
Elihasridas
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, Andalas University, Kampus Limau Manis, Padang 25163,
Indonesia
Mardiati Zain
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, Andalas University, Kampus Limau Manis, Padang 25163,
Indonesia
LiveDNA: 62.19662
Yetti Marlida
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, Andalas University, Kampus Limau Manis, Padang 25163,
Indonesia
LiveDNA: 62.22962
Andri
Department of Animal Production, Faculty of Animal Science, Andalas University, Kampus Limau Manis, Padang 25163, Indonesia
Pakistan Journal of Nutrition
Year: 2019 | Volume: 18 | Issue: 1 | Page No.: 60-66







ABSTRACT
Background and Objective: This study aimed to determine the energy and protein balance in male Simmental cattle when fed diets with different protein and energy levels. Methodology: The experimental design used a randomized block design (RBD) with a 2×3×3 factorial pattern. There were two factors, namely, Factor A, which was the level of dietary energy (65 or 70% TDN) and Factor B, which was the level of dietary protein (10, 12 or 14%). Each treatment was repeated 3 times. The diet consisted of ammoniated rice straw and a concentrate and was fed at a ratio of 40:60. The concentrate consisted of palm kernel cake, rice bran, cassava, mineral and urea. The measured variables were intake; the digestibility of dry matter (DM), organic matter (OM) and crude protein (CP), daily gain, urine allantoin, blood metabolites and feed efficiency. Results: There was no interactive effect between the protein and energy levels of the diet on nutrient intake (DM intake, OM intake and CP intake), weight gain, or efficiency (p<0.05). There was an interactive effect between the dietary protein and energy levels on nutrient digestibility (DM, OM and CP) and urine allantoin (p>0.01). Nutrient intake tended to decrease with increasing levels of energy and protein in the diet, while nutrient digestibility increased with increasing energy levels in the diet but did not significantly change with increasing protein levels in the diet. The average daily gain and feed efficiency improved with the increasing levels of energy in the diet. There was an interactive effect (p<0.05) between the energy and protein levels on the cholesterol and HDL levels. Cholesterol levels increased with increasing levels of protein in the ration. There was no interactive effect (p>0.05) between the protein and energy levels on LDL, total protein, glucose, albumin, globulin, calcium or urea. Conclusion: It can be concluded that treatment A2B2, namely, an energy level of 70% TDN and a protein level of 12%, could provide the best reproductive performance of Simmental cattle in tropical areas.
PDF Abstract XML References Citation
Received: July 28, 2018;
Accepted: October 05, 2018;
Published: December 15, 2018
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Elihasridas, Mardiati Zain, Yetti Marlida and Andri, 2019. Effect of the Level of Energy and Dietary Protein of Ammoniated Rice Straw on the Productivity of Male Simmental Cattle in a Tropical Area. Pakistan Journal of Nutrition, 18: 60-66.
DOI: 10.3923/pjn.2019.60.66
URL: https://scialert.net/abstract/?doi=pjn.2019.60.66
DOI: 10.3923/pjn.2019.60.66
URL: https://scialert.net/abstract/?doi=pjn.2019.60.66
INTRODUCTION
Protein and energy are important nutrients to consider when formulating rations for ruminant livestock. Abubakar et al.1showed that to improve livestock productivity, nutrients can be manipulated by adjusting the feed protein-energy ratio. Livestock productivity depends on not only genetic potential but also the fulfillment of nutritional needs aimed at maximizing animal production. Energy plays an important role in supporting daily gain as well as animal health2. Dietary energy and protein levels affect the ruminal fermentation behaviors associated with total amino acid composition and microbial synthesis in ruminants3. Rations with a high energy content will lead to carbohydrate fermentation and an increase in VFA in the rumen4, which will be absorbed through the walls of the reticulum and omasum, with some passing into the abomasum and the small intestine as an energy source for the animals5. A lack of energy in the diet will lead the animal to catabolize body reserves, including body protein, which will raise the urea concentration in the blood6. Additional protein and energy supplements are needed for livestock feed that is of low quality due to poor management7.
