
Gholam Hosein Shahidi Bonjar
Department of Plant Pathology, College of Agricultural Sciences, Bahonar University of Kerman, Iran
Parvin Rashid Farrokhi
Department of Agronomy and Plant Breeding, College of Agricultural Sciences,
Bahonar University of Kerman, Iran
Sonia Aghighi
Department of Plant Pathology, College of Agricultural Sciences, Bahonar University of Kerman, Iran
Leyla Shahidi Bonjar
Department of Plant Pathology, College of Agricultural Sciences, Bahonar University of Kerman, Iran
Ataollah Aghelizadeh
Management and Planning Organization of Kerman Province, Kerman, Iran
Plant Pathology Journal
Year: 2005 | Volume: 4 | Issue: 1 | Page No.: 78-84







ABSTRACT
Antifungal actinomycete strains were assayed for antagonistic activity against Verticillium dahliae, Alternaria solani, Fusarium solani and Geotrichum candidum, four worldwide phytopathogenic fungi. For better evaluation of the spectral activity of the isolated Actinomycetes, two yeasts, Saccharomyces cerevisiae and Candida albicans were also included in the survey. From 110 soil inhabitant strains that have been isolated from soil samples of Kerman province, southeast of Iran, 10 strains showed antifungal activities as determined through screening and bioassays by agar disk and well-diffusion methods. Active strains that have broad spectrum of antifungal activity were grown in submerged cultures and active crude was prepared upon which some preliminary characterizations conducted.
PDF Abstract XML References
How to cite this article
Gholam Hosein Shahidi Bonjar, Parvin Rashid Farrokhi, Sonia Aghighi, Leyla Shahidi Bonjar and Ataollah Aghelizadeh, 2005. Antifungal Characterization of Actinomycetes Isolated from Kerman, Iran and Their Future Prospects in Biological Control Strategies in Greenhouse and Field Conditions. Plant Pathology Journal, 4: 78-84.
DOI: 10.3923/ppj.2005.78.84
URL: https://scialert.net/abstract/?doi=ppj.2005.78.84
DOI: 10.3923/ppj.2005.78.84
URL: https://scialert.net/abstract/?doi=ppj.2005.78.84
INTRODUCTION
In modern fungal disease-management of plants, there are constant needs for novel plant chemotherapeutic agents different from the currently used fungicides in mode of action and chemical structures. The most important reason for these demands includes problems of fungicide resistance, which result in high application rates of many synthetic fungicides and deleterious effects on environments in consequence[1-3]. Since the systematic screening programs in the search for antimicrobial metabolites were first performed by Waksman in 1940s, microbial cultures have been a major source of antibiotic substances[4]. Actinomycetes are one of the most attractive sources of antibiotics and other biologically active substances of high commercial value, such as vitamins, alkaloids, plant growth factors, enzymes and enzyme inhibitors[5]. Their ability to parasitize and degrade spores of fungal plant pathogens is well established[6]. They are able to metabolize many different compounds including sugars, alcohols, amino acids and aromatic compounds by producing extracellular hydrolytic enzymes. Their metabolic diversity is due to their extremely large genome which has hundreds of transcription factors that control gene expression, allowing them to respond to specific needs[7]. Streptomyces species are the most widely studied and well known of the actinomycete. Soil Streptomyces are of the major contributors to the biological buffering of soils and have roles in decomposition of organic matter conductive to crop production. Besides, they have been much studied as potential producers of antibiotics and exert antagonistic activity against wide range of bacteria and fungi[4,8,9]. These prokaryotes have been