker, 1980; Marshall, 1982; Venkatasubbaiah et al., 1984; D'Ercole et al., 1985; Strashnov et al., 1985). Beside T. harzianum, other Trichoderma spp have been shown to control various plant fungal pathogens. Tu and Vaartaja (1981) and Beagle-Ristaino and Papavizas (1985) showed reduced incidences respectively of root rot of white beans and stem canker and black scurf of potato, all caused by R. solani, by application of T. (Gliocladium) virens. The Trichoderma species have also been applied to control other fungal pathogens such as Sclerotium rolfsii with T. harzianum on beans (Elad et al., 1980). T. harzanium also suppressed Fusarium spp. including Fusarium oxysporum f. sp. vasinfectum, on cotton, F. oxysporum f. sp. melonis on melon, F. oxysporum f. sp. radicis on tomato and F. culmorum on wheat respectively (Sivan and Chet, 1986). Other Trichoderma species such as T. virens was also observed to reduce damping of Pythium ultimum (Howell, 1982). Other fungi, including Laetisaria arvalis have been demonstrated to control other fungal pathogens. For example, Lewis and Papavizas (1980) applied mycelial and sclerotia preparations of L. arvalis to soil and decreased severity of cucumber fruit rot caused by R. solani. L. arvalis has been observed to control R. solani on other corps (Odvody et al., 1980; Allen et al., 1982; Martin et al., 1984; Larsen et al., 1985). Fungi within other genera such as non-pathogenic Pythium spp, Fusarium spp and binucleate R. solani are increasingly being considered for biocontrol of phytopathogens. Honeycutt and Benson (2001), for example, were able to obtain control of damping-off by R. solani on impatiens with some formulations of binucleate Rhizoctonia spp.
No information about either the application of archaea or algae to control soil inhabiting plant pathogens has been found in my literature search. However, some algae, in particular the Cyanobacteria are known to produce a variety of toxins (Katircioglu et al., 2004). Some of these toxins may play a defensive role against competitors including soil inhabiting plant pathogens and therefore play significant roles in the survival of the algae in the soil. This opens up a possible selective application of some algae for biocontrol of various soil inhabiting plant pathogens. This suggestion warrants serious investigations in the future.
Soil flora modes of antagonism: The basis for biocontrol by the flora is very diverse and encompasses various forms of growth inhibiting relationships (antagonism) (Lartey and Conway, 2004). The relationship is characterized by growth or survival of the biological agent to the detriment of the targeted pathogen. Within the realm of these growth inhibition relationships are amensalism, competition, antibiosis and parasitism. Competition, antibiosis and parasitism are very typical of the flora and occur quite frequently.
Amensalism: With regard to microbial interaction, there have been few research reports on documented cases of amensalism. This may be due to difficulty in resolving the boundary between amensalism, parasitism and competition (Bull and Slater, 1982). In one study of the interaction between Zymomonas mobilis and Saccharomyces cerevisiae in a mixed culture experiment, growth of S. cerevisiae was inhibited with acetaldehyde that was produced by Z. mobilis under aerobic conditions (Tanaka et al., 1999). The acetaldehyde had no effect on Z. mobilis.
Competition: Based on a definition by Clements and Shelford in 1936, Clark (1965) described competition as the active demand in excess of the immediate supply of material or condition on the part of two or more organisms. A classical example of competition was reported by Gause (1934), who observed that growth and population levels remained constant when two closely related ciliated Protozoa, Paramecium caudatum and Paramecium aurelia, were cultured individually with an adequate supply of bacteria. When combined however, only P. aurelia survived after 16 days even though both organisms neither attacked the other nor secreted toxic substances. P. aurelia grew more rapidly and thus out-competed P. caudatum for the available food. More recently, competition was observed among species of yeasts that colonize ripening and deteriorating fruits during studies on interaction among different species (Abranche et al., 2001). The competition was attributed to such factors as limiting singular or multiple nutrients.
