
Sukamto
Department of Agriculture and Food Technology, Widyagama University of Malang, Indonesia
LiveDNA: 62.28828
M. Sui
Department of Agriculture and Food Technology, Widyagama University of Malang, Indonesia
Sudiyono
Department of Agriculture and Food Technology, Widyagama University of Malang, Indonesia
P. Domas Galih
Department of Agriculture Technology, Brawijaya University, Malang, Indonesia
Fatimah Karim
Department of Culinary Art, State University, Malang, Indonesia
Pakistan Journal of Nutrition
Year: 2019 | Volume: 18 | Issue: 8 | Page No.: 783-790







ABSTRACT
Background and Objective: Black komak seeds (Dolichos lablab) are a potential source of protein. Protein from seeds is thought to contain a protein fraction that is resistant to pepsin. The purpose of this study was to separate the protein fractions that are resistant to pepsin. Materials and Methods: The protein fraction was isolated from komak seed flour. Hydrolysis was performed in vitro; the protein isolates were hydrolyzed using pepsin at 37°C for 30 min in a water bath. A 0.05 M HCl solution was used as a buffer. The ratio between the protein and pepsin was 100:1. The solution was diluted to 5%. Proteins resistant to pepsin were separated and further fractionated into 7S and 11S protein fractions. Results: This study showed that pepsin could only hydrolyze some proteins. The amount of protein resistant to pepsin was 67.93 g/100 g. The 7S protein fraction was dominant compared with the 11S protein fraction. The ratio between the 7S protein fraction and the 11S protein fraction was 18.84: 1.00. Both fractions had different functional properties compared with their protein isolates. Fraction 7S consisted of 3 dominant bands with molecular weights of 41.21, 35.48 and 25.11 kDa. Fraction 11S consisted of 2 dominant bands with molecular weights of 35.48 and 28.18 kDa. Both fractions contained globulin proteins and their functional properties were different from the native protein fractions. Conclusion: This study found that pepsin can be used to separate the protein fraction that is dominated by globulins, specifically the 7S globulin fraction of protein. The 7S globulin fraction has the potential to be developed as a functional food ingredient.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Sukamto, M. Sui, Sudiyono, P. Domas Galih and Fatimah Karim, 2019. Effects of Pepsin on the Separation of the Globulin 7S and 11S Fractions from Lablab Bean (Komak) Seeds (Dolichos lablab) and Its Functional Properties. Pakistan Journal of Nutrition, 18: 783-790.
DOI: 10.3923/pjn.2019.783.790
URL: https://scialert.net/abstract/?doi=pjn.2019.783.790
DOI: 10.3923/pjn.2019.783.790
URL: https://scialert.net/abstract/?doi=pjn.2019.783.790
INTRODUCTION
In general, the globulin fraction is a dominant protein fraction in legume seeds. The percentage of protein can reach 50-90%1, while komak seeds contain 26.86±0.38% protein2. Mature komak seeds contain 26-35% protein3. The globulin fraction is resistant to pepsin4. On the other hand, the globulin protein fraction plays an important role in providing quality food protein5. The globulin protein fraction has functional properties with potential for use in the food industry6. Ramos and Bora1 examined the extraction of the globulin fractions in Brazil nut (Bertholletia excelsa HBK) using 50 mM phosphate buffer containing 0.5 M NaCl (pH 7.2) to dissolve approximately 81% protein, of which 72% were globulins. Venkatachalam et al.4 explained that komak seeds harbor a glycoprotein containing at least 3 polypeptides with a molecular weight of 51-64 kDa. The functional properties of the lablab bean protein, globulin and albumin fractions and lablab bean protein concentrates have been widely studied for development in the food industry. Hrckova et al.7 reported that the use of different protease enzymes to hydrolyze fat-free soy protein can change the functional properties of the foam capacity and the gel properties of proteins. Meinlschmidt et al.8 reported that the allergens and bitter taste of soy protein isolates hydrolyzed using papain can be reduced, while their functional properties can be improved. This study focused on the isolation of the protein fraction of komak seeds from Indonesia that is resistant to pepsin. Proteins were fractionated into 7S and 11S fractions and these fractions were characterized and evaluated for their functional properties. The protein fraction resistant to pepsin is strongly suspected to contain globulin protein.
This study was designed to identify a more effective method for isolating the 7S globulin fraction from the protein fraction using the pepsin. In the future, these protein fractions can be utilized as an ingredient for functional foods to reduce cholesterol, hyperglycemia, obesity and so on.
