
Ble Celestin Melecony
Center for Research of Oceanology (CRO), Department of Aquaculture, P.O. Box V 18, Abidjan, Cote d’Ivoire
LiveDNA: 225.28791
Yeo Maurice Gopeyue
Laboratory of Biocatalysis and Bioprocesses, Faculty of Food Science and Technology, Nangui Abrogoua University, 02 P.O. Box 801 Abidjan 02, Cote d’Ivoire
Etchian Assoi Olivier
Laboratory of Biology and Animal Cytology, Faculty of Natural Science, Nangui Abrogoua University, 02 P.O. Box 801 Abidjan 02, Cote d’Ivoire
Yao Alla Laurent
Center for Research of Oceanology (CRO), Department of Aquaculture, P.O. Box V 18, Abidjan, Cote d’Ivoire
Dabonne Soumaila
Laboratory of Biocatalysis and Bioprocesses, Faculty of Food Science and Technology, Nangui Abrogoua University, 02 P.O. Box 801 Abidjan 02, Cote d’Ivoire
Pakistan Journal of Nutrition
Year: 2019 | Volume: 18 | Issue: 8 | Page No.: 716-725







ABSTRACT
Background and Objective: In fish feed, fishmeal is the main source of protein that provides energy for fish growth. However, its availability is limited due to the overexploitation of capture fisheries and its price continues to increase each year. The replacement of fishmeal with less expensive energy-rich feed ingredients is necessary for the sustainable development of aquaculture. The objective of this study was to evaluate the effects of different levels of the digestive juice of the snail Archachatina ventricosa in carbohydrate-rich diets on the growth performance and intestinal glycosidase activities of Nile tilapia (Oreochromis niloticus) Materials and Methods: Five experimental diets (R-0, R-1.5, R-2.5, R-5 and R-I) were formulated based on different levels of snail digestive juice (0, 15, 25 and 50 mg g–1). The diet R-I contained 5% inactivated digestive juice. O. niloticus fingerlings with an initial body weight of 22.55±2.29 g reared in glass hatchery tanks were fed with the experimental diets for 56 days. Results: The daily weight gain (DWG), specific growth rate (SGR), feed conversion rate (FCR), protein efficiency ratio (PER), intestinal glycosidase activities and fish body composition were significantly (p<0.05) influenced by the digestive juice. The DWG, SGR, FCR and PER were higher in the fish fed with the diet R-5. The 2.5% digestive juice was shown to have high invertase activity in the intestines of the fish. Conclusion: This study showed that the incorporation of 5% digestive juice from the snail A. ventricosa in experimental diets containing 40% carbohydrates improved growth in juvenile tilapia, O niloticus. The determination of the optimum content of the digestive juice enzyme would better use this ingredient in the formulation of carbohydrate-based fish feeds.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ble Celestin Melecony, Yeo Maurice Gopeyue, Etchian Assoi Olivier, Yao Alla Laurent and Dabonne Soumaila, 2019. Effects of Carbohydrate-Rich Diets Containing Snail Digestive Juice on the Growth Performance and Glycosidase Activities of Nile Tilapia (Oreochromis niloticus). Pakistan Journal of Nutrition, 18: 716-725.
DOI: 10.3923/pjn.2019.716.725
URL: https://scialert.net/abstract/?doi=pjn.2019.716.725
DOI: 10.3923/pjn.2019.716.725
URL: https://scialert.net/abstract/?doi=pjn.2019.716.725
INTRODUCTION
Fishmeal has emerged as the primary protein source in aquafeeds1 and has been estimated to constitute 20-60% of fish diets2,3. However, the availability of this main dietary protein source is limited and its price continues to rise, constraining the long-term sustainable development of aquaculture4,5. Thus, the cost of fish farming production can be reduced by the replacement of fishmeal with cheaper alternative feed ingredients6. The use of carbohydrates in the formulation of aquafeeds would be of great value as a renewable resource for sustainable aquaculture development7.
Tilapias are currently among the most important warm water fish used for aquaculture production8,9. Indeed, these aquatic vertebrates are well-suited to fish farming since these fish are highly prolific and tolerant to a range of environmental conditions. Interestingly, the intensification of aquaculture production systems has resulted in the rapid expansion of tilapia farming and the introduction of these fish into many countries around the world10,11. Among the wide variety of tilapias, the Nile tilapia, (Oreochromis niloticus), is the most cultured species12-14.
In several fish species, it has been observed that the inclusion of carbohydrates in the diets significantly improved the zootechnical performance parameters15. An increase in dietary carbohydrate content improves metabolism and growth in Nile tilapia (O. niloticus)16,17. Unfortunately, the use of carbohydrate-rich diets remains limited due to the low digestibility of carbohydrates by fish18. Thus, the use of exogenous digestive enzymes is an option to improve the nutritive value of carbohydrate-rich diets for cultured fish. Archachatina ventricosa is an abundant snail in Côte d’Ivoire and its digestive juice contains enzymes such as amylases and glucosidases that are able to hydrolyse some substrates, such as starch19,20. This study aimed to evaluate the effects of different levels of A. ventricosa digestive juice in carbohydrate-rich diets on the growth performance and intestinal glycosidase activities of O. niloticus.
