
D. Dhanasekaran
Department of Microbiology, Adhiparasakthi College of Science, G.B. Nagar, Kalavai, Vellore District, Tamil Nadu, India
P. Sivamani
Department of Microbiology, Adhiparasakthi College of Science, G.B. Nagar, Kalavai, Vellore District, Tamil Nadu, India
A. Panneerselvam
P.G. Research Department of Botany and Microbiology,
AV.V.M. Sri Pushpam College, (Autonomous), Poondi, 613 503, Thanjavur District, Tamil Nadu, India
N. Thajuddin
Department of Microbiology, National Faculty for Marine Cyanobacteria,
(Sponsored by DBT, Govt. of India), Bharathidasan University, Trichirapalli, 620 024, Tamil Nadu, India
G. Rajakumar
1Department of Microbiology, Adhiparasakthi College of Science, G.B. Nagar, Kalavai, Vellore District, Tamil Nadu, India
S. Selvamani
Department of Microbiology, Adhiparasakthi College of Science, G.B. Nagar, Kalavai, Vellore District, Tamil Nadu, India
Plant Pathology Journal
Year: 2005 | Volume: 4 | Issue: 2 | Page No.: 91-95







ABSTRACT
Twenty six antibiotic producing Streptomyces were isolated from soils of Cuddalore Tamil Nadu, India and were evaluated for their ability to inhibit plant pathogenic Rhizoctonia solani in vitro. It was found that the isolates having pathogenic inhibitory ability and also controls Rhizoctonia solani damping off in tomato in interested field soils. The Streptomyces isolates tested significantly reduced damping off severity in tomato. A variation in their effect on plant disease severity, percentage dead plants and plant biomass in the presence of the pathogen, was observed among the isolates.
PDF Abstract XML References Citation
How to cite this article
D. Dhanasekaran, P. Sivamani, A. Panneerselvam, N. Thajuddin, G. Rajakumar and S. Selvamani, 2005. Biological Control of Tomato Seedling Damping off with Streptomyces
sp. Plant Pathology Journal, 4: 91-95.
DOI: 10.3923/ppj.2005.91.95
URL: https://scialert.net/abstract/?doi=ppj.2005.91.95
DOI: 10.3923/ppj.2005.91.95
URL: https://scialert.net/abstract/?doi=ppj.2005.91.95
INTRODUCTION
Chemicals that either have direct fungicidal activity against the pathogen or converted into toxic derivatives by the pathogen or host plant tissue often control plants fungal diseases. These are highly efficient, systemic fungicides, including the triazoles and acylalanines, which effectively control some plant diseases[1]. However, the intensive use of systemic fungicides has not only created problems of fungicide resistance and increased contamination of the environment but may also have adverse mammalian health effects[2,3]. In recent years, studies on the biological control of plant diseases, particularly using antibiotic metabolites of microbial origin, have been expanded and such materials may supplement to be an alternative to chemical disease control[4]. Some of the microbial secondary metabolites have highly selective activity against particular plant pathogen and have little effects on beneficial organisms. These metabolites often do not accumulate in nature[5].
Streptomyces sp. is gram-positive filamentous bacteria that produce and secrete a wide array of biologically active compounds including antibiotics, hydrolytic enzymes (Proteases, Lipases) and enzyme inhibitors. They are resistant to desiccation and nutrient stress, by their ability to produce spores. These characteristics make Streptomycetes attractive candidates for biological control agents against soil-borne plant pathogens[6]. Streptomycetes have been implicated in antagonism of a variety of plant pathogens. In petri-plate assays, a number of species produce compounds that inhibit the growth of plant-pathogenic fungi[7,8]. When inoculated into soil (or) on seeds, many Streptomycetes protect plants from fungal diseases[9,10].
The objectives of the present study were to identify antagonistic actinomycetes strains from 26 isolates and purify the antibiotic substances from their culture filtrates. This study also reports the effect of Streptomyces culture filtrate on growth of Rhizoctonia solani, a causative agent of damping off in tomato.