MATERIALS AND METHODS
Materials: The ingredients in this study were komak seeds from Indonesia. The seeds were optimally mature, had a uniform harvesting age and were free from dirt, pests and diseases.
Preparation of raw flour: Komak seeds were dried at 60°C to reach a moisture content of 12-14%. The seeds were ground into flour with an 80-mesh grain size. The flour was ground again with a plate mill and sifted until the granules escaped from 100 mesh. Fat in flour was separated using hexane. The flour was soaked in a hexane solution at a ratio of 1:5 b/v and stirred with a medium speed mixer for 30 min. Then, the flour was filtered with coarse filter paper and was washed again twice with hexane. The flour was drained at room temperature for 24 h to produce fat-free komak seed flour3.
Preparation of protein isolate: Protein isolation was performed in a manner similar to that descried by Sukamto et al.3 as follows. Lablab bean seed flour was dissolved in a 0.1 M alkaline NaOH solution pH 9.0±0.5 at a ratio of 1:10 b/v and stirred with a low-speed mixer at room temperature for 30 min. Furthermore, the dissolved protein fraction was separated by centrifuging at 3000 rpm for 10 min. The dissolved protein fraction was added to distilled water at a ratio of 1:5 and centrifuged again. Then, the protein fraction was separated again (this step was repeated twice). The dissolved protein fraction was collected in a glass beaker and precipitated at pH 4.0±0.2 using 0.1 M HCl. Protein deposits were separated from the supernatant using Whatman No. 4 filter paper. The precipitate was washed with distilled water thrice. Protein deposits were collected and dried using a frozen dryer.
Isolation of the protein fraction: Isolation of the protein fraction that was not hydrolyzed using pepsin was performed by protein hydrolysis by using pepsin following the Chavan et al.9 method with slight modification, as follows: (1) Pepsin enzyme buffer was prepared from 0.05 M HCl at pH 3.0±0.2, (2) The protein fraction concentration was diluted to 5 g/100 mL using 0.05 M HCl pH 3.0-3.5, (3) Diluted pepsin was mixed with the protein fraction at a ratio of 100:1 (protein: enzyme) and (4)The solution was hydrolyzed at 37°C in a water bath for 30 min. The protein fraction that precipitates is a protein that is not hydrolyzed by pepsin and is strongly suspected to be globulin fraction. The separation of protein deposits that are not hydrolyzed using pepsin was performed as follows: (1) Samples of hydrolysis results were centrifuged at 3500 rpm for 30 min, (2) The supernatant was separated from the sediment, (3) The sediment was collected, added to distilled water at a ratio of 1:5 and centrifuged again (repeated thrice) and (4) Non-hydrolyzed protein fraction after exposure to pepsin was transferred to Whatman No. 4 filter paper and washed with distilled water thrice. Furthermore, proteins that are not hydrolyzed by pepsin were fractionated into fractions 7S and 11S at their respective isoelectric pH points. The 7S fraction was at 4.8 pH, while the 11S fraction was at pH 6.4.
Electrophoresis: Protein fractions 7S and 11S were analyzed using SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel electrophoresis)10. Glycoprotein kits were used to perform the confirmation test of the globulin protein.
Microstructure: The microstructure was observed using TM 3000 Hitachi Scanning Electron Microscope (SEM) manufactured by Hitachi High-Technologies Corporation Tokyo Japan.
Amino acid analysis: Amino acid compositional analysis was performed with the following modified steps4. One hundred milligram of the 7S and 11S fractions were added to 4 mL of 6 N HCl and heated for hours and then cooled. The solution was neutralized with 6 N NaOH to pH 7. (Note that the addition of NaOH is performed drop by drop). The solution was diluted to 10 mL and filtered with an HPLC filter. After derivatization of post-column amino acids, the solution was added to 20 μL of OPA solution. The reactants were then injected to a Shimadzu 10 HPLC Diode Array Detector A. The intensity of the OPA-derivatization of amino acids was monitored at an excitation wavelength of 345 nm and an emission wavelength of 450 nm. The amount of amino acid from chromatogram results was confirmed with PDA table 1/340 nm, 4 nm.