Antibiosis: Antibiotics are organic products of microorganisms, which are detrimental in low concentrations to growth and metabolism of other microorganisms (Gottlieb and Shaw, 1970). A review of the subject by Baker (1980) reinforced the acceptance of inhibition by antibiotics in the soil as a form of antagonism. Chang and Kommedahl (1968) and Kommedahl and Mew (1975) inoculated corn grains with Bacillus sublitis or Chaetomium globosum and observed protection against seedling blight caused by Fusarium roseum; the pathogen apparently was inhibited by antibiotics produced by the biocontrol agents during the period of maximum susceptibility (Kommedahl and Mew, 1975). Crown gall caused by A. tumefaciens has been controlled by Agrobacterium radiobacter strain 84, which induced antibiosis after inoculation of seeds or nursery plant roots before planting (Kerr, 1972; New and Kerr, 1972; Moore and Warren, 1979). Species of Streptomyces, Pseudomonas, Penicillium, Trichoderma and Aspergillus also are known to produce antibiotics (Curl et al., 1985). The antibiotic phenazines produced by Pseudomonas aureofaciens have been described as key to its competitive antagonism against some fungal pathogens such as Geumanomuces graminis (Pierson and Pierson, 1996). According to Aluko and Hering (1970), T. virens produces two antifungal antibiotics, gliotoxin and viridin and also a non-volatile antibiotic which is active against Heterobasidion annosum and other fungi. Tu (1980) observed inhibition of sclerotia formation by Sclerotinia sclerotiorum; moreover, those sclerotia which were already formed were parasitized by T. virens.
Parasitism: Richards (1987) described a parasite as an organism that lives either in or on its host, the latter serving as both the habitat and the substrate. Under this long-term relationship, the parasite derives nutrients from the host and the host’s death may not occur immediately or may not occur at all. The host is the source of energy and biosynthesis and usually is exploited to such an extent that its metabolic activities are adversely affected with a resultant suppression of growth and development. Parasitism of one microorganism upon another one is common but highly specific (Curl et al., 1985). Various parasites have been isolated and identified. The bacterium Bdellovibrio was observed parasitizing pathogenic Pseudomonads (Stolp, 1973). T. harzianum was also observed parasitizing R. solani (Camporota, 1985). Using infrared photographic techniques, parasitism of R. solani by T. harzianum was also demonstrated (Stolp, 1973). Mycoparasitism, parasitism of fungi by other fungi, also has been observed by several researchers (Boosalis, 1964; Boosalis and Mankau, 1965; Lumsden, 1981). Parasitism of nematodes by fungi has been reported in Britain (Gottlieb and Shaw, 1970; Willcox and Tribe, 1974; Tribe, 1979). Morgan-Jones et al. (1981; 1983) also demonstrated parasitism of Meloidogyne arenaria by Verticillium chlamydosporium. Parasitism as a mechanism of biocontrol of R. solani by L. arvalis has also been observed in laboratory cultures (Lartey et al., 1994). The antagonist penetrated hyphal strands of R. solani and developed in the host to create a stable relationship. Phillips (1986) observed parasitism of S. sclerotiorum, Sclerotinia minor, Botrytis cinerea, Sclerotium rolfsii and Macrophomina phaseolina by T. virens.
SOIL FAUNA AS AGENTS FOR BIOLOGICAL CONTROL
In his discussions, Garrett (1965) did not include small soil animals as potential pathogen deterrents in the early history of biological control research but nevertheless provided some evidence that might have been recognized as implicating the fauna in disease suppression. Ever since then, the fauna have increasingly been recognized as potential suppressants of plant pathogens. Most of the research efforts on control of soil inhabiting pathogens with the fauna have focused on fungal pathogens. Viable propagules of Gaeumannonyces graminis var. tritici (take-all disease pathogen) decline more rapidly in "diseases suppressive soils" (Schneider, 1982) than in non-suppressive soils. Chakraborty et al. (1983) and Chakraborty (1985) revealed that mycophagous amoebae were strongly implicated when higher numbers were found associated with the pathogen in suppressive soils than in disease-conducive soils. Chakraborty and Warcup (1985) then demonstrated suppression of take-all disease of wheat in pot bioassays with introduced populations of Gephyramoeba, Saccamoeba and Thecamoeba.