MATERIALS AND METHODS
Source of the digestive enzymes: The enzymes were derived from the digestive juice of the snail A. ventricosa. The production of this source of digestive enzymes was previously described by Niamke et al.21. Briefly, specimens of A. ventricosa were fasted for three days to complete digestion. After breaking the shell, the digestive tract was isolated from the visceral mass and the digestive juice was collected. Then, the juice was centrifuged at 6000 rev min–1 to remove all mucus and shell debris. This source of digestive enzymes was stored in a refrigerator at 4°C before use.
Feed ingredients and diets formulation: The protein in the diet was supplied by fishmeal and soybean meal. Maize starch and wheat bran were used as a source of carbohydrates and crude fish oil was used as a lipid source22. The proximate compositions of the main ingredients used in the present study are given in Table 1. The ingredients were ground to a suitable size and mixed in a commercial mixer (Kenwood Chef) for 15 min. Vitamin and mineral mixes were gradually added to the mix. Distilled water (45°C) was slowly added while mixing to achieve a consistency suitable for pellet production. Five experimental diets (R-0, R-1.5, R-2.5, R-5 and R-I) were formulated based on the different levels of the digestive juice (0, 15, 25 and 50 mg g–1). The diet R-I contained 5% digestive juice heated at 100°C for 5 min. After the digestive juice was added, the mixture was incubated at 45°C for 1 hour. After incubation, 3 mm diameter pellets were formed using a kitchen meat grinder (Panasonic MK-G 1800P) and dried for 24 h. All the diets were sealed in plastic bags and stored at 20°C throughout the experiment. The formulation and composition of the experimental diets (% dry weight) are reported in Table 2.
Fish feeding experiment: In this study, fingerlings (22.55±2.29 g) of Nile tilapia, (O. niloticus), were obtained from the aquaculture experimental station at Layo (Dabou, Côte d’Ivoire). One hundred fifty fish were transported to the Oceanological Research Center (CRO, Abidjan) and acclimated to the experimental conditions for two weeks before the feeding experiment began. During the acclimation period, the animals received a commercial diet (35% protein) twice daily (8 am and 4 pm). Then, fish were distributed into fifteen glass aquaria with 10 animals per aquaria and 3 replicate glass aquaria were assigned to each of 5 dietary treatments. The glass aquaria (50 L containing 45 L tap water that had undergone a natural dechlorination of 24 h before adding fish) were in a closed water system23. An electric motor pump ensured a constant flow (1.5 L min–1) of well-aerated water. The water was filtered by setting at 30% daily exchange. The water was completely replaced weekly.
| Table 1: | Proximate composition of the feed ingredients (% dry weight) |
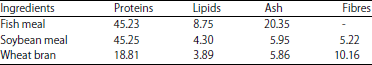 | |
| Table 2: | Formulation and composition of the experimental diets (% dry weight) |
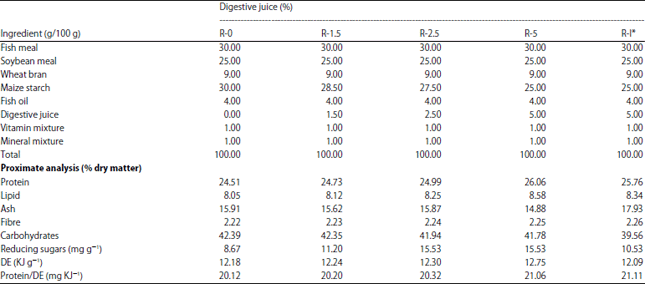 | |
1Per kg premix; Cobalt: 20 mg, Iron: 17 600 mg, Iodine: 2 000 mg, Copper: 1 600 mg, Zinc: 60 000 mg, Manganese: 10 000 mg and Selenium: 40 mg. 2Per kg premix; Vitamin A1: 760 000 IU, Vitamin D3: 880 000 IU, Vitamin E: 22 000 mg, Vitamin B1: 4 400 mg, Vitamin B2: 5 280 mg, Vitamin B6: 4 400 mg, Vitamin B: 1 236 mg, Vitamin C: 151 000 mg, Vitamin K: 4 400 mg, Vitamin P: 35 200 mg, Folic acid: 880 mg, Choline chloride: 220 000 mg and Pantothenic acid: D 14 080 mg. 3Digestible energy: 18,8×protein +37,7×lipid content +11,3×carbohydrate content (Smith50 and Page and Andrews51). *Diet containing 5% digestive juice heated to 100°C for 5 min | |
Every day at 7:30 am, before feeding, the temperature (28.37-28.43°C) and dissolved oxygen (5.89-6.29 mg L–1) were recorded using a multi-parameter (BANTE 900P) and the pH (7.09-7.18) was measured with a pH metre (WTW 330). The total ammonia-N nitrate-N concentration (0.24-0.44 mg L–1) was measured according to Aminot and Chaussepied24.