Energy and protein are the most important sources of nutrients for beef cattle, as these nutrients stimulate rumen micro organism growth and rumen fermentation and enhance the productive functions of the animals. In the rumen, simultaneous carbohydrate fermentation is essential for microbial protein synthesis8. This study aimed to balance the protein and energy needs of male Simmental cattle that live in tropical areas, such as Indonesia. No previous studies have reported on the protein and energy balances of male Simmental cows in Indonesia. The purpose of the present research was to determine the ideal energy and protein balances of male Simmental cattle diets to obtain the best reproductive performance.
MATERIAL AND METHODS
The experimental design used was a randomized block design (RBD) with a 2×3×3 factorial pattern consisting of 6 treatment rations and three groups of body weight as replicates. The treatment consists of 2 factors: Factor A = TDN level (A1: TDN 65%, A2: TDN 70%) and Factor B = protein level (B1: protein 10%, B2: protein 12% and B3: protein 14%). Eighteen Simmental male cattle with initial body weights of approximately 225-260 kg were used in this study. The treatment diets are shown in Table 1. Digestibility trials were conducted using three animals separated into individual pens for each treatment. Cattle were fed ad libitum during the preliminary phase (15 days) and were then restricted during the collection period (last 6 days) to 90% of the feed that was offered at 7:00 and 16:00 h. During the collection period, accurate records were kept on individual feed intake and fecal excretion. Live weight was measured every two weeks. Total fecal excretion was collected once daily and 10% of the representative samples were dried at 60°C overnight and kept in sealed bags until analysis. Feed and fecal materials were ground to pass through a 1 mm screen and were composited. Dry matter, organic matter and nitrogen were analyzed by standard methods. Samples of feed, feed refusals and feces were analyzed for dry matter (DM), organic matter (OM) and crude protein (CP) using standard methods9. Urine allantoin levels were analyzed according to Chen et al.10. Blood samples were taken once, before the livestock were fed (fasting). Blood samples were taken from a jugular vein using a 10 mL capacity syringe and placed in a vacutainer. The blood serum was separated by centrifugation at a rate of 3000 rpm for 10 min. Glucose, total protein, urea, albumin, triglyceride, total cholesterol, HDL and LDL analyses were performed using the HumaStar 80® Auto Analyzer tool. Data were analyzed using ANOVA and means were compared using Duncan multiple range test11.
| Table 1: | The Ingredient composition of and the chemical composition (dry matter%) |
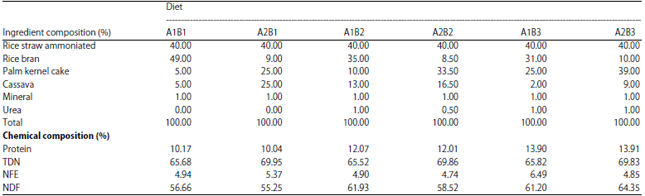 | |
RESULTS AND DISCUSSION
Nutrient intake: The effect of treatment on nutrient intake is shown in Table 2. The results showed that there was no interactive effect (p>0.01) between the protein and energy levels on dry matter, organic matter and protein intake. This finding indicates that different energy and protein ratios have no effect on dry matter, organic matter and protein intake. The average dry matter and organic matter intake from all treatments in this study were 7.06 and 6.57 kg h–1, respectively.
Table 2 shows that higher protein and energy levels caused dry matter and organic matter intake to be lower. Dry matter intake was influenced by the type of concentrate, nutrient quality and the rate of degradation. This finding is supported by that of Davidson et al.12, who reported that rations with increased protein levels (16.5, 16.8, 17.2 and 19.2%) in dairy cattle tended to decrease the dry matter intake (24.1, 23.4, 23.1 22.9 and 23.3 kg day–1, respectively). The decreased dry matter intake associated with increased levels of protein was caused by higher concentrations of proteins that escaped rumen fermentation and decreased rumen fermentation activity, resulting in decreased dry matter intake13. In addition, Tauqir et al.14 reported an increase in dry matter intake in buffaloes fed a low protein content and a decrease in dry matter intake as protein levels increased.
The dry matter intake of the treatments was 5.67-6.78 kg days–1, equivalent to 2-2.5% of body weight. It is generally accepted that animal intake of low-quality tropical forages is usually limited by a low content of nutrients, especially proteins that are essential for the growth of animal microbes in the rumen. However, protein supplements may not be used effectively in the rumen if an energy source is not available. Under such conditions, the protein is damaged, becomes an amino acid and undergoes a deamination process. In ruminants, urea is an end product of N metabolism but bacteria have the enzymatic capability to hydrolyzeurea, thereby making urea-N available as NH3. The resulting N-ammonia is absorbed by the animal and excreted in urine in the form of urea15.