Investigations of mycophagous nematodes for suppression of fungal pathogens and root disease have focused primarily on species of Aphelenchus and Aphelenchoides. Several root-infecting fungi have been effectively controlled in greenhouse environments using relatively large populations of Aphelenchus avenae. These include Pythium arrhenomanes on corn (Rhoades and Linford, 1959), R. solani on bean (Barker, 1964) and R. solani and Fusarium solani on alfalfa seedlings (Barnes et al., 1981). Aphelenchoides cibolensis and Aphelenchoides composticola suppressed Armillaria mellea and root-rot disease of penderosa pine (Pinus ponderosa) in agar-sand cultures (Riffle, 1973). Damage to wheat by Fusarium culmorum was significantly reduced in greenhouse pots infested with Aphelenchoides hamatus (Rossner and Nagel, 1984). Thus, these limited tests under artificial conditions provide ample evidence for potential disease suppression by the activities of mycophagous nematodes.
Collembola as plant pathogen suppressants have gained attention since the first research on Collembola and rhizosphere microflora interactions in relation_to cotton seedling disease of R. solani was conducted by Wiggins and Curl (1979). Subsequent experiments in large glass tubes of sterilized sandy loam and pots containing non-sterilized soil in greenhouse resulted in significant reduction of disease severity of R. solani and cotton seedling diseases by populations of P. minuta and O. encarpatus. Bollen et al. (1991) also observed that mesofauna suppression of Rhizoctonia disease of potato sprouts was less effective under conditions of high inoculum pressure. Results of studies in Japan (Nakamura et al., 1991) showed that Folsomia hidakana grazed and multiplied on R. solani and, when introduced into pots of soil, suppressed seedling diseases of radish, cucumber, cabbage and burdock. Further investigations by El Titi and Ulber (1991) provided evidence for collembolan suppression of other root diseases such P. ultimum on sugar beet seedlings. Application of an insecticide which resulted in reduced seedling emergence and increased infestation with Pythium in the experimental sugar beet plots, was attributed to the exclusion of mycophagous Collembola. In subsequent pot tests with sugar beet, the two authors (El Titi and Ulber, 1991) showed that Onychiurus fimatus could provide significant control of P. ultimum and reduce seedling losses.
While this discussion has so far focused on the activities of amoebae, nematodes and microarthropods, other small animals such as Enchytraeidae (pot worms) and Lumbricidae (earthworms) which occur in large numbers in various cropping systems could also indirectly affect plant pathogens (El Titi and Ulber, 1991). Stephens et al. (1993) demonstrated the potential of the earthworm Aporrectodea trapezoides that was applied to soil at the equivalent of 471m-2, to reduce the root-disease rating of R. solani on wheat. Additional tests with A. trapezoides and Aporrectodea rosea in both simulated system and field trials (Stephens et al., 1994) resulted in significant reductions in take-all (Gaeumannomyces graminis var, tritici) disease of wheat. Also, pot experiments demonstrated suppression of the clubroot pathogen, Plasmodiophora brassicae on cabbage seedlings by the earthworm Pheretima hilgendorfi at 110 m-2 (Nakamura et al., 1995).
Soil fauna mode of antagonism: Predation. The nature of antagonism of soil inhabiting plant pathogens by the fauna is direct predation (feeding). Micro- and mesoscopic species of soil fauna depend to a large extent on microflora as food sources in the rhizosphere and during organic matter decomposition. This creates an opportunity for the consumption of the pathogens and their propagules with resultant reduction of inoculum potential as defined by Garrett (1970). Interestingly, both earlier and subsequent discussions as well as analyses of research activities on crop sequence (Curl, 1963; Glynne, 1965) and organic amendments (King et al., 1934; Patrick and Toussoun, 1965; Papavizas, 1974) as biological control schemes offered no recognition that high populations of the micro- and mesofauna might have an interactive role with antagonistic microflora and help to explain the mechanisms of disease suppression or failure. As indicated, (Curl, 1982; Curl and Truelove, 1986; Curl and Harper, 1990) the role of microflora-feeding fauna in the disease-control process is now widely recognized.