MATERIALS AND METHODS
Bacterial and fungal strains: Twenty six strains Streptomyces were isolated from a marine soil in Cuddalore, Tamilnadu, India, pathogenic Rhizoctonia solani were stored until use in 20% glycerol and silica gel at 4°C, respectively.
Identification of Streptomyces isolates: All the isolated organisms proved to be aerobic, spore-forming, gram positive and formed extensive branched hyphe. They have L-diaminopimelic acid (DAP) in their whole cell hydrolystate and produced a wide variety of pigments responsible for the colouration of aerial and substrate mycelia and produced antimicrobial agents. These results confirmed that these isolates belong to the genus Streptomyces.
In vitro antibiosis assay: A double layer agar method was used to determine in vitro inhibition of Rhzoctonia solani by 26 Streptomyces strains. In order to determine the potential of Streptomyces strains to influence the production of antimicrobial compounds by other in culture, antibiosis assays were carried out initially with one Streptomyces strain per agar plate and results were compared with assays in which Rhizoctonia solani was cultured on each agar plate. Spores of Streptomyces strains were obtained from 10 days old sporulation agar culture, spot plated on to inorganic salt starch agar medium[11] at 1x104 to 1x105 Colony Forming Unit (CFU) and incubated at 28°C for 3 days. Inverting the plates over 3 mL of chloroform for 1 h killed the bacteria. After drying for 30 min, the plates were overlaid with 15 mL molten 1% water agar containing a suspension of Rhizoctonia solani tested. Following 3 to 5 days of incubation at 24°C, the plates were evaluated for growth inhibition of the pathogens, as observed as clear zones around the spot of Streptomyces, by recording the diameter of the inhibition zone. The pathogen was tested on three plates and the experiment was repeated three times.
Bioassay for Rhizoctonia solani damping-off in tomato: The rolled paper towel assay was used to evaluate the efficacy of coating tomato seeds with spores of Streptomyces strain to control infection of tomato seedlings by Rhizoctonia solani. Streptomyces spore suspensions were obtained by flooding 10 day-old starch casein agar plate cultures with sterile distilled water. Suspensions were diluted with sterile distilled water to obtain a range of spore concentrations for coating seeds. Surface sterilized seeds of tomato were soaked for 30 min in Streptomyces spore suspensions and air-dried in a laminar flow hood. To determine the number of spores present on the seed coat after inoculation, a sample of seed was agitated in sterile distilled water and an aliquot of the resulting suspension was plated onto starch casein agar. The number of Streptomyces colonies was determined after 10 days of incubation at 28°C. The Rhizoctonia solani inoculum was produced by infesting 1 kg sterilized soil amended with 5 g kg-1 finely chopped 3 day-old Potato dextrose agar culture of the fungus. Infested soil was incubated at room temperature for 10 days and then refrigerated at 5°C until used. Inoculum density of Rhizolonia solani in this soil was 3-6 CFU g-1 soil. Seeds were spread on a moistened sterile 26 x 30 cm paper towel, 20 seeds per towel and covered with 10 g of the Rhizotonia solani infested soil. The towels were rolled, placed in plastic bags and incubated for 5 days at room temperature. Three towels were used per treatment and the experiment was carried out three times. After incubation, disease severity was rated using a 5-class scale.
| 1 | = | Health seedling |
| 2 | = | Primary root tip necrotic but firm |
| 3 | = | Primary root tip soft and rotted |
| 4 | = | Dead seedling, germinated seed with rotted radical |
| 5 | = | Dead seed, ungerminated rooted seed and expressed as the average disease severity index (ASI) described by Altier and Thies[12]. |
Biocontrol of Rhizoctonia solani root rots of tomato with Streptomyces isolates in naturally infested soil.
