
Nyoman Pugeg Aryantha
Department of Biology and Research Development Group on Life Sciences, Institut Teknologi Bandung, Jalan Ganesha 10, Bandung 40132, Indonesia
David I. Guest
School of Botany, The University of Melbourne, Victoria-Australia
Plant Pathology Journal
Year: 2004 | Volume: 3 | Issue: 1 | Page No.: 19-25







ABSTRACT
The present study shows that Potassium phosphonate has been proven to slow down the growth rate of P. cinnamomi in in vitro. Phosphonate drench as low as 1 g L-1 was effective in protecting Thryptomene calycina, Banksia grandis and B. spinulosa in pot and field trials. In glass house trials, concentrations as low as 1 g L-1 (drench) significantly suppressed the P. cinnamomi population. Concentrations over 2 1/2 g L-1 were phytotoxic to all plant species tested. The most sensitive species was B. spinulosa. Phosphonate (5 g L-1 ) killed all B. spinulosa plants in seven weeks, therefore it must be used with a great care. Phosphonate treatment alone was effective protecting plants from disease in the field, but did not result in high plant health. Despite new root growth in pot trials after seven weeks, poor growth was commonly observed on T. calycina after 14 months in field trials. This suggests that phosphonate is not suitable as sole application particularly for the long term. A combination of phosphonate with compost as well as antagonist as an integrated management will be a good alternative for P. cinnamomi management in the future.
PDF Abstract XML References
How to cite this article
Nyoman Pugeg Aryantha and David I. Guest, 2004. Phosphonate (PO3-) Effectiveness Against Phytophthora
cinnamomi Rands on Thryptomene calycina, Banksia grandis and
Banksia spinulosa. Plant Pathology Journal, 3: 19-25.
DOI: 10.3923/ppj.2004.19.25
URL: https://scialert.net/abstract/?doi=ppj.2004.19.25
DOI: 10.3923/ppj.2004.19.25
URL: https://scialert.net/abstract/?doi=ppj.2004.19.25
INTRODUCTION
Phosphonates are the fourth major group of systemic fungicides which have good activities against plant diseases caused by Peronosporales and Pythiales[1]. These fungicides include a range of salts (Na+, Mg++, Cu++, Zn++, Al+++) of phosphorus acid and its esters. More recently, a range of formulations based on mono, di-potassium phosphonate, have become available[2].
Whether the chemical works directly to the fungus or not, there are several arguments about mode of action of it in controlling plant disease. Coffey and Joseph[3] found that phosphorous acid (4.1 to 6.2 μg ml-1) inhibited the mycelial growth of P. cinnamomi in in vitro studies. Fenn and Coffey[4] suggested that phosphate metabolism may be one target of phosphonate toxicity in Oomycetes. On the other hand, Phosphonates were shown to enhance plant defense responses, including lignification, phytoalexin accumulation and hypersensitive cell death[5,6].
Several trials on different plants have been reported to give a good control of P. cinnamomi such as: on Azaleas Benson[7]; on avocado Coffey et al.[8]; on Leucodendron Marks and Smith[9] and on Xanthorrhoea minor and X. australis Ali and Guest[10]. In addition, Cooke and Little[11] found a significant control of phosphonate against Phytophthora infestans on potato. In natural ecosystem, phosphonate also reported has been successfully protecting native plants from P. cinnamomi in Western Australian forest[12].
In spite of that, there is no study on the use of phosphonates for controlling P. cinnamomi on phosphorus susceptible species of plants. Therefore, this study was investigating the effectiveness of phosphonate in controlling P. cinnamomi on three Australian native plants i.e. Thryptomene calycina, Banksia grandis and B. spinulosa.
MATERIALS AND METHODS
In vitro assay: In vitro trial on the growth inhibition of P. cinnamomi by potassium phosphonate was carried out on potato dextrose agar (PDA) plates. The final concentration of filter sterilized potassium phosphonate incorporated in the medium were : 1, 2.5 and 5 μg mL-1 after sterilization separately. Plates were made by dispensing 15 mL melted PDA into ten centimeters plastic petri dishes. After the plate cool at room temperature inoculation with five day old P. cinnamomi grown on PDA was conducted. Ten replicates were made for each treatment dose. The plates were incubated at 20oC, the mycelial growth was observed and the diameter of colony was measured after five days.
Glasshouse trial
Pot trial with Thryptomene calycina: Two ages of T. calycina were used in this trial i.e one year and three month-old. River sand was used as potting mix after being sieved through three millimetre sieve, then washed under running tap water for 15 min. The potting mix was inoculated with 10% sand-bran inoculum of P. cinnamomi before being transplanted with T. calycina.