much studied as potential producers of antibiotics[4,10]. The results even show that use of Streptomyces enhances growth of the crop plants[11]. With the increased concern about conserving natural resources as air, soil and water, natural or biological control of plant diseases has received increased emphasis. Biological control of plant diseases is slow, gives few quick profits, but can be long lasting, inexpensive and harmless to life. Biocontrol systems do not eliminate neither pathogen nor disease but bring them into natural balance[12]. Phytopathogenic fungi are of major problems in agriculture. Alternaria solani Sorauer, Fusarium solani (Mart.) Sacc., Verticillium dahliae Klebahn and Geotrichum candidum Link ex Pers. are ubiquitous and cosmopolitan phytopathogens causing severe diseases in wide range of plant species[13-15]. With extended environmental diversity, however, the actinomycete s microflora of the Iranian soils has not been very well explored with the goal of exploring new means of biocontrols. The merits of their role in biological control of soil-borne fungal-pathogens, at the present research 110 isolates of actinomycetes were isolated from agricultural soils of Kerman province, Iran and screened against A. solani, F. solani, V. dahliae and G. candidum. For better evaluation of the spectral activity of the isolated actinomycete, two yeasts, Saccharomyces cerevisiae Meyen ex Hansen and Candida albicans (Robin) Berkhout was also included in the survey. S. cerevisiae is commonly known as brewer's yeast, but has also been reported as the cause of both superficial and invasive infections in humans [16]. The objective of the present study was to isolate actinomycetes strains having antagonistic properties against some of the important fungi with the aim that they can serve as gene donors in developing resistant transgenic plants or use as soil amendments in biological control of the plant pathogens. From 110 isolates of actinomycetes, 10 isolates showed antifungal activity at least against one of the tested fungi.
MATERIALS AND METHODS
Culture media: Casein glycerol (or starch) agar (CGA) was used for screening and isolating of actinomycete which composed of: glycerol or soluble starch, 10 g; casein, 0.3 g; KNO3, 2 g; NaCl, 2 g; K2HPO4, 2 g; MgSO4.7H2O, 0.05 g; CaCO3, 0.02 g; FeSO4.7H2O, 0.01 g and agar, 18 g in 1 L of distilled H2O (pH 7.2)[12]. In submerged cultures, agar was excluded (CG medium). Actinomycete colonies with different morphologies were selected and transferred to CGA slants for further studies.
Preparation of fungal isolates: Tested fungi were kind gifts as follows: A. solani (tomato isolate) from Dr. Zare, Mycology Lab., Plant Pests and Diseases Research Institute, Evin, Tehran; G. candidum (tomato isolate) from Mycology Lab., Department of Plant Pathology, College of Agricultural Sciences, Bahonar University of Kerman, Kerman; F. solani (potato isolate) from Dr. Sharifnabi, Department of Plant Pathology, College of Agricultural Sciences, Isfahan University of Technology; V. dahliae (cotton, potato and olive isolates) from Professor Banihashemi, Mycology Lab., Department of Plant Pathology, College of Agriculture, Shiraz University, Shiraz; V. dahliae (Pistachio isolates) from Mr. Aminaii, Plant Pathology Lab., Agricultural Research Center, Ministry of Agriculture, Kerman, Iran. The fungi were grown at 25-26°C and maintained on Potato Dextrose Agar (PDA) (Difco). S. cerevisiae (PTCC No. 5052) from Persian Type Culture Collection (PTCC), Tehran and C. albicans from Mycology Lab., Department of Microbiology, University of Medical Sciences, Kerman, Iran and grown at 30°C on Sabouraud dextrose medium. All cultures stored at 4°C and sub-cultured as needed.