Protein solubility profile as a function of pH (PS): The solubility index (SI) was observed using Sukamto et al.3 method. Briefly, 0.1 g of sample was added to 8 ml of 0.1 N phosphate buffer at pH 2, 3.5, 5, 6.5, 8 and 9.5. This suspension was then stirred with a vortex mixer at maximum speed for 30 min at room temperature. The suspension was then centrifuged at 3000 rpm for 30 min. The precipitate and supernatants were separated. The solubility index (SI) was determined using the following equation11:
![]()
Water holding capacity (WHC): An empty and dry centrifuge tube was weighed (a gram). WHC measurements were performed by inserting 0.5 grams of sample (b gram) into the tube and some water [10-15 times of the sample weight (7.5 mL)] was added. The solution was shaken with a vortex until homogeneous and centrifuged for 5 min at the speed of 3000 rpm. The supernatant was separated from the precipitate. The precipitate that remained in the tube was weighed (c gram). WHC was calculated using the following equation12:
![]()
| a | = | Weight of the empty tube |
| b | = | Sample weight |
| c | = | Weight of water accumulated in the sample |
Oil holding capacity (OHC): An empty and dry centrifuge tube was weighed (a gram). A total of 0.5 g of sample (b gram) was placed into the tube and oil was added at 7× the sample weight (3.5 mL). The tube was shaken with a vortex until the mixture was homogeneous and then centrifuged for 5 min at 2000 rpm. The supernatant was removed and the sediment that remained in the tube was weighed (c gram). Furthermore, the OHC calculation was performed using the following equation12:
![]()
| a | = | Weight of the empty tube |
| b | = | Sample weight |
| c | = | Weight of the oil accumulated in the sample |
Foam capacity and stability: Protein samples were diluted to a concentration of 2% by adding a 0.1 M sodium phosphate buffer, pH was 8.0. The samples were homogenized for 10 min with a mixer at maximum speed. Then, samples were poured into a 25 mL measuring cup and the foam layer volume was measured12.
Foam capacity (FC) is calculated by the following equation:
![]()
Foam stability (FS) is calculated by the equation:
![]()
Emulsion activity index and emulsion stability: Protein samples were weighed. Then, 0.1 g was added to 100 mL of 0.05 M phosphate buffer, pH was 7. The solution was stirred for 15 min. Then, 25 mL of cooking oil was added and the solution was mixed for 3 min. To measure the emulsion power, 1 mL was immediately obtained after blending. Regarding the emulsion stability measurement, 1 mL of the lower emulsion solution was obtained after 5, 10, 15, 20, 60 and 120 min and 24 h. Then, 5 mL of 0.1% SDS was added to each sample and the mixture was vortexed. Then, the absorbance was read at a wavelength of 500 nm13. Furthermore, calculations of the emulsion power and stability of the emulsion were performed using the following formula12:
![]()
| EAI | = | Emulsifying activity index, emulsion activity (m2 g−1) |
| c | = | protein concentration (g mL–1) |
| f | = | volume fraction of oil (mL mL–1) from the emulsion obtained from (the amount of oil volume divided by the total volume of the solution) |
| abs | = | Absorbance |
| Dilution | = | Solution fraction (SDS+emulsion) |
| ESI | = | Emulsifying Stability Index, emulsion stability (h) |
| T | = | Turbidity at 0 h |
| t | = | Difference in the time that will be calculated |
| T | = | Difference in turbidity at 0 h with turbidity at the time that will be calculated |
RESULTS AND DISCUSSION
The results of the protein that was separated but not hydrolyzed using pepsin from komak seeds, on average, was 67.93 g/100 g protein. The resistance of the protein fraction to pepsin was thought to be related to globulin protein folding. Under native conditions, globulins are resistant to pepsin4.
SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel electrophoresis): The SDS-PAGE results of the protein fraction resistant to pepsin are presented in Fig. 1. The protein band (lane 1, Fig. 1) has a molecular weight that varies between 10.182 and 133.35 kDa and the dominant protein bands have a molecular weight between 25.11 and 41.21 kDa. Patil and Benjakul2 reported that the globulin fraction from defatted coconut meat had a molecular weight of 55 kDa. After further fractionation of the protein fraction resistant to pepsin, 2 fractions were obtained, namely, the 7S and 11S fractions. Fraction 7S has several protein subunits that are characterized by major protein bands with molecular weights of 24.88, 34.59 and 43.85 kDa and minor subunits with molecular weights of 10.18, 96.82 and 142.23 kDa.