Phosphonate was applied either as a soil drench or a foliar spray. Soil drenches were applied by dipping the base of each inoculated pot for few seconds in a bucket filled to two centimeter depth with a stock solution of potassium phosphonate. A piece of nylon mesh was put inside the base of the pot to trap the sand in the pot. Control pots were dipped the same way except in deionised water. Foliar sprays were applied using 500 mL plastic hand sprayer to spray until the whole plant became wet. Care was taken to minimize the leaking of the solution from the stem into the potting mix.
Five replicates were used for the one year-old plants, while seven replicates were used in the three months-old plant. Observations on the first trial were carried out by counting the total number of dead branches weekly for seven weeks. In the second trial, plant deaths were counted after seven weeks.
P. cinnamomi populations were estimated by using plate counting method with selective medium (PCH agar) as described by Shew and Benson[13]. Each was done after four weeks following transplanting. The dead pants were tested for the presence of P. cinnamomi by plating the root pieces onto PCH medium.
Pot trials with Banksia grandis and Banksia spinulosa: A similar trial was also conducted using B. grandis and B. spinulosa. Due to the limited number of seedlings, plants in this trial were only treated with soil drenches. Five replicates were used for both Banksia species. Observations were conducted for seven weeks by counting the number of dead plants weekly. To assure the plant was killed by P. cinnamomi, its presence was confirmed by direct isolation onto PCH.
Statistical analysis: In vitro inhibition results and total number of dead branches were analysed using a one way analysis of variance (anova) which is provided in MINITAB 11 computer software (Minitab Inc., Pennsylvania, USA). Total deaths of whole plants were analysed by using logistic regression analysis (MINITAB 11).
RESULTS
In vitro assay: All three concentrations of potassium phosphonate (1, 2.5 and 5 g L-1) significantly inhibited the mycelial growth of P. cinnamomi (Table 1). These data were recorded on the fifth day after inoculation of the agar medium. The treatments did not inhibit fungal growth completely. The highest rate (5 g L-1) inhibited colony diameter by 48%. The fungus kept growing but was slower than the non-treated control. Phosphonate affects colony morphology; colonies growing on phosphonate -amended media are more sparse than colonies on unamended agar.
Glasshouse trials
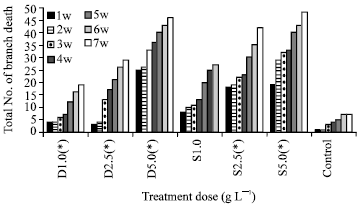
Pot trial of T. calycina: Potassium phosphonate (1 and 2.5 g L-1 soil drench or 2.5 g L-1 foliar spray) significantly reduced plant die back in one year-old T. calycina on the first trial (Fig. 1). This figure shows that both drenches and sprays delay the onset of symptoms and reduce their rate and frail severity.
With three month-old T. calycina in the second trial, drench technique (1 g L-1) and spray technique (5 g L-1) significantly protected the seedlings against P. cinnamomi as estimated by counting total number of plant deaths out of seven replicates (Table 2). An opposite pattern was observed in this trial, i.e deaths are reduced by drenches of 1, but not 2.5 and 5 g L-1. On the other hand, sprays of 5 g L-1 reduced deaths, while 1 and 2.5 g L-1 did not.
Surviving plants treated with soil drenches of 1 g L-1 from this trial produced new and healthy white coloured roots growing back after the first infection by P. cinnamomi (Fig. 2). This kind of root regeneration was very distinctive and was not found on the uninfected plants. Non-infected roots were smaller and light brown in colour. Infected root systems were mostly dead and roots were dark brown to black and rotten in the absence of phosphonate. All dead plants were positive with P. cinnamomi after plating the root pieces on PCH medium.
Quantitative data of P. cinnamomi population from two investigations are presented on Fig. 3. Both results show that potassium phosphonate significantly reduced P. cinnamomi populations over four weeks. All treatments except a spray of 1 g L-1 caused significantly lower P. cinnamomi populations.