Nutrient digestibility: The results of the research showed that there was a significantly different interactive effect between protein and energy on the digestibility of dry matter, organic matter, crude protein and urine allantoin production (p<0.05) (Table 2). Increasing the energy and protein in the diet appeared to increase nutrient digestibility. The higher the availability of dietary energy is, the greater the increase in the energy supply and the better the optimization of rumen microbial growth so that the amount of degradable feed will increasingly improve digestibility of the nutrients.
The digestibility of dry matter and organic matter was significantly different (p<0.05) in the higher protein diet, although no particular trend with respect to varying levels of dietary CP was shown. The digestibility of protein has been reported to increase linearly with increasing dietary protein level16,17. However, no effect of dietary protein levels on DM digestibility was observed in this study, although it was reported in other similar studies18.
Digestibility of DM was significantly different (p<0.05) between diets containing 65 and 70% TDN. The digestibility of DM in cattle diets with high TDN and CP levels was significantly higher than in diets with low TDN and CP levels. The high digestibility of DM and OM in diets with high CP levels is caused by low levels of feed intake. High feed intake will decrease digestion19. Low feed intake and feed degradation rates result in feed leaving the rumen more slowly and a longer digestion time. In terms of feed, digestibility is influenced by the type, amount, composition and intake rate of rations given to livestock20. Other studies have suggested that the digestibility of DM, OM and protein increases when cattle are fed a 13% protein diet21. Archibeque et al.22 stated that dry matter digestibility increased from 71.8-75.8% with an increase in the proteinconcentration in the diet. This finding is supported by a study of Gabler and Heinrichs23, who reported that a ration that contained a high proportion of concentrate could also increase the digestibility of dry matter in cattle because it had high protein and energy content.
| Table 2: | Effect of treatment on intake, digestibility, weight gain and feed conversion |
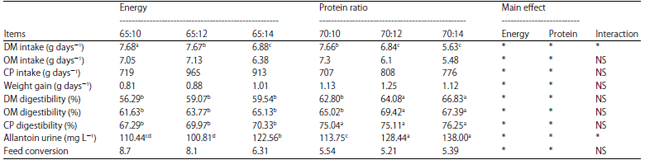 | |
| Means within row bearing different superscripts differ significantly (p<0.05) | |
The digestibility of protein in this study was significantly different (p<0.05). This difference was presumably because the availability of N in the rumen for microbial protein synthesis is high, allowing feed proteins to bypass degradation by rumen microbes and to be digested in the intestine. Protein digestibility is also influenced by protein type and its interaction with other nutrients (feed type, flow rate and pH rumen)24. The utilization of energy is influenced by the quality of feed consumed, including crude protein balance and the total digestible nutrients (TDN) or energy. The need for feed with a balanced protein-energy ratio is greater in rapidly growing young ruminants25. The balanced protein-energy ratio will support optimal fermentation efficiency, in which case, the feed energy utilized for the process will also be optimal26.
Table 2 also shows that the energy content of the ration significantly affects daily weight gain (p<0.01). The increase in body weight corresponds to the increasing energy content of the ration. Increasing the energy level allows the production of more fermentable ME for paunch microorganisms, resulting in increases in the synthesis of microbial proteins and in the amount of protein available to the animal27. This is likely to be the reason why daily weight gain increased when the energy levels increased from 65-70% TDN. In contrast, increased protein content does not significantly increase body weight, in agreement with the findings of Huuskonen and Huhtanen28, who concluded that the body weight gain of growing cattle increased with increasing ME intake [ME/BW(0.75)] and showed only marginal effects of the protein supply on body weight gain. This result is also consistent with the findings of Huuskonen et al.29, who reported that the daily weight gain response of cattle fed a control diet was not related to the protein concentration of the diet and that increased protein supplementation was negatively related to the DMI and ME intake. Haryanto30 reported that animal conditions will influence energy requirements as well as environmental factors. In the tropics, the energy need will be higher than the energy need in a subtropical region due to the quality of feed being relatively lower in general in the tropics. Low-quality feed causes a higher heat increment and results in lower feed efficiency. Heat increment is the energy that livestock emit in the gastrointestinal tract during the feed digestion process. Research results show that the energy needs for live cattle (maintenance) in the tropics are approximately 30% higher than those for live cattle in subtropical regions.