After the acclimation period, the fish were fed until apparent satiation with the experimental diets twice daily (8 am and 4 pm) for 56 days. To quantify the exact feed intake, the refused feed was siphoned out one hour after feeding, dried and weighed. Once a week, five fish were randomly sampled in each glass aquaria to measure their body weights and the glass aquaria were cleaned. The dead fish were removed from the glass aquaria each day. At the beginning of the experiment, an initial sample of ten fish was taken and frozen (-20°C) for subsequent whole-body proximate analysis. At the end of the experiment, the weight of each fish was recorded. Ten fish from each diet group were stored at -20°C for whole-body composition analysis. Three hours after feeding, five fish from each diet group were dissected and the intestines were collected. These were ground in sodium chloride (9 g L–1) and the homogenate was centrifuged at 6000×g for 30 min. The supernatant was collected in a sample vial and stored at -20°C until use.
Evaluation of the growth performance parameters: The daily weight gain (DWG), feed conversion ratio (FCR), specific growth rate (SGR), protein efficiency ratio (PER), protein gain (PG), lipid gain (LG) and gross energy (GE) metabolism were calculated as follows:
![]()
![]()
| W1 | = | The initial body weight |
| W2 | = | The final body weight |
| t | = | Duration of the experimental period |
![]()
![]()
Protein gain or lipid gain = Final carcass nutrient content-Initial carcass nutrient
GE = 22.2×protein content+38.9×lipid content+17.2×carbohydrate content25
Proximate composition analysis: The proximate composition of the feeds and the fish carcasses were analysed following the AOAC26 methods: crude protein (N×6.25) was determined by the Kjeldahl method after acid digestion; crude lipids were determined by the ether extraction method (Soxtherm, Gerhardt, Germany); dry matter was determined by oven-drying at 105°C for 24 h; the ash and crude fibre were determined by combustion at 550°C in a muffle furnace to a constant weight and by acid/alkali digestion, respectively; the gross energy content was calculated for the fat and protein contents using the equivalents of 38.9 kJ g–1 crude fats, 22.2 kJ g–1 crude proteins and 17.2 kJ g–1 carbohydrates (NFE)25.
Glycosidase analysis: The activities of amylase needed to digest starch and glycogen into glucose and maltose27 and invertase, also called beta-fructofuranosidase or sucrase splitting sucrose into glucose and fructose28, were measured in the intestines of the O. niloticus. The activities of these enzymes were measured by estimating the reducing sugars released after a known period of time29. Briefly, the enzyme preparation was mixed with 1% (w/v) starch or a sucrose solution and a phosphate buffer. The reaction mixture was incubated at 37°C for 30 min. Then, dinitro salicylic acid (DNS) was added to stop the reaction and kept in a boiling water bath for 5 min. After cooling, the reaction mixture was diluted with distilled water and the absorbance was measured at 540 nm. Glucose was used as the standard and amylase activity was expressed as μ mols of glucose released from carbohydrates per min per mg protein (UI or U mg–1 protein). The quantification of the protein in the intestines was measured using bovine serum albumin as the protein standard30.
Statistical analysis: The data were analysed using a one-way analysis of variance (ANOVA) using Statistica 7.1 software. Duncan’s multiple range test was used to compare the differences between the treatment means when significant F-values were observed. All percentage and ratio data were arc-sin transformed before analysis31. The treatment effects were considered to be significant at p<0.05.
RESULTS AND DISCUSSION
RESULTS
Survival rate and weight: The survival rate of the O. niloticus fed the experimental diets (R-0, R-1.5, 2.5, 5 and R-I) for 56 days is illustrated in Fig. 1a.
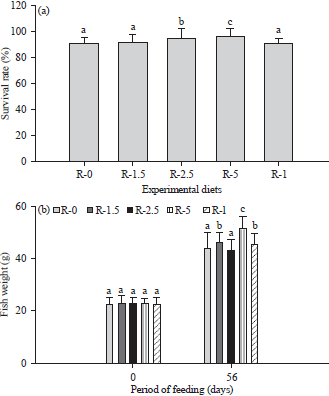 | |
| Fig 1(a-b): | Comparison of the (a) Survival rate and (b) Weight of the tilapia O. niloticus after 56 days of feeding with diets containing different amounts of digestive juice from the snail A. ventricosa |
The data are expressed as the mean±SD. For the weight, different letters denote significant differences (Duncan’s test, p<0.05) between the diet treatments within each feeding period | |
The percentages of the fish that survived were 90.7, 91.3, 94.7, 96.7 and 90.3% in the groups fed with diet R-0, R-1.5, R-2.5, R-5 and R-I, respectively. After 56 days of feeding, the body weights ranged from 43.06-51.29, with the highest value observed in the fish fed with diet R-5 (51.29±4.67 g) (Fig. 1b). The fish fed with diet I containing 5% inactivated digestive juice (heated at 100°C for 5 min) showed a body weight value of 45.6±4.1 g compared to 43.88±5.88 g for the fish fed with 0% digestive juice (control diet). There was a significant increase in the mean survival rate and the body weight between the diet treatments, with the R-5 group having the highest mean survival rate and body weight (p<0.05).