Isolation of actinomycete from soil: Soil samples were collected from grasslands, orchards and vegetable fields in different localities of Kerman province, Iran. Several samples randomly were selected from mentioned localities using an open-end soil borer (20 cm in depth, 2.5 cm in diameter) as described by Lee and Hwang[17]. Soil samples were taken from a depth of 10-20 cm below the soil surface. The soil of the top region (10 cm from the surface) was excluded. Samples were air-dried at room temperature for 7-10 days and then passed through a 0.8 mm mesh sieve and were preserved in polyethylene bags at room temperature before use. Samples (10 g) of air-dried soil were mixed with sterile distilled water (100 mL). The mixtures were shaken vigorously for 1 h and then allowed to settle for 1 h. Portions (1 mL) of soil suspensions (diluted 10-1) were transferred to 9 mL of sterile distilled water and subsequently diluted to 10-2, 10-3, 10-4, 10-5 and 10-6. Inocula consisted of adding aliquots of 10-3 to 10-6 soil dilutions to autoclaved CGA (1 mL-25 mL CGA) at 50°C before pouring the plates and solidification. Three replicates were considered for each dilution. Plates were incubated at 30°C for up to 20 days. From day 7 on, actinomycete colonies were isolated on CGA, incubated at 28°C for one week and stored refrigerated as pure cultures before use. For screening studies 110 pure actinomycete strains were collected.
Screening procedures and in vitro antifungal bioassays Agar disk-method: Each actinomycete isolate was smeared on CGA medium as a single streak and after incubation at 28°C for 4-6 days, from well-grown streaks 6 mm agar disks of actinomycete colony mass was prepared by using sterile cork borers. Disks were then aseptically transferred to PDA plates having fresh lawn cultures of fungal isolates. Controls included using plain disks from CGA medium. Plates were incubated at 25-26°C for 4-6 days and bioactivity was evaluated by measuring the diameter of inhibition zones (DIZ, mm)[12,18].
Dual culture bioassay: Fungal mycelial-disks (diameter of 6 mm) prepared from growing margin of cultures of test fungal isolates and placed in the center of PDA plates and at 30 mm distance from it, the actinomycete disks (prepared as mentioned) were placed. Plates incubated at 27°C for 2-3 weeks [4]. Antifungal activity was evaluated as follows and the ratings used were modified from those of Lee and Hwang[17], El-Tarabily et al.[19]: (1) no inhibition = mycelial growth not different from control (-); (2) weak inhibition = partial inhibition of mycelial growth, measured as a distance of 5-9 mm (+) to actinomycete colonies; (3) moderate inhibition = almost complete inhibition of mycelial growth, measured as a distance of 10-19 mm (+ +); (4) strong inhibition = complete inhibition, in which most mycelia did not grow, measured as a distance of > 20 mm (+ + +) to actinomycete colonies. Controls included fungal mycelial plugs in center of non-actinomycete inoculated PDA plates.
Well diffusion-method: For evaluation of antifungal activity of aqueous samples, by use of sterile cork borer wells (6x4 mm, 2 cm apart) were punctured in fresh fungal lawn-cultures or at 30 mm distance from fungal plugs. Respective concentrations in dimethyl sulfoxide: methanol (1/1: v/v) solvent (DM solvent) were then administered to fullness in each well. Plates were incubated at 26°C for 4-6 days for lawn cultures and 14 days for dual culture disk-plugs. Bioactivity was determined by measuring inhibitory zones (mm). Each experiment was repeated three times and the mean of inhibitory zones recorded. Controls included use of blank wells and use of DM solvent without test compounds[12].
Submerged culture of active strains and monitoring optimum production time of active compounds: Active strains of actinomycete were grown in submerged cultures of CG medium on rotary shakers under 130 rpm at 30°C. To monitor the activity, aseptically small aliquots of culture media were taken every 24 h for 20 days and the activity was evaluated by well diffusion-method[12,18].
Preparation of crude extract: When the activity reached maximum, the cultures were harvested; spores and mycelia were excluded by filtration through two layers of cheese cloth. The clarified sap was then dried to dark crude under reduced air at 50°C and kept refrigerated before use.
Classification of actinomycete: Actinomycete colonies were characterized morphologically and physiologically to the genus level following the direction mentioned in the methods manual of international cooperative project for description and deposition of cultures of Streptomyces (ISP)[20]. Identification procedures were done in the Department of Biological Sciences, University of Science and Technology, Irbid, Jordan[21-23].