An important criterion for effective biological control is the reduction of inoculum density that might result from feeding by these fauna and antagonism by other agents. Mitchell (1979) described absolute inoculum potential as a measure of the maximum capacity of a pathogen population to infect fully susceptible plant tissue under optimum conditions for infection. Consequently, the inoculum potential will vary with the inherent nature of different soilborne pathogens to produce propagules, to survive in soil and to infect host tissue.
It is therefore logical that the feeding activities of protozoa, nematodes and microarthropods in the soil could affect inoculum density. Old and Chakraborty (1986) thoroughly reviewed the biology and significance of mycophagous soil amoebae in relation to suppression of certain soilborne plant pathogens and disease. Prominent genera of fungus-feeding amoebae are Arachnula, Vampyrella, Cashia, Gephyramoeba, Mayorella, Saccamoeba, Thecamoeba and Theratromyxa. Selectivity or preference for certain food sources (fungi, bacteria and algae) is common among amoebae species. Feeding activity on fungal hyphae or spores is evidenced by perforation and lysis of cell walls (Howell, 1982), leaving uniform holes 0.2- 6.0 μm in diameter, or by general lysis of propagules taken into digestive vacuoles. Recognition of the destructive feeding habits of amoebae also led to subsequent investigations to determine the impact on inoculum density disease relationships by Anderson and Patrick (1978) who demonstrated that vampyrellid amoebae extracted from field soil in Ontario perforated chlamydospores and mycelium of Thielaviopsis basicola and conidia of Cochliobolus sativus resulting in reduced populations of both fungi.
Regarding nematodes, Aphelenchus avenae which was frequently found associated with roots and rhizomes was reared in vitro on a wide range of soil fungi and over 92 species were recorded as food sources. In laboratory tests Mankau and Mankau (1963) found that the common root-infecting fungi, including R. solani, Verticillium albo-atrum, F. solani and T. basicola were highly preferred diets for the reproduction and development of A. avenae. Aphelenchoides spp. which are largely polyphagous, feed and reproduce on a variety of phytopathogenic fungi and can be reared to large populations on specific fungi such as Agaricus campestris (Rossner and Nagel, 1984).
Many species of microarthropods (Acarina and Collembola) are mycophagous, feeding upon pathogens as well as saprophytes. However, soil mites as natural biological control agents have received little experimental attention. In one of the few such studies, Bollen et al. (1991) tested combinations of the nematode A. avenae, the Collembola Tul1bergia krausbaueri and mite species of Histiosoma, Tyrophagus and Rhizoglyphus for control of R. solani on potato sprouts and concluded that the mites were less effective than the other agents. However, the known microphagous feeding habits and high populations of the Cryptostigmata mite order in agroecosystems suggest a strong possible influence on inoculum density and disease potential which calls for more research efforts.
Studies on feeding habits of Collembola led to the investigation and recognition of P. minuta and O. encarpatus as potential agents for control of R. solani and cotton seedling disease by Curl (1979). In Japan, the collembolan Sinella curviseta from rhizosphere of vegetable crops grazed and multiplied on F. oxysporum f. sp· cucumerinum (Nakamura et al., 1992). Subsequent pot experiments then demonstrated the ability of the insect to suppress infection of cucumber by the pathogen. This subject matter has been extensively reviewed by Curl (1988), Curl et al. (1988) and Curl and Harper (1990).
