
Mohamed A. Rizk
Department of Botany, Faculty of Science, University of Cairo, Giza 12613, Egypt
Tarek A. A. Moussa
Department of Botany, Faculty of Science, University of Cairo, Giza 12613, Egypt
Plant Pathology Journal
Year: 2003 | Volume: 2 | Issue: 1 | Page No.: 28-38







ABSTRACT
Sugarbeet seeds of cultivars Raspoly, Kawemira and Farida were subjected to different gamma irradiation doses (0.0, 0.5, 1.0, 2.0 and 4 kGy) and the response of these cultivars to infection by Rhizoctonia solani, Sclerotium rolfsii and Fusarium oxysporum was studied. The lower doses of gamma irradiation (0.5 and 1.0 kGy) significantly increased the emergence, plant length and dry weight of sugarbeet seedlings in the three cultivars but the higher doses (2.0 and 4.0 kGy) have adverse effect on the emergence of seedlings. The infection rate was significantly decreased with increased the gamma irradiation doses. The TRV was significantly increased in the three sugarbeet cultivars with increased the gamma doses, while the DRV was significantly decreased with increased gamma doses.
PDF Abstract XML References
How to cite this article
Mohamed A. Rizk and Tarek A. A. Moussa, 2003. Impact of Gamma Irradiation Stresses I. Response of Gamma-irradiated Sugarbeet Seeds to Infection by Soil-borne Fungal Pathogens. Plant Pathology Journal, 2: 28-38.
DOI: 10.3923/ppj.2003.28.38
URL: https://scialert.net/abstract/?doi=ppj.2003.28.38
DOI: 10.3923/ppj.2003.28.38
URL: https://scialert.net/abstract/?doi=ppj.2003.28.38
INTRODUCTION
Research on the effects of ionizing radiation on plant cells and seeds started early in the century, when Guilleminot (1908) exposed pumpkinseed to x-rays and observed retardation of growth and decreased viability of the plants. Osborne et al. (1963) found that when pre-irradiation humidity level increased from 30 to 100%, seedling growth of Brassica napus L. decreased. Woodstock and Justice (1967) observed that a 0.5 kGy treatment of seeds stimulated the growth of radish seedling and 0.8 kGy inhibited growth. Chachin et al. (1972) observed that inhibitory effects of gamma radiation on the germination of radish seeds were reduced by treatment with high concentrations of gibberellic acid.
Selim and El-Banna (2001) concluded that irradiation with low doses of 0.5 to 5.0 kR were the safety doses that can be used as a perfect tool for pea seed preservation and stimulating seed germination, growth and metabolic processes as well as increasing seed yield and the nutritional value of seeds under the Egyptian conditions. On the other side, doses of 10, 15 and 20 kR were inhibitory doses; doses of 25 up to 40 kR were lethal.
Control of plant and human pathogens by irradiation was not studied until relatively recently. Cuero et al. (1986) investigated the influence of gamma irradiation and sodium hypochlorite sterilization on maize seed microflora and germination. The germination of the maize seed was not adversely affected by radiation doses up to 1.2 kGy and the microflora were eliminated. Ramakrishna et al. (1991) compared sodium hypochlorite (12.5, 25, or 50%), mercuric chloride (0.1 or 0.2%), methyl bromide, propylene oxide and gamma irradiation for their effectiveness in killing microorganisms on or within barely seeds. Gamma irradiation at 4 kGy eliminated most Alternaria, Fusarium and Epicoccum spp. but 12 kGy was required to kill Bacillus spp. Germination was improved up to 8 kGy but gradually decreased at higher doses up to 15 kGy.
Among crop plants, cultivated barley (Hordeum vulagare L.) has been extensively used as a model plant for studying the cytogenetical effects of a wide range of mutagens (Gaul, 1965; Veleminksky and Gichner, 1975; Nilan, 1981; Lundqvist and Lundqvist, 1987). Genotypes/varieties within a species may differ regarding the occurrence of mutations. Mutagens induce various kinds of damage such as growth reduction, chromosome breakage, DNA damage and gene mutation; all the organisms may develop repair mechanisms for such damages. The frequency and type of mutations produced depend on chromosome and DNA repair mechanisms. Certain chemicals are known to enhance radiation effects by inhibiting the energy rich enzymatic repair processes and protein synthesis and by retarding the DNA polymerase activity (Jagannath and Bhatia, 1974; Kamra et al., 1975).