Determination of Minimal Inhibitory Concentrations (MIC) and dose response: To measure the MIC values, two-fold serial dilutions of 20, 10, 5, 2.5, 1.25, 0.625 and 0.312 mg mL-1 of the crude extract were prepared in DM solvent and assayed by well diffusion-method as mentioned earlier. The MIC was defined as the lowest concentration able to inhibit any visible fungal growth. All data represent average of three replicated experiments.
Temperature effect on activity: The effect of temperature on bioactivity was determined by exposing small aliquots (10 mg mL-1) of soluble crude to each of 30, 40, 50, 60, 70, 80 and 90°C for 10 min[24]. Bioactivity of treated samples was evaluated using well diffusion method. Control included incubation of an untreated sample at 26°C.
Determination of shelf life or stability of active crude: To measure the stability of the active crude in both soluble and dry states, 5 mg mL-1 of each sample was prepared in DM solvent and 5 mg dry samples placed in small vials. Some of the samples were kept at room temperature and some refrigerated at 4°C and tested using agar diffusion-method for antifungal activity at 14 days intervals as long as the activity persisted.
Effect of culture filtrate on fungal spore-germination: To evaluate of the ability of culture filtrate of strain101 to inhibit conidial germination of V. dahliae isolates, to each well of depression slides, 0.2 mL of the culture filtrate of strain101 was placed and dried at room temperature. Spore suspension (5x103 mL) from each of V. dahliae isolates prepared in distilled water and added over each of the dried culture filtrates separately. Slides incubated in a humid chamber at 26°C for 24 h. Controls included CG medium plus spore suspension. Spore germination of V. dahliae isolates and germ tube characteristics were monitored and compared to the control with a light microscope[12,25].
Investigation for fungicidal and/or fungistatic activity: Small blocks of inhibition zones (1 mm3) of active actinomycete isolates against the phytopathogenic fungi were transferred to fresh PDA plates and incubated for 7 days at 26°C. During incubation, growth or lack of growth of fungi was investigated microscopically. Rejuvenation of growth was indicative of fungistatic and lack of growth represented fungicidal properties of the antagonist.
RESULTS
Screening and bioassays: In screening for actinomycete having antifungal activity, 110 isolates were screened from which 10 isolates showed activity against at least one fungal isolate. Figure 1 represents actinomycetes colonies in preliminary soil cultures from which pure cultures prepared for the tests. Bioassay results on Alternaria solani of actinomycete strains are indicated in Fig. 2.
| Table 1: | Screening results of antifungal activity in some Iranian actinomycete strains measured against nine fungal isolates as determined by well diffusion method |
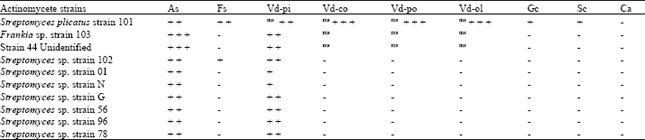 | |
| As: Alternaria solani (Tomato isolate); Fs: Fusarium solani (Potato isolate); Vd-pi: Verticillium dahliae (Pistachio isolate); Vd-co: Verticillium dahliae (Cotton isolate); Vd-po: Verticillium dahliae (Potato isolate); Vd-ol: Verticillium dahliae (Olive isolate); Gc: Geotrichum candidum (Tomato isolate); Sc: Saccharomyces cerevisiae (PTCC No. 5052), Ca: Candida albicans. +: 5-9 mm, weak; + +: 10-19 mm, moderate; +++: > 20 mm, strong and -: no inhibition. ms: Inhibition of micro sclerotia and melanin production in V. dahliae isolates | |
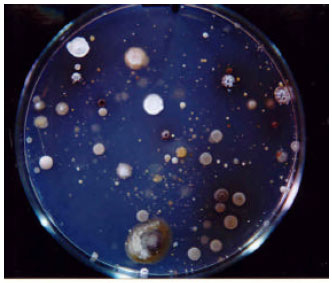 | |
| Fig. 1: | Actinomycetes colonies in preliminary soil culture-plates from which pure cultures prepared for the tests |
Table 1 shows antifungal activity of active actinomycete strains on seven phytopathogenic fungal isolates and two yeasts.