Seeds of tomato and spore suspensions of Streptomyces isolates were prepared as described above. The soil used for each experiment was mixed thoroughly and put into 8 cm diameter pots. Pots were watered one day before sowing. A total of three treatments were evaluated for each crop [five isolates of Streptomyces and one an antagonist control]. Five tomato seeds were placed into each pot. One millimeter of spore suspension of Streptomyces was inoculated directly over seed immediately after sowing. Control for tomato was treated with corresponding volume of sterile starch casein broth only. Seeds were covered with soil following inoculation of Streptomyces. Water saturated conditions were kept during the first 10 days to encourage infection and then followed by 4 to 5 days of dry conditions to stimulate drought stress in the field. All pots were subsequently watered when soil surface was dry.
Seedlings were harvested by carefully removing them from the soil, washing gently with tap water and blotting dry with paper towels. Disease incidence, disease severity, seedling height, fresh weight and root length were measured for every seedling in each pot as described previously.
RESULTS
In vitro inhibition potential of Streptomyces sp.: Of the 26 isolates tested, only six isolates showed were able to inhibit the growth of Rhizoctonia solani in vitro. The isolates namely DPTB110 and DPTB13 maximum inhibition zones (17 mm) against Rhizoctonia solani. Hence these two isolates were selected from the above isolates for further study based on the mean size of the inhibition zone against the pathogen (Table 1).
| Table 1: | In vitro antifungal activity of Streptomyces |
 | |
| Table 2: | Activity Streptomyces isolates in sterilized soil |
 | |
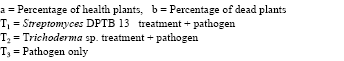 | |
Biocontrol activity of antibiotic producing Streptomyces isolates in sterilized soil: The Streptomyces isolates with the lowest percentage of dead plants (DPTB 110 and BPTB13) were tested further for their ability to reduce disease and enhance emergence of tomato seedlings in two subsequent experiments performed under two pathogen inoculum concentration. At the higher inoculums concentration Streptomyces isolate DPTB13 significantly reduced the percentage of dead plants and increased the percentage of healthy plants as compared with the pathogen only control. Under the lower inoculums concentration, two Streptomyces isolates significantly reduced the percentage of dead plants and increased the percentage of healthy plants compared with the pathogen only treatment combination of Streptomyces and commercial fungicide were more effective. The results were similar to positive control (Table 2).
Bio control of Rhizoctonia solani on tomato with Streptomyces in naturally infested soil: Streptomyces isolates had no effect on the emergence of tomato seedlings compared with the control treatment in naturally infested soil. Streptomyces isolate DPTB13 significantly reduced the percentage of dead plants and increased the percentage of healthy plants in experiment I as compared with non-inoculated control. In the same experiment the Streptomyces isolate DPTB13 significantly increased the mean plant fresh weight. Similar results were obtained from a repeated of experiment though the disease percentage was lower in the second experiment than in the first experiment (Table 3).
Effect of antibiotic producing Streptomyces isolates on tomato seedlings: Streptomyces isolate DPTB 13 significantly increased mean weight, height; taproot length of tomato seedlings grown in sterilized soil as compared with the non inoculated control. Between the two Streptomyces isolates tested, the isolate DPTB110 did not provide any plant growth benefit where compared with the DPTB13. Similar results were also obtained in a repeat of this experiment (Table 4).
DISCUSSION
Different Streptomyces strains isolated from Cuddalore, effectively reduced Rhizoctonia solani growth on tomato and increased the proportions of healthy plants. Of the 26 Streptomyces isolates, the DBTB13 isolate was most effective Streptomyces isolate in bio control of Rhizoctonia solani infected with tomato, in naturally infested soil (Table 2). In addition, this isolate produced larger average inhibition zones (17 mm) against Rhizoctonia solani in vitro. These data suggest that, in addition to its potential for controlling Rhizoctonia solani on tomato, Streptomyces isolate may also be useful in integrated control against diverse soil born plant pathogens.
Both antagonists and pathogen inoculum densities had a substantial effect on disease control. Generally, disease control was better at higher antagonist inoculums densities, though there were limits to the effectiveness of increasing antagonist’s densities. Determining optimal antagonist inoculum densities will require further comparative antagonist inoculums doses in the presence of varying pathogen densities (Table 2).