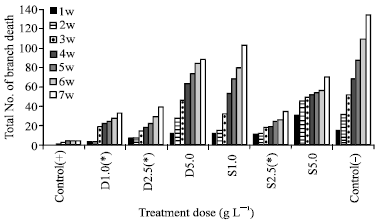 | |
| Fig. 1: | Potassium phosphonate effect on 1 year-old T. calycina over 7-weeks infected with P. cinnamomi (D = Drench; S = Spray; w = week) [Stars (*) indicate a significantly different result from the control α =0.05] |
| Table 1: | In vitro inhibition assay of potassium phosphonate on P. cinnamomi growth after 5 days |
 | |
| (Different letters after the data indicate significancy at α =0.05) | |
| Table 2: | Potassium phosphonate pot trial against P. cinnamomi in 3 months-old T. calycina over 7 weeks |
 | |
| (Letter after the data indicate significancy at α =0.05) | |
| Table 3: | Pot trial of potassium phosphonate against the disease in B. grandis and B. spinulosa over 7-weeks (Plants were infected with P. cinnamomi) |
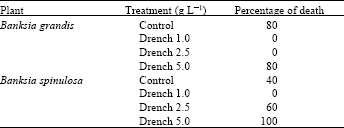 | |
| Table 4: | Phytotoxicity of potassium phosphonate in B. grandis and B. spinulosa over 7-weeks (Plants were not infected with P. cinnamomi) |
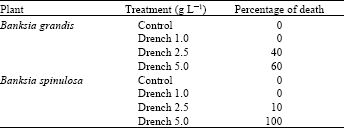 | |
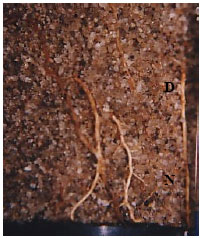 | |
| Fig. 2: | Root growing effect of potassium phosphonate (1 g L-1) on 3 months-old T. calycina after 7 weeks infection. (D) dead root; (N) new root |
As can be seen in this figure, drenches suppress pathogen population more effectively than sprays. The higher the concentration of phosphonate drench, the stronger the suppression of the pathogen population.
Despite positive results against root rot, phosphonate also causes branch and plant death in uninoculated controls. Phytotoxicity effects were observed at a concentrations of 2.5 g L-1 as foliar sprays and 5 g L-1 as soil drenches (Fig. 4). Spraying seemed to affect the plant more than drenching; more die back was found on sprayed plants compared with drenched plants at the same dose.
Phytotoxicity symptoms were observed mainly as burning of the leaves, sometimes even the entire branch. Stronger symptoms were observed as extensive defoliation. On diseased plants, the colour of the leaf slowly turns dull, somewhat pale and becomes desiccated. In contrast, phytotoxicity symptoms appeared suddenly and appeared as leaf burn and extensive defoliation followed by plant death.
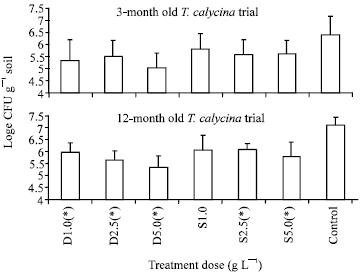 | |
| Fig. 3: | P. cinnamomi populations in phosphonate treated soil on two different repetitions of T. calycina pot trial, each assessed four weeks after inoculation (D = Drench; S = Spray). (Stars (*) indicate statistically different from the control, α =0.05) |
 | |
| Fig. 4: | Phytotoxicity effect of potassium phosphonate on 1 year-old uninfected T. calycina after 7-weeks (D = Drench; S = Spray; w = week) [Stars (*) indicate statistically different from the control, α=0.05] |
 | |
| Fig. 5: | Phytotoxicity of potassium phosphonate on B. spinulosa. Early symptom indicated by arrows at the edge of the leaf (left) and late symptom all leaves are yellow (right) |
Pot trial with B. grandis and B. spinulosa: Treatments with 1 and 2.5 g L-1 gave the best result in protecting B. grandis from P. cinnamomi (Table 3). None of the plants died after application of these two different concentrations of phosphonate over seven weeks. On the other hand, four plants out of five died on the control and 5 g L-1 dose of phosphonate. Doses of 1 g L-1 gave the best protection to B. spinulosa compared to the control and the other concentrations. Doses of 2.5 g L-1 seemed to be less effective than 1 g L-1, as there were three plants that died at this level of phosphonate.
B. spinulosa is more sensitive to phosphonate than B. grandis, but more resistant to P. cinnamomi. Five plants out of five (100%) died at 5 g L-1 phosphonate concentration over seven weeks. On the other hand, B. grandis is less sensitive to phosphonate, as only three plants out of five died at the same level of phosphonate. More plants died at the same concentration for B. grandis due to combination of disease and phytotoxicity. Comparison of these results reveal that most plant death on pots treated with 5.0 g L-1 soil drench were due to phytotoxicity rather than root rot.