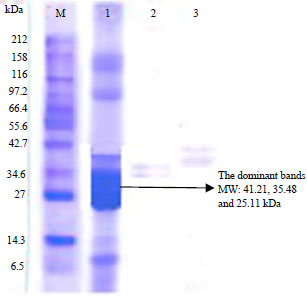 | |
| Fig. 1: | SDS-PAGE of protein fractions which are not hydrolyzed by pepsin (lane 1), supernatant, (lane 2) pepsin, (lane 3) and M (marker protein) enzymes |
Furthermore, the protein bands n the 11S fraction are thinner than the 7S potein fraction bands that are not hydrolyzed by pepsin because the 11S protein fraction is lower than the 7S protein fraction. The ratio of the 7S and 11S protein fractions is 18.84:1.
A confirmation test using a glycoprotein kit is performed to ensure that the 7S and 11S protein fractions resistant to pepsin activity are glycoproteins. The glycoprotein kit confirmation results are presented in Fig. 2. The confirmation test results for glycoproteins in the 7S and 11S protein fractions suggest that the proteins are globulin proteins. Both of these fractions include globulin in the form of a glycoprotein, which is marked by a red protein band (Fig. 2). According to Cai et al.14, the SDS-PAGE results using a glycoprotein dye appear in red. Glycoproteins react with the Periodic Acid Schiff (PAS) reagent, which is a technique used to test for carbohydrates in a protein compound using the Schiff/Glycoprotein Detection Reagent. The glycoprotein levels of the 7S and 11S protein fractions resistant to pepsin are 4.448 and 4.146 mg mL–1, respectively. Venkatachalam et al.4 also reported that globulin is a glycoprotein.
Microstructure of 7S and 11S protein fractions: The structures of the 7S and 11S protein fractions resistant to pepsin under freeze-dried conditions were observed using a scanning electron microscope. The observation results are shown in Fig. 3. Globular proteins interact with each other in both the 7S and 11S protein fractions and form irregular protein aggregates. This interaction occurs as a result of extraction and fractionation processes.
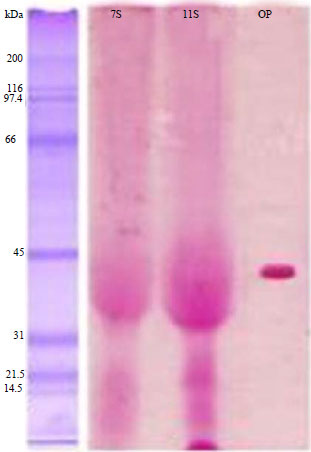 | |
| Fig. 2: | Test results of confirmation of glycoprotein Kit 7S and 11S protein fractions resistant to pepsin |
 | |
| Fig. 3: | Structure of 7S and 11S protein fractions that are resistant to the activity of the pepsin enzyme observed using a 1000X scanning election microscope |
During the extraction and fractionation processes, it is hypothesized that most of the protein fraction undergoes deformation and aggregation to form a protein collection. Crystal fractions of the 7S and 11S globulin fractions were observed at their respective isoelectric points, demonstrating that the polymers themselves easily interact with hydrophobic bonds to form protein aggregates.
Amino acid composition: The amino acid compositions of the 7S and 11S protein fractions are shown in Table 1. The amino acids glycine, alanine and tyrosine were not found in the 11S fraction, whereas serine, histidine and lysine amino acids were not found in the 7S fraction. The quantity of methionine in the 11S protein fraction was considerably higher compared to that in the 7S fraction. While the amount of phenylalanine in the 7S protein fraction was higher than that in the 11S protein fraction. The difference in the number of amino acids between the 7S and 11S fractions of komak seed protein not hydrolyzed by pepsin is thought to be due to differences in the amino acid sequences of each of the polypeptides in the protein subunits.
Functional properties: The results showed that the 7S and 11S protein fractions resistant to pepsin activity had functional properties, as noted in Table 2.
Solubility index (SI): The solubility of the 7S and 11S fractions of proteins that are not hydrolyzed by pepsin is affected by pH. The relationship between pH and solubility of the 7S and 11S fractions not hydrolyzed by pepsin is shown in Fig. 3a and b. The solubility of the 7S and 11S fraction of komak protein not hydrolyzed by pepsin below or above the isoelectric point increased (Fig. 4). The same phenomenon also occurred in several types of protein isolates, including white komak protein isolates12 and beach pea9. Under low pH (below the isoelectric point) and high pH (above the isoelectric point), the interaction between proteins and solvents is facilitated. The difference in the level of solubility between the 7S and 11S fractions is very clear in acidic and alkaline pH.