Induced mutations in crop plants increase genetic variability. Hence, appropriate methods of mutation induction are of high relevance to meet this requirement efficiently. Efficiency is defined as the frequency of desirable mutation in relation to non-desirable biological alteration (Mikaelsen, 1966). Gamma radiation was found to have a significant effect on the first generation of Sudan grass (Krishna et al., 1983). They discovered a variation in Rhode grass as the level of gamma radiation increased. Gamma radiation treatment on Sudan grass showed some changes in stem, leaf, reproductive organs morphology, plant height and habit. Many of the changes favourably affected the green matter yield and other economical traits (Kirillenko and Golovin, 1987).
Application of physical and chemical mutagens to plants may eventually lead to useful mutant plant lines. A number of chemical and physical mutagens were applied to soybean (Katoh et al., 1992, 1993, 1994, 1995; Baoge et al., 1995; Böhmová and Gajdošová, 1993; Rahman et al., 1994; 1995; Byun et al., 1993, 1995).
Cowpea is one of the important pulse crops in tropical Africa. The seeds are a major source of dietary protein in most developing countries (Duke, 1990). Induced mutation breeding, which has been recognized as a valuable supplement to conventional breeding in crop improvement, has been least applied in grain legumes. For example, only eight out of over 1000 improved mutant varieties of different crops released up to 1989 in over 48 countries were cowpeas (Micke et al., 1990).
In the present study, we investigated the effect of gamma irradiation on the response of sugarbeet seeds to infection by pathogenic fungi. We also measured the sugar content and growth parameters in the developing seedlings of sugarbeet cultivars.
Materials and Methods
Pathogenic fungi, culture conditions, plant material and soil
Rhizoctonia solani (AG 2-2) Kühn, Sclerotium rolfsii Sacc. and Fusarium oxysporum were isolated from diseased sugarbeet roots (El-Abyad et al., 1988) and maintained on the medium composed of (g/l): dextrose, 30; KH2PO4, 1; MgSO4.7H2O, 00.5; KCl, 0.5; KNO3, 2; and 1 ml of each of stock solutions (1 g/l) of FeSO4.7H2O, MnSO4.7H2O, ZnSO4. 7H2O and thiamine; agar, 20 g (Johnson and Curl, 1972).
Seeds of sugarbeet (Beta vulgaris L.) cultivars Raspoly, Kawemira and Farida were obtained from the Agricultural Research Center, Giza, Egypt.
The soil used in this study was obtained from the sugarbeet fields at Kafr-El-Sheikh. The total salinity of the soil was 0.730 mmohse measured by an electric conductivity meter (LF 90), pH 7.9.
Pathogenicity experiment
Pathogenicity of R. solani, S. rolfsii and F. oxysporum to non-irradiated and irradiated sugarbeet Raspoly, Kawemira and Farida was examined in pots (30 cm diameter) in the greenhouse.
Inoculum (2% w/w) for R. solani, S. rolfsii or F. oxysporum (determined in a preliminary experiment) was mixed with the soil in the pots and then additional soil (1 cm in thickness) was added at the surface.
Sugarbeet seeds were irradiated by using Russia 60Co gamma chamber, in National Center for Radiation Research and Technology (NCRRT), Nasr City, Cairo, Egypt. The dose rate was 1 kGy/h. The irradiation doses were 0.5, 1, 3 and 5 kGy. Five days after infesting the soil with fungal inoculum, 10 sugarbeet seeds were sown 0.5 cm deep in each pot. Four replicates were used for each treatment, and all pots were kept in the greenhouse at 27±3°C. Emerged and infected seedlings were assessed 45 days later.
Effect of γ-irradiation of sugarbeet seeds on the growth parameters
The pots were kept in the greenhouse for 45 days. The plants were removed, washed with tap water, then plant length and dry weight were measured.
Effect of γ-irradiation of sugarbeet seeds on carbohydrate analysis of sugarbeet roots
The seedlings at each treatment were uprooted and washed with sterile distilled water. Carbohydrates were extracted from the root tissues according to Nemec et al. (1989). Root tissues were dried at 90°C for 90 min, then at 70°C for 22 h. The tissues were ground and 1.5 g samples were extracted with 76% ethanol for 6 h in a soxhelt apparatus. The ethanol extract was evaporated to near dryness in a rotatory evaporator, diluted with water and then filtered and the solution completed to a known volume with distilled water. The sugar content in the filtrate was estimated by determining the direct reducing value (DRV) and the total reducing value (TRV) using the Nelson-Somogyi solution and Spectronic 20 spectrophotometer as described by Naguib (1964).