Daily weight gain and specific growth rate: The daily weight gain was significantly influenced by the diet treatments (p<0.05, Fig. 2a), with the highest mean value observed in the R-5 group (0.51±0.07 g day–1). No differences in the daily weight gain were recorded between the R-0 (0.38±0.07 g day–1) and the R-2.6 (0.36±0.04 g day–1) groups and between the R-1.5 (0.41±0.06 g day–1) and the R-I (0.41±0.05 g day–1) groups.
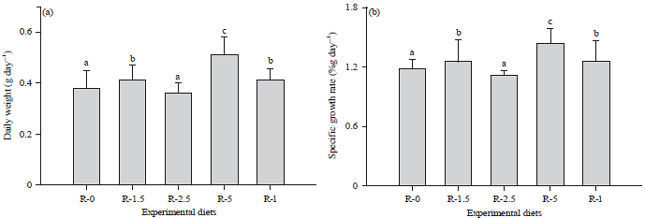 | |
| Fig 2(a-b): | (a) Daily weight gain (g day–1) and (b) Specific growth rate (% g day–1) of O. niloticus after 56 days of feeding with diets containing different levels of digestive juice from the snail (A. ventricosa). Each value represents the Mean±SD Different letters indicate significant differences (Duncan’s test, p<0.05) between the diet treatments |
| Different letters denote significant differences (Duncan’s test, p < 0.05) between the diet treatments | |
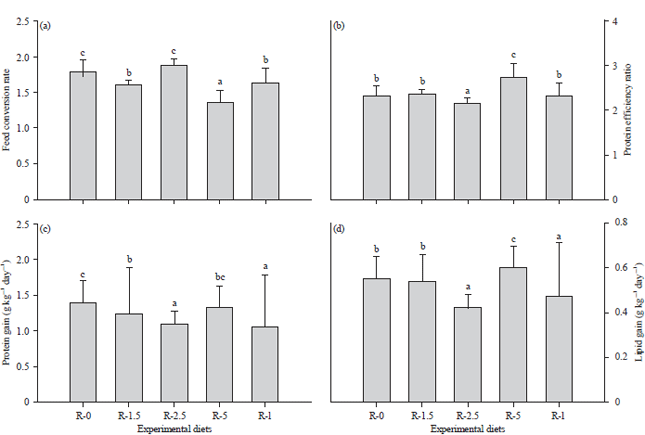 | |
| Fig 3(a-d): | The mean (a) Feed conversion rate, (b) Protein efficiency ratio, (b) Protein gain and (b) Lipid gain in O. niloticus after 56 days of feeding with diets containing different levels of digestive juice collected from the snail A. ventricosa |
| Different letters denote significant differences (Duncan’s test, p < 0.05) between the diet treatments | |
The specific growth rate (Fig. 2b) was significantly influenced by the diet treatments (p<0.05), with the highest value measured in the fish fed with R-5 (1.44±0.14% day–1). The R-0 and R-2.5 groups showed similar responses, which was also the case for the fish fed R-1.5 and R-I.
Feed conversion rate and protein efficiency ratio: The best feed conversion rate corresponding to the lowest value (1.37±0.16) was observed in the fish fed with R-5 (Fig. 3a), compared to the R-2.5 (1.87±0.1), R-0 (1.78±0.17), R-I (1.64±0.2) and R-1.5 (1.60±0.07) diets. With respect to the protein efficiency ratio (Fig. 3b), the fish fed R-5 showed the highest mean protein efficiency ratio (2.73±0.31), while the lowest value (2.15±0.11) was found in the heat inactivated digestive juice group (R-I).
| Table 3: | Body composition of Nile tilapia fed with the different diets (on percentage fresh basis) |
 | |
Each value indicates the mean±standard deviation (Mean±S.D) (n = 3). Different letters indicate significant differences at p<0.05 between the diets. Initial composition of the fish; Moisture: 75.1%, Crude protein: 13.78%, Fat: 4%, Ash: 3.94% and Gross energy: 5.16 kJ g–1 | |
The feed conversion rate and protein efficiency ratio were significantly influenced by the diet treatments (p<0.05).
Protein gain and lipid gain: The protein gain and lipid gain data are shown in Fig. 3c and d, respectively. The protein gain and lipid gain were significantly influenced by the experimental diets (p<0.05). For protein gain, similar responses were observed in the R-2.5 and R-I groups, the R-1.5 and R-5 groups and the R-5 and R-0 groups. For the lipid gain data, the highest value (0.6 ± 0.1 g kg–1 day–1) was observed in the O. niloticus fed the diet R-5, while a similar response was detected in the fish fed with the R-2.5 and R-I diets and for those fed with the R-0 and R-1.5 diets.
Fish body composition: The final body composition of the Nile tilapia at the start and end of the 56 d experimental period is summarized in Table 3. The R-5 group showed the lowest mean moisture value (73.89±0.53), whereas the highest values (approximately 75%) were observed in the fish fed the control diet (R-0) and the other experimental (R-1.5, R-2.5 and R-I) diets. The highest crude protein content value was recorded in the R-5 and R-I groups and the lowest value was obtained from the fish fed the R-0, R-1.5 and R-2.5 diets. The fat content was high in the R-5 group, while the other groups showed low fat levels. A high ash level (4.8%) was recorded in the fish reared with the R-0 and R-2.5 diets compared to the ash content of 3.5% for the other experimental diets. A high gross energy level was recorded in the fish fed the R-5 diet and the lowest value was observed with the control and R-2.5 diets.