Taxonomy of actinomycete: Broad spectrum strains against phytopathogenic fungi were identified as reported recently, but strain 44 has not been identified yet. Based on the literature reviews, strains 101 and 103 were new records from Iran[22].
Activity versus time in submerged cultures: Production time versus activity for 3 strains is indicated in Fig. 4. In all of the strains, activity reaches its maximum approximately 8-9 days after inoculation. In strain 44 the activity drops sharply to zero after 13-14 days.
MIC and dose response: Activity versus concentration in Streptomyces plicatus strain 101 is indicated in Fig. 3 and 5, the MIC was determined 5 mg mL-1 accordingly.
| Table 2: | Temperature effect on active principles of strain 101 at 10 mg mL-1 against V. dahliae |
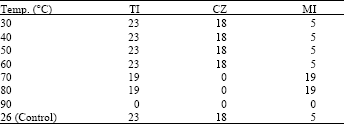 | |
| TI: Total inhibition zone, mm; CZ: Clear inhibition zone, mm; MI: Micro sclerotia inhibition zone, mm | |
| Table 3: | Fungicidal and fungistatic activity of 5 active strains of actinomycete against three phytopathogenic fungi |
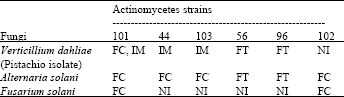 | |
| FC: Fungicidal activity; FT: Fungistatic activity; IM: Inhibition of microsclerotial formation; NI: No inhibition | |
Stability of active crudes: Stability of the active crude in DM solvent and dry form at room temperature (12-30°C) was 2 and 6 months, respectively, assayed by using agar diffusion-method against four Verticillium dahliae isolates. At 4°C, refrigerated crudes remained active for one and half years, the time of this report. Further investigation would be continued as long as the activity persists.
Antifungal activity of submerged cultures: Activity versus post seeding time in submerged media cultures of three strains is indicated in Fig. 4. Since the activity reaches its maximum after 9-10 of post seeding, this time was used to harvest cultures for preparation of crude extracts.
Spore germination: Culture filtrate of strain 101 inhibited spore germination of all four V. dahliae isolates whereas controls showed no inhibition.
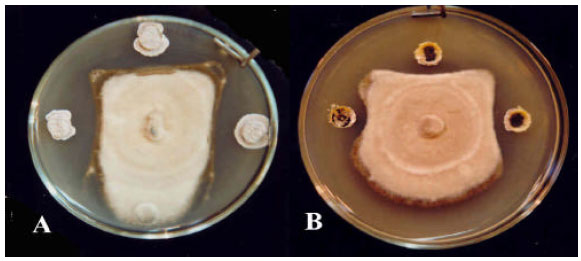 | |
| Fig. 2: | Bioassay results of actinomycete strains against Alternaria solani. A) Clockwise from top strains of 56, G, blank disk (control) and 96 in Agar disk method; B) Clockwise from top strains of 56, G and 96 in Well diffusion method. Inhibition zones are indicative of antagonistic activity of actinomycetes |
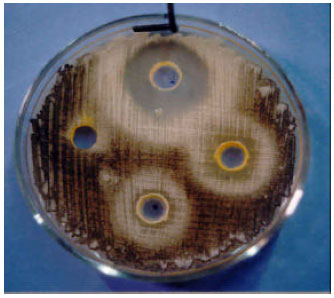 | |
| Fig. 3: | Antifungal activity of crude extract of actinomycete strain 101 measured in Well diffusion method at different concentrations. Clockwise from top 20, 10, 5 and 2.5 mg mL-1 of crude sap administered to each well. As indicative in 3rd and 4th wells, the MIC of crude is 5 mg mL-1 |
Temperature effect: Temperature effect on active principles of strain 101 at 10 mg mL-1 against V. dahliae is indicated in Table 2. As it is indicated, two types of inhibitory reactions behave different. At 70°C no clear zone was observed, apparently the active compound is inactivated but microsclerotial formation persists at least beyond 80°C indicative of presence of another active principle. Total activity disappeared at 90°C.