Death caused by phytotoxicity on non-disease infected both Banksia species were very obvious as can be seen on the data (Table 4). The phytotoxity symptoms mostly started from the edge or tip of the leaf spreading to the inner part of the lamina until the entire leaf and plant became burnt and died (Fig. 5). The difference between disease and phytotoxic symptom is clearer at later stages. At this stage, it is easy to distinguish plant death due to burning (phytotoxic) or from desiccation (disease). Plants killed by burning turned rusty brown in colour, while plants killed by disease were pale green. Disease symptoms on B. grandis also include a blackening of the stem.
DISCUSSION
In an in vitro study; potassium phosphonate has been shown to slow down the growth rate of P. cinnamomi. The same effect was also found on P. palmivora by Grant et al.[14] and on P. cryptogea and P. capsici[15]. Coffey and Joseph[3] also found that 4.1 to 6.2 μg mL-1 phosphorus acid could inhibit 50% of mycelial growth of P. cinnamomi. A similar level on growth inhibition of P. cinnamomi was found at a concentration of 5.0 μg mL-1. In addition, Coffey & Bower[16] also discovered that P. cinnamomi together with P. citricola and P. citrophthora were the most sensitive species to phosphorous acid among nine Phytophthora spp. studied. It does not kill the fungus as most chemical agents do. It seems that the physiology of the fungus has been altered, as can be noticed from the mycelial growth rate and the colony morphology. Grant et al.[14] found that a low concentration of phosphonate could alter the metabolism of P. palmivora; reducing the amount of macromolecular materials of the fungus.
In the glass house study, phosphonate reduced the population of P. cinnamomi at concentrations as low as 1 g L-1 (drench) or 2.5 g L-1 (spray). It is interesting that the population of P. cinnamomi is also reduced by foliar sprays. This perhaps supports the complex mode of action of this fungicide as been proposed by Guest and Bompeix[17]. It seems that the lower population of P. cinnamomi compared with the control is due to plant reaction. Perhaps the plant absorbed the phosphonate through leaves and stems, then used and released some metabolic product as a defense mechanism. This metabolic product may inhibit the formation of reproductive structure such as sporangium or may also abort the germination of chlamydospore of P. cinnamomi. Vegh et al.[18] found that fungicidal activity of phosphonate breakdown product (ie H3PO3) may persist in soil for several months. Another possibility is that the remnant of phosphonate on the leaves and stems may be washed into the soil during watering and encounter the fungus directly. This is possible to happen in pots, since the volume of the pot is small. As a result, the run-off of phosphonate does not go any where. This small amount of phosphonate may be enough to inhibit the growth or the formation of reproductive structures of P. cinnamomi. As a comparison, the formation of P. clandestina sporangia were 50% inhibited by as low as 1.4 ppm phosphonate in sterile pond water[19].
The glass house trial showed that a concentration of potassium phosphonate as low as 1 g L-1 effectively protected T. calycina, B. grandis and B. spinulosa from root rot caused by P. cinnamomi. Phosphonate was also found to be effective against P. cinnamomi, P. nicotianae and P. palmivora infecting lupin, tobacco and paw-paw when applied as a drench[20]. Marks and Smith[9] found that phosphonate was most effective against P. cinnamomi on Leucodendron when applied as a prophylactic. The fact that many new, healthy and vigorous roots grew from T. calycina treated with 1 g L-1 drench of phosphonate partly explains the reduced severity of symptoms.
Concentrations as high as 5 g L-1 or above were found to be phytotoxic to all species tested. The most sensitive species was B. spinulosa. Phosphonate killed all five plants at 5 g L-1 and one plant out of five at 2.5 g L-1 concentration in seven weeks. At the latter concentration, all the remaining plants were suffering severe phytotoxic effects; and it would eventually die within a few weeks. Therefore, it can be concluded that 2.5 g L-1 phosphonate is phytotoxic to B. spinulosa and 5 g L-1 is phytotoxic to T. calycina and B. grandis. Ali and Guest[10] found that 5 g L-1 of phosphonate spray was also phytotoxic to Xanthorrhoea australis and X. minor in a glass house study.
Phytotoxicity symptoms were found to be different from disease symptom. In general, phytotoxicity shows as leaf burning while disease symptom typically involve desiccation of the plants. The colour of the leaves in late stages of phytotoxic symptoms were bright rusty brown while in the disease symptoms are pale green. On B. grandis, the disease symptom was also associated with a blackening of the stem base which was not common on the toxicity symptoms. In the early stages of disease symptoms, the leaf changed its colour from green to bright yellow starting from the center of the leaf (from the veins) towards the edge. In phytotoxic symptoms, the spread of the symptoms scattered randomly and there was no gradual change of colour as such; the leaves became burnt in a relatively shorter time.