Direct reducing value (DRV)
A known aliquot of the filtrate was cleared and neutralized to phenol red. The protein in the samples was precipitated by mixing neutralized samples (5 ml each) with 3 ml basic lead acetate, then filtering the mixture on 3 ml sodium dihydrogen phosphate to which 1 ml of H2SO4 was added. The sample was filtrated again to remove the precipitate. The DRV measured in the samples as described by Naguib (1964) at 700 nm.
Total reducing value (TRV)
A known volume of the previously cleared sample was hydrolyzed with a known volume of 1 N HCl for 30 min (total 0.5 N) to estimate the total reducing value (TRV).
The difference between TRV and DRV gives an estimate of non-reducing sugars. All values obtained were calculated in terms of mg glucose/g dry weight root tissues.
Results and Discussion
Pathogenicity
The emergence rate of healthy seedlings of cvs Raspoly, Kawemira and Farida was significantly decreased at higher gamma radiation doses (1.0, 2.0 and 4.0 kGy) but not reduced at lower gamma dose (0.5 kGy) as compared with control (without radiation nor fungal pathogens, Table 1). The results are in agreement with Woodstock and Justice (1967) who observed that a 0.5 kGy treatment of seeds stimulated the growth of the radish seedlings and 0.8 kGy inhibited growth. In soil infested with pathogens, the emergence rate of seedlings was significantly increased in lower gamma doses (0.5 and 1.0 kGy) but significantly decreased in the higher gamma doses (2.0 and 4.0 kGy). The infection percentage in sugarbeet seedlings was significantly decreased in all sugarbeet cultivars infected by the three fungal pathogens R. solani, S. rolfsii and F. oxysporum, with increasing the gamma doses (Table 1). The sugarbeet cultivar Farida was the most susceptible cultivar to fungal infection. These results confirm the conclusion of Cuero et al. (1986) who investigated the influence of gamma irradiation and sodium hypochlorite sterilization on maize seed microflora and germination. The germination of the maize grains was not adversely affected by radiation doses up to 1.2 kGy and the microflora were eliminated. Also Ramakrishna et al. (1991) compared sodium hypochlorite (12.5, 25, or 50%), mercuric chloride (0.1 or 0.2%), methyl bromide, propylene oxide and gamma irradiation for their effectiveness in killing microorganisms on or within barely seeds.
| Table 1: | Effects of different γ-ray doses (KG) of the sugarbeet seeds, on the emergence (E%) of three sugarbeet cultivars and infection (I%) by Rhizoctonia solani, Sclerotium rolfsii and Fusarium oxysporum in emerged seedlings 4 weeks after sowing |
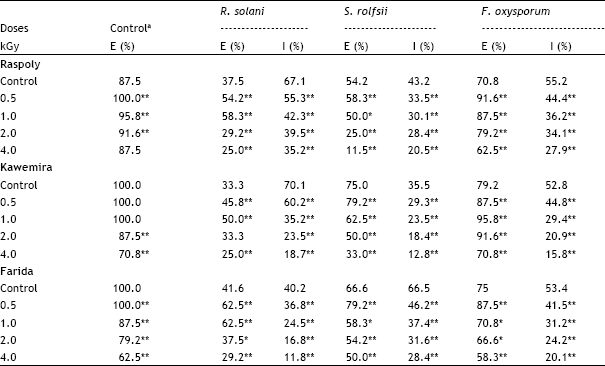 | |
| a Control without pathogen. P <0.05, *significant; P <0.01, **highly significant related to control. | |
Gamma irradiation at 4 kGy eliminated most Alternaria, Fusarium and Epicoccum spp. but 12 kGy was required to kill Bacillus spp. Germination was improved up to 8 kGy but gradually decreased at doses up to 15 kGy. Sitton et al. (1995) irradiated wheat grain infested with teliospores of Tilletia controversa and Tilletia tritici with high-energy electrons (10 Mev). Doses of 4.6-6.7 kGy prevented germination of T. tritici 10.2 kGy was required to eliminate germination of T. controversa. A dose of 1.2 kGy resulted in abnormal germination of the wheat. Hagenmaier and Baker (1997) reduced the normal microflora on commercially prepared fresh-cut lettuce by treating it with a radiation dose of 0.19 kGy. Eight days after irradiation, the unirradiated lettuce had 220,000 cfu/g, while the irradiated lettuce had 290 cfu/g. Hagenmaier and Baker (1998) compared irradiation (0.5 kGy) in an atmosphere of CO2 to chlorination of shredded carrots. Nine days after treatment, the irradiated carrots had a microbial population of 1,300 cfu/g, compared to 87,000 cfu/g on the nonirradiated chlorinated controls.