Glycosidase activities: The activities of the two glycosidases (UI or U mg–1 protein) measured in the intestines of the O. niloticus fed with the experimental diets are illustrated in Figure 4. Amylase activity was found in all the groups (Fig. 4a), while no invertase activity was detected in the fish fed the R-0 and R-I diets (Fig. 4b).
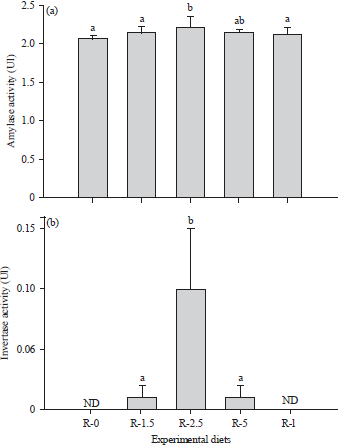 | |
| Fig 4(a-b): | Mean enzyme (amylase and invertase) activities in the intestines of O. niloticus fed diets containing different levels of digestive juice of A. ventricosa |
ND: Not detected. Different letters indicate significant differences (Duncan’s test, p<0.05) between the diets | |
The activity of amylase ranged from 2.07±0.03 U mg–1 protein (R-0) to 2.22±0.13 U mg–1 protein (R-2.5). For invertase, the highest value (0.1±0.05 U mg–1 protein) was recorded in the R-2.5 group and the lowest (0.01± 0.01 U mg–1 protein) was in the R-1.5 and R-5 groups.
DISCUSSION
In the current study, the growth performance of O. niloticus was affected when fish were fed carbohydrate-rich diets supplemented with the digestive juice of the snail A. ventricosa. Overall, the body weight, daily weight gain and specific growth rate increased with the increase in the amount of digestive juice, with the highest growth performance observed in the fish fed diets containing 5% digestive juice (diet R-5). The fish fed diets containing 5% heated digestive juice (R-I) showed a similar growth performance to the fish fed the control diet (R-0). The incorporation of heated digestive juice did not improve the digestibility of the diets. The digestibility of the diet, in other words, the proportion of the nutrients in the feed digested and absorbed by the fish32, is reported to be crucial for growing animals33. The best feed conversion rate (1.37±0.16) (Fig. 3a) and the highest protein efficiency ratio (2.73±0.31) (Fig. 3b) were obtained for the fish fed the R-5 diet. This shows the good digestibility of the diets containing the snail digestive juice. In fact, the digestive juices of snails are recognized to contain most of the carbohydrase enzymes responsible for glucide digestion19,34-36. The results showed that the addition of digestive juice containing the enzymes able to degrade carbohydrates could increase the nutritional value of the diets for fish. As reported by Castillo and Gatlin37, the use of exogenous carbohydrases improved nutrient digestibility by hydrolysing the non-starch polysaccharides from plant feedstuffs. Soybean meal is known to contain a variety of anti-nutritional factors, such as trypsin inhibitors, which have a negative effect on its nutritional value38. In our study, soybean was used as a plant source of proteins in the fish diet formulation (Table 1 and 2). The digestive juice of the snail A. ventricosa, which contains lipases and proteases39,40, could be used to degrade the anti-nutritional factors in the diets of fish to improve growth performance.
In the Nile tilapia, the intestinal amylase activity ranged from 2.07-2.22 U mg–1 protein and showed a high value in the fish fed the R-2.5 and R-5 diets that contained more reducing sugars. These results are consistent with those of Al-Tameemi et al.41, who reported that O. niloticus had the ability to increase their amylase activity with increasing amounts of carbohydrates in their diet. As shown in Table 2, adding the digestive juice of A. ventricosa to diets rich in carbohydrates (42.03±0.22% dry matter) increased the reducing sugars from 8.67 mg g–1 (R-0 diet) to 15.53 mg g–1 (R-2.5 and R-5 diets). When comparing the amylase activity values of different fish species, the large differences observed between species is attributed to their feeding habits42, the nature of the formulated feed43 and the feeding frequency44. Nevertheless, most reports on amylases in fish conclude that omnivorous species such as tilapia fish have higher amylase activities than carnivorous fish41,45. In the majority of studies thus reported, of the two forms of amylase, α-amylase is the most studied in fish species41,42, including tilapia46. The study performed by Fagbenro et al.47 reported the presence of α-amylase in the different regions of the O. niloticus gut, i.e., the stomach (0.951±0.069 mg glucose min–1 mg–1 protein), duodenum (0.758±0.084 glucose min–1 mg–1 protein) and ileum (0.421±0.013 glucose min–1 mg–1 protein). Regarding the second glycosidase, the fish fed the R-2.5 and R-5 diets rich in reducing sugars showed higher invertase activity than the fish reared with the control diet and the other experimental diets. The current findings are in agreement with Fagbenro et al.47, who reported a variety of glycosidases, such as maltase, lactase and cellulase, in O. niloticus, indicating the ability of this fish to digest a variety of carbohydrate food components.