On B. spinulosa clear phytotoxic symptoms were observed at an early stage. Phytotoxic symptoms included a colour change from green to rusty yellow, starting from the edge of a leaf moving in towards the center part. Stems normally remain alive (yellow greenish), even though all leaves are already burnt and look dead. This did not happen in disease symptoms, where the whole plant wilted, then died.
On Thryptomene, defoliation is the obvious symptom of toxicity, while diseased plants usually retain their leaves. The colour of the basal leaves usually changed from green to yellow as a result of phytotoxicity. Disease symptoms appeared as desiccation of the whole plant, a single leaf or one or many branches, but leaves did not drop as in the toxic symptom.
ACKNOWLEDGMENTS
The authors acknowledge with thanks to the Mount Gibson Trust of Australia for sponsoring this research.
REFERENCES
- Cohen, Y. and M.D. Coffey, 1986. Systemic fungicides and the control of oomycetes. Ann. Rev. Phytopathol., 24: 311-338.
Direct Link - Guest, D.I., K.G. Pegg and A.W. Whiley, 1996. Phytophthora diseases, Part 1-their control. J. Aust. Plants, 18: 366-369.
Direct Link - Fenn, M.E. and M.D. Coffey, 1985. Further evidence for the direct mode of action of fosetyl-Al and phosphorous acid. J. Phytopathol., 75: 1064-1068.
CrossRef - Guest, D.I., 1984. Modification of defense responses in tobacco and capsicum following treatment with Fosetyl-Al [Aluminium tris (o-ethyl phosphonate)]. J. Physiol. Plant Pathol., 25: 125-134.
Direct Link - Benson, D.M., 1990. Landscape survival of fungicide treated Azaleas inoculated with Phytophthora cinnamomi. J. Plant Dis., 74: 635-637.
CrossRef - Coffey, M.D., S.D. Campbell and F.B Guillemet, 1984. Chemical control of Phytophthora cinnamomi on avocado rootstocks. J. Plant Dis., 68: 956-958.
Direct Link - Marks, G.C. and I.W. Smith, 1992. Metalaxyl and phosphonate as prophylactic and curative agents against stem infection of Leucodendron caused by Phytophthora cinnamomi. J. Exp. Agric., 32: 255-259.
CrossRef - Ali, Z. and D.I. Guest, 1998. Potassium phosphonate controls Phytophthora cinnamomi root rot of Xanthorrhoea australis and X. minor. J. Aust. Plant Pathol., 26: 40-44.
CrossRef - Cooke, L.R. and G. Little, 2002. The effect of foliar application of phosphonate formulations on the susceptibility of potato tubers to late blight. Pest Manage. Sci., 58: 17-25.
Direct Link - Shew, H.D. and D.M. Benson., 1982. Qualitative and quantitative soil assays for Phytophthora cinnamomi. Phytopathology, 72: 1029-1032.
Direct Link - Grant, B.R., R.H. Dunstan, J.M. Griffith, J.O. Niere and R.H. Smillie, 1990. The mechanism of phosphonic (phosphorous) acid action in Phytophthora. Aust. Plant Pathol., 19: 115-121.
CrossRefDirect Link - Perez, V., A.M. Mamdouh, J.C. Huet, J.C. Pernollet and G. Bompeix, 1995. Enhanced secretion of elicitins by Phytophthora fungi exposed to phosphonate. Cryptogamie Mycol., 16: 191-194.
Direct Link - Coffey, M.D. and L.A. Bower, 1984. In vitro variability among isolates of eight Phytophthora species in response to phosphorous acid. J. Phytopathol., 74: 738-742.
CrossRef - Guest, D.I. and G. Bompeix, 1990. The complex mode of action of phosphonates. J. Aust. Plant Pathol., 19: 113-115.
CrossRef - Greenhalgh, F.C., R.F. Deboer, P.R. Merriman, G. Hepworth and P.J. Keane, 1994. Control of Phytophthora root rot irrigated subterranean clover with potassium phosphonate in Victoria, Australia. J. Plant Pathol., 43: 1009-1019.
Direct Link - Smillie, R., B.R. Grant and D. Guest, 1989. The mode of action of phosphite: Evidence for both direct and indirect modes of action on three Phytophthora spp. in plants. J. Phytopathol., 79: 921-926.
Direct Link