Preferential feeding: Preferential feeding is crucial for successful application of soil fauna either alone or in combination with floral antagonists to control soil inhabiting pathogens. As mentioned earlier, Mankau and Mankau (1963) pointed out that some common root-infecting fungi were highly favorable food source for A. avenae. They found, however, that abundantly sporulating fungi such as Aspergillus terreus, Penicillium spp., Rhizopus nigricans and Trichoderma viride were not favorable food sources; some species of Pythium and Phytophthora also were poor food sources and apparently were toxic. Subsequent studies by Cayrol and Combettes (1977) provided supporting evidence that both A. avenae and Aphelenchoides composticola were aggressive and inhibitory on cultures of R. solani, but neither inhibited growth of Pythium ultimum.
The role of preferential feeding of the soil fauna in disease suppression has gained significant attention in recent years. It was previously thought that Collembola had no preferential feeding habit. Gut-content analyses of field-collected Collembola, for example, reflect non-specific feeding habits (Poole, 1959; Petersen, 1971; Anderson and Healy, 1972; Visser and Whittaker, 1977; Takeda and Ichimura, 1983). However, preferential feeding upon specific fungi has been demonstrated in several laboratory experiments (Mills and Sinha, 1971; McMillan, 1976; Visser, 1985; Shaw, 1988; Lartey et al., 1989; Sabatini and Innocenti, 2000) and to a limited extent in field tests (Newell, 1984; Shaw, 1985). As demonstrated by Lartey et al. (1989), the basis for preferential feeding may be influenced by several factors. In one such test, a strong feeding preference by P. minuta and O. encarpatus for the damping off pathogen R. solani in the presence of T. harzianum, T. virens, or L. arvalis was demonstrated. When the pathogen was omitted in subsequent tests with P. minuta, there was a clear choice for L. arvalis as the preferred food over T. harzianum or T. virens and greater egg production occurred on the basidiomycete. With no other food available, the Collembola showed no preference between T. harzianum and T. virens and grazed lightly on hyphal tips up to time of abundant sporulation, which repelled the collembolan insects. Unwashed spores were lethal when ingested by P. minuta, whereas washed spores could be ingested without apparent harm (Lartey et al., 1994). Compatibility of fungal food sources for collembolans also was reflected in the reproductive capacity of P. minuta in monocultures on R. solani, T. harzianum, T· virens and L. arvalis. After 12 days, recovered animals from cultural tubes of cornmeal and sterile soil substrate were highest with cultures of either R. solani or L. arvalis. Lartey et al. (1989) concluded that the preference for R. solani was a matter of choice and in the absence of the pathogen, L. arvalis was the alternate choice. In addition to probable distaste for the Trichoderma species, production of toxic metabolites during sporulation appears also to have influenced the aversion for the Trichoderma species by the collembolan insects. Other observations by Visser and Whittaker (1985) indicated that Onychiurus subtenuis preferred dark-pigmented fungi such as Cladosporium spp. over hyaline basidiomycetes, whereas 0. armatus showed a preference for the basidiomycete Lactarius rufus over several other species from a pine forest. Bengtsson et al. (1988) suggested that some Collembola, as in this case 0. armatus, may locate preferred food sources by odor attraction of volatile compounds released from mycelium.
INTEGRATION OF AGENTS FOR BIOCONTROL
Combinations of two or more biological control agents which are compatible at the target environ such as spermosphere-soil interface, the root-soil interface or the bulk soil should provide enhanced protection from pathogen activity. Some strains of Fluorescent pseudomonads when combined in greenhouse, growth chamber and field tests resulted in enhanced suppression of take-all Gaeumannomyces graminis var. tritici and improved wheat growth (Pierson and Weller, 1994). Treatment with some individual strains provided neither enhanced control nor yield improvement. Control of Fusarium wilt of radish caused by F. oxysporum was enhanced by combining two strains of P. pituda (WC358) with apparently different mechanisms (de Boer et al., 2003) Bollen et al. (1991) used combinations of mycophagous nematodes, Collembola and mites from potato fields to demonstrate enhanced control of R. solani on potato sprouts in pots of sandy soil. A combination of the nematode A. avenae and the collembolan Folsomia fimetaria was more effective than either alone in disease suppression. Similarly, A. avenae and Tullbergia krausbaveri together, or these two combined with mite species, provided superior disease control.