It is noteworthy that the Nile tilapia fed the R-5 diet displayed the highest lipid gain (0.6 g kg–1 day–1) compared to the fish fed the control diet (R-0), while for protein gain, no significant response (1.4 g kg–1 day–1) was noted between the fish fed the R-5 and control (R-0) diets. In fish, growth is recognized as the deposition of protein and lipid48. According to our results, protein gain was more important than lipid gain and one reason is that fish are gaining more protein and less lipid during growth. At the end of the 56 d experimental period, the fish fed with the R-5 diet had the highest body protein content (15.1.±0.03% on a dry matter basis) compared to 14.38±0.03% for the fish reared with the control diet (R-0). The tilapia fed the R-5 diet also showed the highest final body lipid level (5.44±0.03% dry matter) compared to the fish reared with the control diet (4.85±0.1% dry matter). From the beginning, these findings showed that O. niloticus has the ability to utilize carbohydrates for energy, saving protein for deposition and growth. Most commercial feeds today are formulated to increase growth performance by exploiting the protein-sparing effect of high energy carbohydrate diets49.
Findings of the present study agree with earlier study by Yeo et al.23, suggesting the replacement of fish meal protein with non-protein energy sources such as carbohydrates for aquaculture, specifically tilapia culture. The addition of digestive juice containing digestive enzymes such as glycosidase from the snail A. ventricosa to diets rich in carbohydrates increased food digestibility and improved some zootechnical performances in the cultured fish. The present study has indicated that adding 5% digestive juice to diets allows for optimal growth. Since the presence of the digestive juice at 2.5% showed high invertase activity in the intestines of the fish, further study must be performed to determine an optimal proportion of enzymes to add to the diet for better growth optimization. Unlike proteins and lipids, there is no need for carbohydrates in fish. However, one must pay special attention to the use of high carbohydrate foods for tilapia. Indeed, the addition of enzymes leads to a high concentration of simple sugars in the diets. It would be interesting to further investigate whether the use of carbohydrate-rich diets would affect glycaemia regulation in O. niloticus.
CONCLUSION
This study showed that the incorporation of 5% A. ventricosa digestive juice in experimental diets containing 40% carbohydrates improved growth in juvenile tilapia, O niloticus. The determination of the optimum content of the digestive juice enzyme would better use this ingredient in the formulation of carbohydrate-based feeds for fish.
SIGNIFICANCE STATEMENT
This study evaluates the efficiency of snail digestive enzymes in cereal-rich diets on growth performance in juvenile tilapia, Oreochromis niloticus. The results of this research make it possible for food manufacturers and fish farmers to develop cheaper and cost-effective cereal-based foods. This research opens up the prospect of using natural enzymes to improve the feed efficiency of cereals in fish.
ACKNOWLEDGMENT
S
We extend our thanks to the staff at the aquaculture experimental station at Layo for providing the fish and the necessary facilities for the current study.
REFERENCES
- Tacon, A.G.J. and M. Metian, 2008. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture, 285: 146-158.
CrossRefDirect Link - Watanabe, T., 2002. Strategies for further development of aquatic feeds. Fish Sci., 68: 242-252.
Direct Link - Glencross, B.D., M. Booth and G.L. Allan, 2007. A feed is only as good as its ingredients: A review of ingredient evaluation strategies for aquaculture feeds. Aquacult. Nutr., 13: 17-34.
CrossRefDirect Link - Gatlin III, D.M., F.T. Barrows, P. Brown, K. Dabrowski and T.G. Gaylord et al., 2007. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquacult. Res., 38: 551-579.
CrossRefDirect Link - Huntington, T.C. and M.R. Hasan, 2009. Fish as Feed Inputs for Aquaculture-Practices, Sustainability and Implications: A Global Synthesis. In: Fish as Feed Inputs for Aquaculture: Practices, Sustainability and Implications, Hasan, M.R. and M. Halwart (Eds.). Food and Agriculture Organization, Rome, Italy, ISBN: 978-92-5-106419-1, pp: 1-61.
- Otchoumou, A.K., C.M. Ble, O.A. Etchian, Y.L. Alla, S.L. Niamke and J.K. Diopoh, 2012. Effects of increasing dietary protein levels on growth, feed utilization and body composition of Heterobranchus longifilis (Valenciennes, 1840) fingerlings. Afr. J. Biotechnol., 11: 524-529.
Direct Link - Charo-Karisa, H., H. Komen, M.A. Rezk, R.W. Ponzoni, J.A. van Arendonk and H. Bovenhuis, 2006. Heritability estimates and response to selection for growth of Nile tilapia (Oreochromis niloticus) in low-input earthen ponds. Aquaculture, 261: 479-486.
CrossRefDirect Link - Montcho, S.A., H. Agadjihouede, E. Montchowui, P.A. Laleye and J. Moreau, 2015. Population parameters of Oreochromis niloticus (Cichlidae) recently introduced in Lake Toho (Benin, West Africa). Int. J. Fish. Aquat. Stud., 2: 141-145.