Although Streptomyces isolates selected based on in vitro antibiotic activity were effective in controlling Rhizoctonia solani in sterilized soil and infested field soil, the size of in vitro inhibition zones for individual Streptomyces isolates against pathogen was not significantly correlated with successful biocontrol of those pathogen (Table 3 and 4). This result is consistent with the findings of other antagonistic microorganisms have been reported to cause a decline in the population of plant pathogens[13]. Sneh et al.[14] suggesting that in vitro antibiotic assays provide at best an imperfect means for identifying antagonists.
There is no significant positive correlation between tomato seedlings growth when inoculated with antagonists in sterile soil in the absence of the pathogen and those when inoculated with antagonists in pathogen-infested field soil was noted. Overall, the strongest predictor of tomato plant growth when inoculated with an antagonist and grown in pathogen-infested field soil was tomato biomass following inoculation on of the same antagonist in sterile soil.
| Table 3: | Effect of Streptomyces on Rhizoctonia solani grown naturally infested soil tomato seedlings |
 | |
| Table 4: | Effect of Streptomyces against Rhizoctonia solani in tomato seedlings |
 | |
In this case, direct enhancement of tomato growth by Streptomyces may be one of the mechanisms by which Streptomyces antagonists enhance plant health in field soil. The relationship of plant growth promoting rhizobacteria to plant growth has been discussed by Kloepper and Schroth[15] (Table 3 and 4). Thus, antagonist-screening approaches that consider plant growth in the absence of any pathogens may provide an alternative a complement to more traditional antibiotic screening assays. Further research is underway to determine the mechanisms of growth enhancement of tomato by Streptomyces antagonists.
Rhizoctonia solani varied significantly in their sensitive to inhibition by different Streptomyces isolates. This suggests that a single isolate will be unlikely to provide broad-spectrum control of diverse soil borne plant pathogens. Furthermore, many factors are, likely to effect biological control of damping off, including environmental conditions such as soil temperature, morphology, water status and nutrient availability and pathogen population densities. Indigenous soil microbes can also interact with inoculated antagonists, thereby influencing their activities in soil. Consequently, it seems unlikely that a single antagonist will be capable of providing consistent and long-lasting control in different field locations under varying environmental conditions[16,17]. The future of the present research is the development of integrated management scheme for soil borne plant pathogen that incorporates combinations of complementary Streptomyces isolates, resistant plant cultivars and cultural practices including tillage and rotational schemes that integrated green manures optimal for the management of antagonists population in soil.
REFERENCES
- Crawford, D.L., J.M. Lynch, J.M. Whipps and M.A. Ousley, 1993. Isolation and characterization of actinomycete antagonists of a fungal root pathogen. Applied Environ. Microbiol., 59: 3899-3905.
Direct Link - Jones, C.R. and D.A. Samac, 1996. Biological control of fungi causing alfalfa seedling damping-off with a disease-suppressive strain of Streptomyces. Biol. Control, 7: 196-204.
CrossRefDirect Link - Harman, G.E., I. Chet and R. Baker, 1980. Trichoderma hamatum effects on seed and seedling disease induced in radish and pea by Pythium spp. or Rihizoctonia solani. Phytopathology, 70: 1167-1172.
Direct Link - Kloepper, J.W. and M.N. Schroth, 1981. Relationship of in vitro antibiosis of plant growth-promoting rhizobacteria to plant growth and the displacement of root microflora. Phytopathology, 71: 1020-1024.
CrossRefDirect Link - Jones, C.R. and D.A. Samac, 1996. Biological control of fungi causing alfalfa seedling damping-off with a disease-suppressive strain of Streptomyces. Biol. Control, 7: 196-204.
CrossRefDirect Link - Samac, D.A., A.M. Willert, M.J. Mcbride and L.L. Kinkel, 2003. Effects of antibiotic producing Streptomyces on nodulation and leaf spot in alfalfa. Applied Soil Ecol., 22: 55-66.
CrossRefDirect Link