Water holding capacity: The water holding capacity of the protein fractions not hydrolyzed by pepsin and the 7S and 11S fractions is similar to the WHC from protein isolates, such as peanut at 2.05±0.07 mL g–1 15 and cowpea at 1.38±0.09 mL g–1 16. The ability to absorb and maintain the water protein fraction is influenced by the proportion of hydrophilic groups and hydrophobic groups in the surface of protein molecules12. Solubility is influenced by the ability to absorb water. The greater the solubility, the greater the tendency of WHC to increase.
| Table 1: | Amino acid compositions of the 7S and 11S protein fractions resistant to the pepsin |
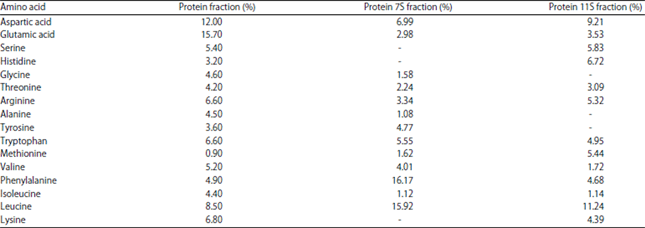 | |
| Table 2: | Functional properties of the protein fractions, 7S protein fractions and lablab bean seed proteins not hydrolyzed by pepsin |
 | |
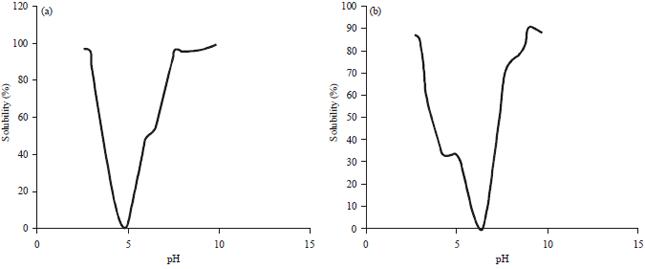 | |
| Fig. 4: | The relationship between pH and the solubility of the 7S (a) and 11S (b) Fractions of the protein resistant to pepsin |
In addition, the WHC value is influenced by polar function groups and non-polar functional groups on the side groups of amino acid residues in the two fractions.
Oil holding capacity: The oil holding capacity averages of the 7S and 11S fractions of protein that were not hydrolyzed by pepsin were greater than the OHC of the protein fraction (Table 2). The OHC values were also higher than those of Brazilian nut globulin isolates (Bertholletia excelsa HBK = 1.4 g g–1) as reported by Ramos1 and isolates of white komak (hyacinth bean) protein = 254±0.2% as reported by Subiago12. The OHC values in the two fractions are related to the number of side groups of non-polar amino acids.
Emulsion activity index and emulsion stability index: The emulsion properties show the ability of proteins to absorb oil and water that enter into the space between molecules during emulsion formation. The EAI values of the 7S and 11S protein fractions not hydrolyzed by pepsin were lower than those obtained from the EAI komak seeds protein fraction (Table 2). Proteins with low solubility exhibit low EAI but high ESI values.
The emulsion stability index: The emulsion stability index values of the 7S and 11S protein fractions not hydrolyzed by pepsin were higher than those of ESI proteins (Table 2). If these values are compared in terms of ESI, soy protein isolates exhibited much lower values. The ESI of soy protein isolates is 52 h13. Differences in the emulsion stability of the 7S and 11S protein fractions are caused by differences in the protein fractions that affect the inter-partial strength, conformational stability and protein solubility. Subiago12 stated that changes in the stability of the emulsion depend on the interferential strength of the material in maintaining the hydrophobic interaction between the oil and the hydrophobic group of proteins in a solution. Emulsion stability is related to solubility and conformational stability. High solubility will produce a high EAI but lead to lower stability.
Foam capacity and stability: The foam capacity of both he 7S and 11S protein fractions not hydrolyzed by pepsin is very high (Table 2). The foam capacity does not follow the stability of foam. The stability of both the 7S and 11S fractions is different from the stability of soy protein isolates. The foam stability of soy protein isolates was 1.66 mL h–1 13. Unstable foam is thought to be caused by the low protein elasticity due to the low number of disulfide bonds in the fraction. Proteins exposed to air with a low elasticity in solution will easily break in the flow of water, thus causing foam collapse.
This study showed that the 7S globulin fraction from komak seeds was successfully isolated based on the nature of its resistance to pepsin. This method can be improved as an alternative assay that is effective in separating protein globulin fractions. The 7S globulin fraction has the potential to be developed as a functional food ingredient that can be used to reduce cholesterol, hyperglycemia and obesity.