Direct Link - Gomez-Marquez, J.L., B. Pena-Mendoza, I.H. Salgado-Ugarte and J.L. Arredondo-Figueroa, 2008. Age and growth of the tilapia, Oreochromis niloticus (Perciformes: Cichlidae) from a tropical shallow lake in Mexico. Rev. Biol. Trop., 56: 875-884.
Direct Link - Ble, C.M., O.A. Etchian, Y.L. Alla, A.A. Adingra, S. Niamke, J.K. Diopoh and R. Arfi, 2012. Seasonal effects of the water quality in ponds on nutritive potential and digestibility of tilapia Oreochromis niloticus in natural feeding condition (Mali, West Africa). Afr. J. Agric. Res., 7: 1436-1441.
Direct Link - Dos Santos, V.B., E.A. Mareco and M.D.P. Silva, 2013. Growth curves of Nile tilapia (Oreochromis niloticus) strains cultivated at different temperatures. Acta Scientiarum. Anim. Sci., 35: 235-242.
CrossRefDirect Link - Azaza, M.S., N. Khiari, M.N. Dhraief, N. Aloui, M.M. Kraiem and A. Elfeki, 2015. Growth performance, oxidative stress indices and hepatic carbohydrate metabolic enzymes activities of juvenile Nile tilapia, Oreochromis niloticus L., in response to dietary starch to protein ratios. Aquacult. Res., 46: 14-27.
CrossRefDirect Link - Katya, K., M.Z.S. Borsra, G. Kuppusamy, M. Herriman and N.A. Ali, 2017. Preliminary study to evaluate the efficacy of bambara groundnut (Vigna subterranea (L.) Verdc.) meal as the dietary carbohydrate source in Nile tilapia, Oreochromis niloticus. Madridge J. Aquacult. Res. Dev., 1: 13-17.
CrossRefDirect Link - Shiau, S.Y. and M.S. Lei, 1999. Feeding strategy does affect carbohydrate utilization by hybrid tilapia Oreochromis niloticus×O. aureus. Fish. Sci., 65: 553-557.
CrossRefDirect Link - Soro, R.Y., J.K. Diopoh, R.M. Willemot and D. Combes, 2007. Enzymatic synthesis of polyglucosylfructosides from sucrose alone by a novel α-glucosidase isolated from the digestive juice of Archachatina ventricosa (Achatinideae). Enzyme Microb. Technol., 42: 44-51.
CrossRefDirect Link - Cisse, M., L.T. Zoue, Y.R. Soro, R.M. Meganaou and S. Miamke, 2013. Physicochemical and functional properties of starches of two Quality Protein Maize (QPM) grown in Cote d'Ivoire. J. Applied Biosci., 66: 5130-5139.
CrossRefDirect Link - Yeo, G.M., M.C. Ble, K.A. Otchoumou, S. Dabonne, L.A. Yao and A.O. Etchian, 2017. Digestibility and growth performance in fingerlings of tilapia Oreochromis niloticus fed with diet containing high-carbohydrate ingredients. Int. J. Fish. Aquat. Stud., 5: 171-175.
Direct Link - Yeo, M.G., C.M. Ble, S. Dabonne, Y.L. Alla, A.K. Otchoumou and A.O. Etchian, 2017. Effect of dietary gelatinized starch on growth performance, feed utilization and body composition of Nile tilapia (Oreochromis niloticus L.). Int. J. Food Sci. Nutr., 2: 125-129.
Direct Link - AOAC., 2000. Official Methods of Analysis of AOAC International. 17th Edn., Association of Official Analytical Chemists, Gaithersburg, Maryland.
Direct Link - Taniguchi, H. and Y. Honnda, 2009. Amylases. In: Encyclopedia of Microbiology, Schaechter, M. (Ed.). 3rd Edn., Academic Press, New York, USA., ISBN: 978-0-12-373944-5, pp: 159-173.
Direct Link - Kulshrestha, S., P. Tyagi, V. Sindhi and K.S. Yadavilli, 2013. Invertase and its applications-a brief review. J. Pharm. Res., 7: 792-797.
CrossRefDirect Link - Bernfeld, P., 1955. Amylases, α and β. In: Methods in Enzymology, Volume 1: Preparation and Assay of Enzymes, Colowick, S.P. and N.O. Kaplan (Eds.). Chapter 17, Academic Press Inc., New York, USA., ISBN-13: 9780121818012, pp: 149-154.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Hixson, S.M., 2014. Fish nutrition and current issues in aquaculture: The balance in providing safe and nutritious seafood, in an environmentally sustainable manner. J. Aquacult. Res. Dev., Vol. 5.
CrossRefDirect Link - De Silva, S.S., K.F. Shim and A.K. Ong, 1990. An evaluation of the method used in digestibility estimations of a dietary ingredient and comparisons on external and internal markers and time of faeces collection in digestibility studies in the fish Oreochromis aureus (Steindachner). Reprod. Nutr. Dev., 30: 215-226.