Fungicidal and fungistatic activity: Fungicidal and fungistatic activity of five active strains of actinomycete against three phytopathogenic fungi are indicated in Table 3.
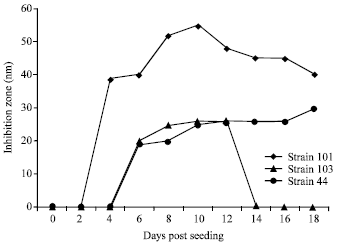 | |
| Fig. 4: | Activity versus post seeding time in submerged media cultures of strains: 101, 44 and 103 against Pistachio root-isolate of Verticillium dahliae monitored by well diffusion-method and monitoring optimum production time of active compounds |
It was determined that clear zones were indicative of fungicidal and the zones bearing only mycelial growth had static effect on microsclerotial formation, hence the latter can be phrased as microsclerostatic activity.
DISCUSSION
Actinomycete, particularly Streptomyces spp. by virtue of their wide distribution, filamentous growth in soil and antibiotic production, may participate actively in establishing the microbiological equilibrium in soil and may be a factor in affecting the incidence of certain soil-borne plant pathogens[26].
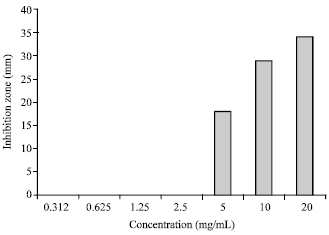 | |
| Fig. 5: | Dose response of Streptomyces plicatus strain 101 crude extract against Pistachio root-isolate of Verticillium dahliae determined by agar diffusion method at 26°C for 7-10 days of inoculation |
One possible approach to biological control of important diseases such as Verticillium and Fusarium wilts, Fusarium rots and Early blight in the fields, greenhouses and orchards in Iran is to inoculate soil with complexes consist of selected antagonists. However, this requires investigation of conditions which favor the survival of the antagonists, because soil is very complex substrate in which numerous factors influence the number of microorganisms as well as the qualitative composition of its microflora. In this study, we attempted to isolate and study a preliminary screening of actinomycete in restricted area of Kerman Province, southeast of Iran. The results may be considered for further studies of actinomycete microflora in native Iranian soils with the goal to find new agents in biocontrol of soil born diseases of plants[27]. The genes encoding many antifungal characteristics are currently being used by agribusiness to create genetically modified plants that have increased fungal resistance in the field. Whether these transgenic plants and the crops derived from them gain acceptance in the marketplace remains to be seen[28]. If the definition of biological control includes the plant induced or genetically modified to defend itself, then biological control has been the most significant approach to plant health management during the 20th century and promises though modern biotechnology to be even more significant in the 21st century. Indeed, the promise of the new tools of biotechnology for crop-based agriculture has arrived, all based on the use of transgenes from microorganisms. Furthermore, the many facts of genomics research, applied to plants and microorganisms, can be expected to reveal entirely new biological approaches to plant health management-approaches likely to make the current use of transgenes for resistance seem crude by comparison[29]. Nearly all private investments in biological control today, at least in the United States, are for transformation of plants to express genes from microorganisms. In these examples, the plant rather than the microorganism becomes the biological control agent[30]. We believe that the results of these findings can form the avenue for production of resistant transgenic-plants with recombinant DNA having antifungal genes cloned from biologically active Streptomyces spp.
ACKNOWLEDGMENTS
Thanks to Management and Planning Organization of Kerman Province, Kerman, Iran for financial support. Thanks to Head of Research Affairs Office of Bahonar University of Kerman for help and support. Helpful information and kind gifts of fungi by Dr. Zare, Dr. Sharifnabi, Mr. Aminaii and Prof. Banihashemi are appreciated. We also would like to thank helpful works of Dr. Saadoun and his coworkers for identification of actinomycetes. This research is dedicated to Mr. A. Afzalipour, the founder of Bahonar University in Kerman.