CONCLUSION
The protein fraction resistant to pepsin represented 67.93% of the total seed protein isolates. The 7S protein fraction dominates compared with the 11S protein fractions at a ratio of 18.84:1. The 7S fraction consists of 3 subunits characterized by 3 dominant bands with molecular weights of 41.21, 35.48 and 25.11 kDa. The 11S fractions consist of 2 subunits characterized by 2 dominant bands with molecular weights of 35.48 and 28.18 kDa. Both fractions contain globulin proteins. The functional properties of the WHC, OHC and FC from the 7S globulin fractions were better than those of the native protein isolates. The 7S globulin fractions have the potential to be developed as a functional food ingredient.
ACKNOWLEDGMENT
S
Thanks to the Directorate of Research and Community Service, Directorate General of Research and Development Ministry of Research and the Higher Education Republic of Indonesia, which provided financial assistance through national research programs.
REFERENCES
- Ramos, C.M.P. and P.S. Bora, 2003. Extraction and functional characteristics of Brazil nut (Bertholletia excelsa HBK) globulin. Food Sci. Technol. Int., 9: 265-269.
CrossRefDirect Link - Patil, U. and S. Benjakul, 2017. Characteristics of albumin and globulin from coconut meat and their role in emulsion stability without and with proteolysis. Food Hydrocolloids, 69: 220-228.
CrossRefDirect Link - Sukamto, Aulanni'am and Sudiyono, 2009. [Functional properties of the interaction product between globulin of 7S fraction of lablab bean (Dolichos lablab) with Xantan gum]. J. Teknologi Industri Pangan, 20: 117-123, (In Indonesian).
Direct Link - Kozera, W., B. Barczak, T. Knapowski and M. Ralcewicz, 2013. Total and fractional contents of proteins in bean seeds under the conditions of varied fertilisation with microelements. J. Cent. Eur. Agric., 14: 319-327.
CrossRefDirect Link - Chel-Guerrero, L., S. Gallegos-Tintore, A. Martinez-Ayala, A. Castellanos-Ruelas and D. Betancur-Ancona, 2011. Functional properties of proteins from lima bean (Phaseolus lunatus L.) seeds. Food Sci. Technol. Int., 17: 119-126.
CrossRefDirect Link - Hrckova, M., M. Rusnakova and J. Zemanovic, 2002. Enzymatic hydrolysis of defatted soy flour by three different proteases and their effect on the functional properties of resulting protein hydrolysates. Czech J. Food Sci., 20: 7-14.
Direct Link - Meinlschmidt, P., D. Sussmann, U. Schweiggert-Weisz and P. Eisner, 2016. Enzymatic treatment of soy protein isolates: Effects on the potential allergenicity, technofunctionality and sensory properties. Food Sci. Nutr., 4: 11-23.
CrossRefDirect Link - Chavan, U.D., D.B. Mckenzie and F. Shahidi, 2001. Functional properties of protein isolates from beach pea (Lathyrus maritimus L.). Food Chem., 74: 177-187.
CrossRefDirect Link - Soetrisno, U.S. and Z. Holmes, 1992. Functional properties of acid and salt extracted proteins of yellow peas (Pisum sativum L. Miranda). J. Agric. Food Chem., 40: 975-980.
CrossRefDirect Link - Subiago, A., 2006. Characterization of hyacinth bean (Lablab purpureus (L.) sweet) seeds from Indonesia and their protein isolate. Food Chem., 95: 65-70.
CrossRefDirect Link - Webb, M.F., H.A. Naeem and K.A. Schmidt, 2002. Food protein functionality in a liquid system: A comparison of deamidated wheat protein with dairy and soy proteins. J. Food Sci., 67: 2896-2902.
CrossRefDirect Link - Cai, R., A. McCurdy and B.K. Baik, 2002. Textural property of 6 legume curds in relation to their protein constituents. J. Food Sci., 67: 1725-1730.
CrossRefDirect Link - Sibt-e-Abbas, M., M.S. Butt, M.T. Sultan, M.K. Sharif, A.N. Ahmad and R. Batool, 2015. Nutritional and functional properties of protein isolates extracted from defatted peanut flour. Int. Food Res. J., 22: 1533-1537.
Direct Link - Butt, M.S. and R. Batool, 2010. Nutritional and functional properties of some promising legumes protein isolates. Pak. J. Nutr., 9: 373-379.
CrossRefDirect Link