| Table 2: | Effects of different ã-ray doses (kGy) of the sugarbeet seeds and either Rhizoctonia solani, Sclerotium rolfsii or Fusarium oxysporum, on the length (mm) and dry weight yield (mg) of three sugarbeet cultivars Raspoly, Kawemira and Farida 45 days after sowing |
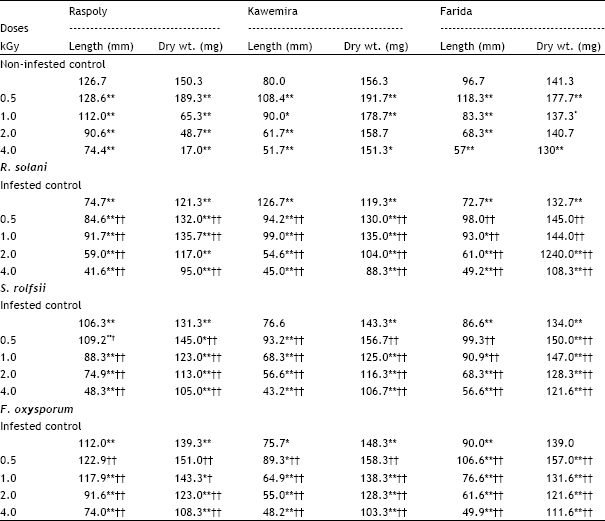 | |
| P <0.05, *significant; P <0.01, **highly significant related to non-infested control. P <0.05, † significant; P <0.01, †H highly significant related to infested control. | |
Growth parameters
The plant shoot height and root depth (plant length) are greatly affected at lower gamma doses (0.5 and 1.0 kGy) the length of plant was significantly increased, while at higher doses (2.0 and 4.0 kGy) it was significantly decreased as compared with non-infested control in the three sugarbeet cultivars (Table 2). In presence of pathogens, the plant length was significantly reduced as compared with non-infested control. At lower gamma radiation doses (0.5 and 1.0 kGy) the plant length was significantly increased, while in higher doses (2.0 and 4.0 kGy) it was significantly decreased as compared with infested control. The plant length of the sugarbeet cultivar Raspoly was significantly decreased in all gamma doses as compared with non-infested control; in cultivars Kawemira and Farida the plant length was significantly increased at 0.5 kGy gamma radiation dose, but was significantly decreased at the rest of doses as compared with non-infested control (Table 2).
| Table 3: | Effect of different ã-ray doses (KGy) of the sugarbeet seeds and either Rhizoctonia solani, Sclerotium rolfsii or Fusarium oxysporum, on the sugar contents (mg glucose g-1 dry weight root tissues) as determined by the total reducing value (TRV) and direct reducing value (DRV) in the roots of three sugarbeet cultivars Raspoly (R), Kawemira (K) and Farida (F) 45 days after sowing |
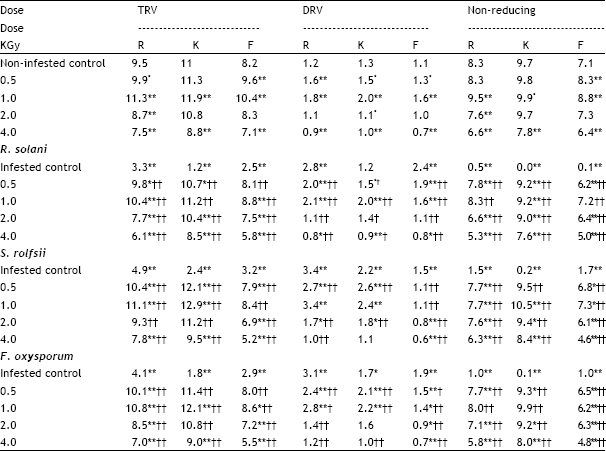 | |
| P <0.05, * significant; P <0.01, **highly significant related to non-infested control. P <0.05, † significant; P <0.01, †Hhighly significant related to infested control. | |
The dry weight yield was significantly decreased in all gamma radiation doses in presence of pathogens in the sugarbeet cultivars Raspoly and Kawemira, but in Farida was increased at 0.5 kGy gamma dose as compared with non-infested control (Table 2). The dry weight yield was significantly increased at lower gamma doses (0.5 and 1.0 kGy) and significantly decreased at higher doses (2.0 and 4.0 kGy) as compared with infested control. In absence of pathogens, the dry weight yields were significantly increased at 0.5 kGy gamma dose and significantly decreased at the rest of gamma doses in cultivar Raspoly, while in cultivars Kawemira and Farida yields were significantly increased at 0.5 and 1.0 kGy gamma doses and decreased at 2.0 and 4.0 kGy gamma doses as compared with non-infested control (Table 2).