REFERENCES
- Cohen, Y. and M.D. Coffey, 1986. Systemic fungicides and the control of oomycetes. Ann. Rev. Phytopathol., 24: 311-338.
Direct Link - Fruh, T., P. Chemla, J. Ehrler and S. Farooq, 1996. Natural products as pesticides: Two examples of strereoselective synthesis. Pestic. Sci., 46: 37-47.
Direct Link - Knight, S.C., V.M. Anthony, A.M. Brady, A.J. Greenland and S.P. Heaney et al., 1997. Rationale and perspectives on the development of fungicides. Ann. Rev. Phytopathol., 35: 349-372.
CrossRefDirect Link - El-Tarabily, K.A., G.E.J. St. Hardy, K. Sivasithamparam, A.M. Hossein and D.I. Kurtboke, 1997. The potential for biological control of cavity-spot disease of carrots, caused by Pythium coloratum by streptomycete and non-streptomycete actinomycetes. New Phytol., 137: 495-507.
Direct Link - Goshi, K., T. Uchida, A. Lezhava, M. Yamasaki, K. Hiratsu, H. Shinkawa and H. Kinashi, 2002. Cloning and analysis of the telomere and terminal inverted repeat of the linear chromosome of Streptomyces griseus. J. Bacteriol., 184: 3411-3415.
Direct Link - Brown, M.E., 1974. Seed and root bacterization. Annu. Rev. Phytopathol., 12: 181-197.
CrossRefDirect Link - Lee, J.Y. and B.K. Hwang, 2002. Diversity of antifungal actinomycetes in various vegetative soils of Korea. Can. J. Microbiol., 48: 407-417.
Direct Link - El-Tarabily, K.A., M.H. Soliman, A.H. Nassar, H.A. Al-Hassani, K. Sivasithamparam, F. McKenna and G.E. Hardy, 2000. Biological control of Sclerotinia minor using a chitinolytic bacterium and actinomycetes. Plant Pathol., 49: 573-583.
Direct Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Shahidi-Bonjar, G.H., M.H. Fooladi, M.J. Mahdavi and A. Shahghasi, 2004. Broadspectrim, a novel antibacterial from Streptomyces sp. Biotechnology, 3: 126-130.
CrossRefDirect Link - Aghighi, S., G.H.S. Bonjar and I. Saadoun, 2004. First report of antifungal properties of a new strain of Streptomyces plicatus (Strain 101) against four Iranian phytopathogenic isolates of Verticillium dahliae, a new horizon in biocontrol agents. Biotechnology, 3: 90-97.
CrossRefDirect Link - Nawani, N.N. and B.P. Kapadnis, 2004. Production dynamics and characterization of the chitinolytic system of the Streptomyces sp. NK1057, a well equipped chitin degrader. World J. Microbiol. Biotechnol., 20: 487-494.
CrossRefDirect Link - Moreno, A.B., A.M. Pozo, M. Borja and B.S. Segundo, 2003. Activity of antifungal protein from Aspergillus giganteus against Botrytis cinerea. Phytopathology, 93: 1344-1352.
Direct Link - Waksman, S.A., 1967. The Actinomycetes: A Summary of Current Knowledge. Roland Press Co., New York, USA., Pages: 286.
Direct Link - Selitrennikoff, C.P., 2001. Antifungal proteins. Applied Environ. Microbiol., 67: 2883-2894.
CrossRefPubMedDirect Link - Cook, R.J., 2000. Advances in plant health management in 20th century. Annu. Rev. Phytopathol., 38: 95-116.
CrossRefDirect Link - Mathre, D.E., R.J. Cook and N.W. Callan, 1999. From discovery to use: Traversing the world of commercializing biocontrol agents for plant disease control. Plant Dis., 83: 972-983.
Direct Link