PubMedDirect Link - Abadom, F.E. and D.W. Milsom, 1991. Digestive enzymes of the West African giant land snail, Archachatina marginata. Biochem. Soc. Trans., 19: 334S-334S.
CrossRefPubMedDirect Link - Flari, V. and M. Lazaridou-Dimitriadou, 1996. Evolution of digestion of carbohydrates in the separate parts of the digestive tract of the edible snail Helix lucorum (Gastropoda: Pulmonata: Stylommatophora) during a complete 24-hour cycle and the first days of starvation. J. Comp. Physiol. B, 165: 580-591.
CrossRefDirect Link - Saki, S.J., T.B. Sea, K.M. Koffi, Y.R. Soro, K.A. Kra and K.J. Diopoh, 2014. Purification and physicochemical characterization of the α-glucosidase of the digestive juice of the snail Limicolria flammea (Muller 1774). Int. J. Plant Anim. Environ. Sci., 4: 376-388.
Direct Link - Castillo, S. and D.M. Gatlin III, 2015. Dietary supplementation of exogenous carbohydrase enzymes in fish nutrition: A review. Aquaculture, 435: 286-292.
CrossRefDirect Link - Ao, T., 2011. Using Exogenous Enzymes to Increase the Nutritional Value of Soybean Meal in Poultry Diet. In: Soybean and Nutrition, El-Shemy, H. (Ed.). Chapter 10, InTech Publisher, Rijeka, Croatia, ISBN 978-953-307-536-5, pp: 201-214.
Direct Link - Guionie, O., C. Moallic, S. Niamke, G. Placier, J.P. Sine and B. Colas, 2003. Identification and primary characterization of specific proteases in the digestive juice of Archachatina ventricosa. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 135: 503-510.
CrossRefDirect Link - Ademolu, K.O., O.D. Fakeye, G.A. Dedeke, O.A. Ajayi and A.B. Idowu, 2013. Digestive enzymes in African giant land snail (Archachatina marginata) during aestivation. Arch. Zootec., 62: 73-77.
CrossRefDirect Link - Al-Tameemi, R., A. Aldubaikul and N.A. Salman, 2010. Comparative study of α-amylase activity in three Cyprinid species of different feeding habits from Southern Iraq. Turk. J. Fish. Aquat. Sci., 10: 411-414.
CrossRef - Fernandez, I., F.J. Moyano, M. Diaz and T. Martinez, 2001. Characterization of α-amylase activity in five species of Mediterranean sparid fishes (Sparidae, Teleostei). J. Exp. Mar. Biol. Ecol., 262: 1-12.
CrossRefDirect Link - Bhilave, M.P., V.B. Nalawade and J.J. Kulkarni, 2014. Amylase activity of fingerlings of freshwater fish Labeo rohita fed on formulated feed. Int. J. Fish. Aquat. Stud., 2: 53-56.
Direct Link - Thongprajukaew, K., S. Kovitvadhi, U. Kovitvadhi and P. Preprame, 2017. Effects of feeding frequency on growth performance and digestive enzyme activity of sex-reversed Nile tilapia, Oreochromis niloticus (Linnaeus, 1758). Agric. Nat. Resour., 51: 292-298.
CrossRefDirect Link - Chaudhuri, A., S. Mukherjee and S. Homechaudhuri, 2012. Diet composition and digestive enzymes activity in carnivorous fishes inhabiting mudflats of Indian Sundarban estuaries. Turk. J. Fish. Aquat. Sci., 12: 265-275.
CrossRefDirect Link - Moreau, Y., V. Desseaux, R. Koukiekolo, G. Marchis-Mouren and M. Santimone, 2001. Starch digestion in tropical fishes: Isolation, structural studies and inhibition kinetics of α-amylases from two tilapias Oreochromis niloticus and Sarotherodon melanotheron. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 128: 543-552.
CrossRefDirect Link - Fagbenro, O., O. Adedire, O. Fateru, I. Oluwabukola and O. Ogunlana et al., 2005. Digestive enzyme assays in the gut of Oreochromis niloticus Linnaeus 1757, Parachanna (Channa) obscura Gunther 1861 and Gymnarchus niloticus Cuvier 1829. Anim. Res. Int., 2: 292-296.
CrossRefDirect Link - Lupatsch, I., G.W. Kissil and D. Sklan, 2003. Comparison of energy and protein efficiency among three fish species gilthead sea bream (Sparus aurata), European sea bass (Dicentrarchus labrax) and white grouper (Epinephelus aeneus): Energy expenditure for protein and lipid deposition. Aquaculture, 225: 175-189.
CrossRefDirect Link - Shiau, S.Y. and C.Y. Peng, 1993. Protein-sparing effect by carbohydrates in diets for tilapia, Oreochromis niloticus×O. aureus. Aquaculture, 117: 327-334.
CrossRefDirect Link - Smith, R.R., 1971. A method for measuring digestibility and metabolizable energy of fish feeds. Progr. Fish.-Cult., 33: 132-134.
Direct Link - Page, J.W. and J.W. Andrews, 1973. Interactions of dietary levels of protein and energy on channel catfish (Ictalurus punctatus). J. Nutr., 103: 1339-1346.
CrossRefPubMedDirect Link