Selim and El-Banna (2001) observed that plant height, number of leaves, dry weights of roots and leaves as well as net assimilation rate were significantly increased with increasing irradiation doses up to 2.5 kR, the intermediate dose 5 kR had no significant effect, doses over 10 kR caused a significant decrease in this respect. Relative growth rate, dry weights of shoot and of whole plant, shoot/root ratio, total leaf area, leaf area index and leaf area, were increased significantly when gamma doses applied up to 5 kR. Rajkowski and Thayer (2001) observed that irradiating alfalfa seeds at doses up to 2 kGy did not unacceptably decrease the yield ratio for production of alfalfa sprouts.
Sugar contents
In absence of pathogens, the TRV was significantly increased in lower gamma doses (0.5 and 1.0 kGy) on the other hand, at higher gamma doses (2.0 and 4.0 kGy) the TRV was significantly decreased in all sugarbeet cultivars as compared with control (without radiation nor fungal pathogens, Table 3). The infection of sugarbeet cultivar by any of the three fungal pathogens significantly decreased the TRV in absence of gamma radiation as compared with non-infested control. In all gamma doses the TRV was significantly increased as compared with infested control, while TRV was significantly increased at lower gamma doses (0.5 and 1.0 kGy) but significantly decreased at higher gamma doses (2.0 and 4.0 kGy) as compared with non-infested control.
In absence of fungal pathogens, the DRV was significantly increased at lower gamma doses, but significantly decreased at higher gamma doses, Table 3. In presence of pathogens, the DRV was significantly decreased in all gamma doses in sugarbeet cultivars Raspoly and Farida but significantly increased at lower gamma doses and significantly decreased at higher gamma doses in cultivar Kawemira as compared with infested control. On the other hand, the DRV was significantly increased in lower gamma doses, but significantly decreased in higher gamma doses as compared to non-infested control.
In absence of pathogens, the non-reducing sugar was significantly increased at lower gamma doses and significantly decreased at higher gamma doses as compared with non-infested control (Table 3). On the other hand, the non-reducing sugar was significantly decreased at all gamma doses as compared with non-infested control but significantly increased at all gamma doses as compared with infested control in all sugarbeet cultivars.
Hayashi et al. (1997) observed significant reductions in starch viscosity in brown rice, rough rice and wheat by a radiation dose of 10 kGy from gamma but not electrons, attributing the difference to the very low penetration of the seed by these low-energy electrons. Selim and El-Banna (2001) subjected seeds of pea cv. Little Marvel to different doses of ionizing gamma irradiation (0.0, 0.5, 2.5, 5, 10, 15, 20, 25, 30, 35 and 40 kR) and observed that treating pea seeds with 0.5 kR led to a slight increase in the concentration of non-reducing sugars, other doses inhibited it. Reducing and total soluble sugars at most plant ages, were not affected by dose of 0.5 kR, other doses lowered them.
Acknowledgment
We thank Prof. M. Kassas, Emeritus Professor of Botany at this department for reading the manuscript.
REFERENCES
- Baoge, Z., G. Aigiu, D. Xiangdoug, G. Yuxuan and L. Zixian, 1995. Effect of caffeine or EDTA post-treatment on EMS mutagenesis in soybean. Mutat. Res., 334: 157-159.
CrossRefDirect Link - Byun, M.W., J.H. Kwon and T. Mori, 1993. Improvement of physical properties of soybean by gamma irradiation. Radiat. Phys. Chem., 42: 313-317.