Collembolan P. minuta and the fungus L. arvalis were evaluated for suppression of R. solani on cotton seedlings in greenhouse tests (Lartey et al., 1991).
| Table 1: | Control of Rhizoctonia solani on cotton in nonsterilized soil by integration of Laetisaria arvalis seed treatment and a population of collembolan Proisotoma minuta (Lartey et al., 1991) |
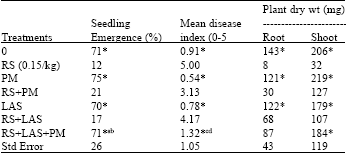 | |
| Means (8 replicate pots) emergence, disease severity and growth (root and shoot dry weights) of cotton seedlings grown in Rhizoctonia solani (RS)(0.15g/kg soil) supplemented nonsterilized soil, that were treated with Proisotoma minuta (PM) (1000/kg soil) and seed applied Laetisaria arvalis (LAS) alone or combined, 0 = Untreated nonsterilized soil | |
| *Mean values differ from control RS, (P = 0.05, Dunnett's test.) a Mean value differs from RS+PM (P = 0.01, contrast comparison) b Mean value differs from RS+LAS (P = 0.01, contrast comparison) c Mean value differs from RS+PM (P = 0.1, contrast comparison) d Mean value differs from RS+LAS (P = 0.05, contrast comparison) | |
The combination of the agents enhanced the suppression of the pathogen in sterilized or non-sterilized soil as compared with either agent used alone; the benefit increased further as numbers of animals applied were increased (Table 1). Continuing studies of this type (Lartey et al., 1994) also demonstrated the added benefit of combining P. minuta populations with T. harzianum or T. virens for control of R. solani, with the combination of L. arvalis and P. minuta treatment, providing the most effective result. The specific mechanisms that enhanced biocontrol by the mycophagous fauna and the fungal combinations were not clearly defined, but must relate to food preference of the animal for the pathogen, thus permitting both Collembola and fungal biocontrol agents to function together.
CONCLUSIONS AND CHALLENGES
Soil inhabitants, microflora and micro- and mesofauna provide an enormous potential for natural suppression of inoculum, disease incidence and severity. The key to understanding the phenomena and the associated potential lies in understanding the soil as an ecosystem and interactions among the potential antagonists and the target pathogens. Current research has concentrated on the rhizosphere in particular and to some degree on the spermosphere. Understandably these research efforts have been directed specifically at soilborne pathogens, disregarding other soil inhabiting pathogens in the bulk soil.
While laboratory and greenhouse experiments have demonstrated that specific microbial antagonists can reduce inoculum density and lower the disease index in controlled environments, more meaningful biological control might be induced through the management of indigenous populations, as visualized by Cook (1991). The soil microflora and fauna respond to pesticides, soil fertility, organic matter, cultivation practices and crop species, thus providing opportunities for agroecosystem manipulation to promote antagonists for crop benefit. Emerging technologies such as ELISA could be applied to monitor accurately the activities of antagonists and induced changes in target soil inhabiting pathogen populations and inoculum density (Caesar-TonThat et al.,). Such a technology, employing PCR for rapid detection of pathogens and infection in plant tissues (Lartey et al., 2003) could also be used to evaluate new approaches to control surviving propagules of folia pathogens such as C. beticola with biological agents in the soil.