Direct Link - Byun, M.W., I.J. Kang, Y. Hayashi, Y. Matsumura and T. Mori, 1994. Effect of gamma irradiation on soya beans proteins. J. Sci. Food Agric., 66: 55-60.
Direct Link - Chachin, K., H. Honjo and K. Ogata, 1972. Physiological studies on the development of resistance in plant tissues to ionizing radiation. I. Effects of a combination of gamma irradiation and treatment with some protective substances on the germination of seeds and potato tubers. Food Irradiation [Shokuhiu-Shoshal], 7: 6-14.
- Hagenmaier, R.D. and R.A. Baker, 1997. Low-dose irradiation of cut iceberg lettuce in modified atmosphere packaging. J. Agric. Food Chem., 45: 2864-2868.
Direct Link - Hagenmaier, R.D. and R.A. Baker, 1998. Microbial population of shredded carrot in modified atmosphere packaging as related to irradiation treatment. J. Food Sci., 63: 162-164.
Direct Link - Hayashi, T., Y. Takahashi and S. Todoriki, 1997. Low-energy electron effects on the sterility and viscosity of grains. J. Food Sci., 62: 858-860.
Direct Link - Jagannath, D.R. and C.R. Bhatia, 1974. Effect of sodium fluoride on radiation sensitivity of barley seeds. Mutat. Res., 23: 51-56.
Direct Link - Katoh, Y., M. Maekawa and Y. Sano, 1993. Effects of 5-azacytidine on somatic mutation in soybean test system. Mutat. Res., 300: 49-55.
Direct Link - Katoh, Y., M. Maekawa and Y. Sano, 1994. Mutagenic effects of nitropyrenes in a soybean test system. Mutat. Res., 320: 59-68.
Direct Link - Katoh, Y., M. Maekawa and Y. Sano, 1995. Mutagenicity of O-diazoacetyl-L-serine (azaserine) and 6-diazo-5-oxo-L-notleucine (DON) in a soybean test system. Mutat. Res., 342: 37-41.
Direct Link - Nemec, S., D. Phelps and R. Baker, 1989. Effect of dihydrofusarubin and isomarticin from Fusarium solani on carbohydrate status and metabolism of rough lemon seedlings. Phytopathology, 79: 700-705.
Direct Link - Osborne, T.S., A.O. Lunden and M.J. Constantin, 1963. Radiosensitivity of seeds. III. Effects of pre-irradiation humidity and gamma-ray dose on seeds from five botanical families. Radiat. Bot., 3: 19-28.
Direct Link - Rahman, S.M., V. Takagi, K. Kubota, K. Miyamoto and V. Kawakita, 1994. High oleic acid mutant in soybean induced by X-ray irradiation. Biosci. Biotechnol. Biochem., 58: 1070-1072.
Direct Link - Rahman, S.M., V. Takagi, K. Miyamoto and T. Kawakita, 1995. High stearic acid soybean mutant induced by X-ray irradiation. Biosci. Biotechnol. Biochem., 59: 922-933.
Direct Link - Rajkowski, K.T. and D.W.L. Thayer, 2001. Alfalfa seed germination and yield ratio and alfalfa sprout microbial keeping quality following irradiation of seeds and sprouts. J. Food Port., 64: 1988-1995.
Direct Link - Ramakrishna, N., J. Lacey and J.E. Smith, 1991. Effect of surface sterilization, fumigation and γ irradiation on the microflora and germination of barley seeds. Int. J. Food Microbiol., 13: 47-54.
CrossRef - Sitton, J.W., J. Borsa, T.R. Schultz and J.D. Maguire, 1995. Electron beam irradiation effects on wheat quality. seed vigor and viability and pathogenicity of teliospores of tilletia controversa and T. tritici. Plant Dis., 79: 586-589.
Direct Link - Woodstock, L.W. and O.L. Justice, 1967. Radiation-induced changes in respiration of corn, wheat, sorghum and radish seeds during initial stages of germination to subsequent seedling growth. Radiat. Bot., 7: 129-136.
Direct Link - Selim, A.F.H. and E.N.E. Banna, 2001. Ionizing irradiation effects on germination, growth, some physiological and biochemical aspects and yield of pea (Pisum sativum L.) plants. Proceedings of the Umweltverschmutzung in Agypt Folgen fur Mensch Tier und Pflanze Symposium, September 29-October 3, 2001, Cairo, Agypt, pp: 1697-1719.