Some soil microbes, such as the algae and archaea have not been explored for their potential to control plant pathogens. The algae, however, have been documented to produce a variety of toxins (Krishnamurthy et al., 1989; Katircioglu et al., 2004). Some of these toxins may well serve to provide protection against potential enemies and thus the algae may serve as antagonists against some soil inhabiting plant pathogens. The ability of the archaea to thrive in extremely harsh environments may be indicative of their competitive ability. It is therefore feasible to explore these organisms for any potential to control soil inhabiting plant pathogens. While no information is available on control of bacterial pathogens with fungal antagonists, information is available on control of bacteria by fungi, recent studies have shown that cercosporin, produced by Cercospora spp. can inhibit growth of E. coli. This observation presents an opportunity to explore the possibility of applying some fungal antagonists and their metabolic products to control bacteria and other soil inhabiting pathogens. Productions of toxic metabolites are not limited to antagonists but are also products of pathogens and may well provide protection from antagonists. Successful antagonism against such pathogens may require ability to detoxify these protective toxins by applied agents, as observed with L. arvalis (Caesar-TonThat et al., 2002). These will have to be considered during search for potential biological agents against such pathogens. In concluding, it is obvious that new challenges lie ahead for the exploitation of these untested "friendly” soil flora, fauna and technologies for the benefit of crops.
ACKNOWLEDGMENTS
I thank Dr. Elizabeth A. Pierson, Division of Plant Pathology and Microbiology in The Dept. of Plant Sciences, University of Arizona, Tucson, AZ, USA, Dr. Marc A. Cubeta, Dept of Plant Pathology, North Carolina State University, Raleigh, NC. USA and Dr. Kenneth E. Conway, Dept of Entomology and Plant Pathology, Oklahoma State University, Stillwater OK, USA for review of the manuscript and excellent suggestions.
REFERENCES
- Beagle-Ristaino, J.E. and G.C. Papavizas, 1985. Biological control of Rhizoctonia stem canker and black scurf of potato. Phytopathology, 75: 560-564.
CrossRefDirect Link - Cronin, D., Y. Moenne-Loccoz, A. Fenton, C. Dunne, D.N. Dowling and F. O'Gara, 1997. Ecological interaction of a biocontrol Pseudomonas fluorescens strain producing 2,4-diacetylphloroglucinol with the soft rot potato pathogen Erwinia carotovora subsp. atroseptica. FEMS Microbiol. Ecol., 23: 95-106.
CrossRefDirect Link - Elad, Y., I. Chet and J. Katan, 1980. Trichoderma harzianum: A biocontrol agent effective against Sclerotium rolfsii and Rhizoctonia solani. Phytopathology, 70: 119-121.
CrossRefDirect Link - Gottlieb, D. and P.D. Shaw, 1970. Mechanism of action of antifungal antibiotics. Ann. Rev. Phytopathol., 8: 371-402.
CrossRef - Hiltner, L., 1904. Uber neuere Erfahrungen und Probleme auf dem Gegiet der Bodenbakteriology und unter besonderer Berucksichtigung der Grundungung und Brache (On new findings and problems in the area of soil bacteriology with special consideration of green manure and fallow) Arb. Deut. Landw. Gesell, 98: 59-78.
- Katircioglu, H., B.S. Akin and T. Atici, 2004. Microalgal toxin(s): Characteristics and importance. Afr. J. Biotechnol., 3: 667-674.
Direct Link - Lartey, R.T., E.A. Curl, C.M. Peterson and J.D. Harper, 1989. Mycophagous grazing and food preference of Proisotoma minuta (Collembola: Isotomidae) and Onychiurus encarpatus (Collembola: Onychiuridae). Environ. Entomol., 18: 334-337.
Direct Link - Lartey, R.T., E.A. Curl, C.M. Peterson and J.C. Williams, 1991. Control of Rhizoctonia solani and cotton seedling disease by Laetisaria arvalis and a mycophagous insect Proisotoma minuta (Collembola). J. Phytopathol., 133: 89-98.
Direct Link - Lartey, R.T., E.A. Curl and C.M. Peterson, 1994. Interactions of mycophagous Collembola and biological control fungi in the suppression of Rhizoctonia solani. Soil Biol. Biochem., 26: 81-88.
CrossRefDirect Link - Mallow, D., R.J. Snider and L.S. Robertson, 1985. Effects of different management practices on Collembola and Acarina in corn production systems. II: The effects of moldboard plowing and atrazine. Pedobiologia, 28: 115-131.
Direct Link - Morgan-Jones, G., G. Godoy and R. Rodriguez-Kabana, 1981. Verticillium chlamydosporium, fungal parasite of Meloidogyne arenaria females. Nematropica, 11: 115-120.
Direct Link - Oskay, A.M., T. Usame and A. Cem, 2004. Antibacterial activity of some actinomycetes isolated from farming soils of Turkey. Afr. J. Biotechnol., 3: 441-446.
Direct Link - Rouatt, J.W. and H. Katznelson, 1961. A study of the bacteria on the root surface and in the rhizosphere soil of crop plants. J. Applied Bacteriol., 24: 164-171.
CrossRefDirect Link - Shaukat, S.S. and I.A. Siddiqui, 2003. Zinc improves biocontrol of Meloidogyne javanica by the antagonistic Rhizobia. Pak. J. Biol. Sci., 6: 575-579.
CrossRefDirect Link - Singh, P.P., Y.C. Shin, C.S. Park and Y.R. Chung, 1999. Biological control of Fusarium wilt of cucumber by chitinolytic bacteria. Phytopathology, 89: 92-99.
PubMed - Whipps, J.M., 2001. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot., 52: 487-511.
PubMedDirect Link - Woese, C.R., O. Kandler and M.L. Wheelis, 1990. Towards a natural systemof organisms. Proposal for the domains archaea, bacteria and eucarya. Proc. Nat. Acad. Sci. USA., 87: 4576-4579.
Direct Link - Bollen, G.J., J. Middelkoop and T.W. Hofman, 1991. Effects of Soil Fauna on Infection of Potato Sprouts by Rhizoctonia solani. In: Biotic Interactions and Soilborne Diseases, Beemster, A.B.R., G.J. Bollen, M. Gerlagh, M.A. Ruissen, B. Schippers and A. Tempel (Eds.). Elsevier Scientific Publishing, Amsterdam, pp: 27-34.
- Curl, E.A., R.T. Gudauskas, J.D. Harper and C.M. Peterson, 1985. Effects of Soil Insects on Populations and Germination of Fungal Propagules. In: Ecology and Management of Soilborne Plant Pathogens, Parker, C.D., A.D. Rovira, K.J. Moore, P.T.W. Wong and J.F. Kollmorgen (Eds.). American Physiological Society, St. Paul, MN., pp: 20-23.
- El Titi, A. and B. Ulber, 1991. Significances of Biotic Interactions between Soil Fauna and Microflora in Integrated Arable Farming. In: Biotic Interactions and Soil-Bourne Diseases, Beemster, A.B.R., G.J. Bollen, M. Gerlagh, M.A. Ruissen, B. Schippers and A. Tempel (Eds.). Elsevier Scientific Publishing, Amsterdam, pp: 1-19.
- Honeycutt, E.W. and D.M. Benson, 2001. Formulation of binucleate Rhizoctonia spp. and biocontrol of Rhizoctonia solani on impatiens. Plant Dis., 85: 1241-1248.
Direct Link - Shaw, P.J.A., 1985. Grazing Preferences of Onychiurus armatus (Insecta: Collembola) For Mycorrhizal and Saprophytic Fungi of Pine Plantations. In: Ecological Interactions in Soil: Plants, Microbes and Animals, Fitter, A.H., D. Atkinson, D.J. Read and M.B. Usher (Eds.). Blackwell Scientific Publishing, Oxford, pp: 333-337.
- De Boer, M., P. Bom, F. Kindt, J.J.B. Keurentjes, I. van der Sluis, L.C. van Loon and P.A.H.M. Bakker, 2003. Control of Fusarium wilt of radish by combining Pseudomonas putida strains that have different disease-suppressive mechanisms. Phytopathology, 93: 626-632.
CrossRefDirect Link - Jacq, V.A. and R. Fortuner, 1979. Biological control of rice nematodes using sulphate reducing bacteria. Revue Nematol., 2: 41-50.
Direct Link