The results of this research show that the best average daily gain obtained from a diet with 70% TDN and 12% CP was 1.25 kg days–1. The average daily gain obtained in this research was higher than that reported by Tufan et al.31, who found that the average daily weight gain of Simmental beef cattle that received rations of hay and corn ranged from 0.9-1 kg d–1. Increasing crude protein in the diet from 10-14% did not significantly affect the average daily gain. It could be concluded that 12% crude protein in the diet was able to provide adequate digestible protein for the intestines to support the gains attained; there was no response in performance due to increasing the protein intake beyond this level. This result agrees with those of Schroeder and Titgemeyer32, who reported that when the supply of digestible protein exceeds the animal’s requirements, additional protein intake does not affect protein deposition.
The increasing TDN level could increase the body weight gain of Simmental cattle. The highest increase in body weight gain was observed at the energy level of 70% with a crude protein level of 12%, i.e., 1.25 kg h–1. Furthermore, Kearl33 stated that feed energy requirements will increase as body weight increases. The balance of protein and energy in the ration greatly determines the efficiency of nutrient utilization, which ultimately affects the productivity of cattle20. Furthermore, Wanapat8 reported that energy and protein are important sources of nutrients for beef cattle and stimulate the growth of microorganisms and rumen fermentation.
Urinary allantoin can describe the synthesis of microbial proteins. Increased urinary allantoin levels may indicate increased synthesis of microbial proteins. Table 2 shows that the interaction between energy and protein levels has a significant effect (p<0.05) on the urinary allantoin levels. The highest urine allantoin level was at the energy level of 65% TDN and 14% protein and was not significantly different from the energy level of 70% TDN and 14% protein. From these results, it was observed that at the level of 14% protein, there is an increase in the protein synthesis of microbes, which is also characterized by an increase in the digestibility of feed during treatment. Increased levels of energy and protein in rations also cause increased urine allantoin. This increase was due to the availability of VFA from carbohydrates and NH3 from protein, which was sufficient for rumen microbial growth. This observation was supported by Lamothe et al.34 and Suryani et al.35, who reported that allantoin is the ultimate result of purine metabolism, which is the most effective indicator for determining the amount of microbial protein formation.
The interaction of energy and protein levels did not have a significant effect (p>0.01) on feed efficiency (Table 2). The greater amount of ration efficiency indicates that the rationis becoming more digestible. Shahzad et al.36 reported no differences in the ADG and feed efficiency of buffalo consuming different dietary energy and protein levels. In a contrasting study, Hoffman et al.21 reported linear and quadratic relationships between the dietary CP and weight gains of Holstein heifers. Feed conversion in this study was obtained at 5.21-8.70. Feed conversion was influenced by biological capabilities and the interaction with the environment and generally, the value of feed conversion was good for cattle, ranging from 4.5-7.537,38.
Blood metabolic profiles: The effect of treatment on blood profiles is presented in Table 3. The results showed that there was an interactive effect (p<0.05) between energy level and protein on cholesterol and HDL levels. Cholesterol levels increase with increasing levels of protein in the rations. The lowest cholesterol levels were found in treatment A1B1 and the highest was in treatment A1B3. This result is higher than the results of Radostits et al.39, who obtained cow blood cholesterol in the normal range of 161.95 mg dL–1. These different blood cholesterol levels likely occur as a result of rations with different compositions.
Table 3 shows that the HDL levels obtained were higher than the LDL levels. The highest HDL was in the A1B2 treatment. Andersen et al.40stated that high levels of HDL were important because HDL also serves as an antioxidant and anticoagulant that can prevent the occurrence of various diseases in the bodies of livestock.
There was no interactive effect (p>0.05) between the protein and energy levels on LDL, total protein, glucose, albumin, globulin, calcium or urea. Increased total protein indicates a positive outcome, as there was a decrease in the proteolytic process in the rumen through the inhibition of amino acid conversion to ammonia41. Oikonomou et al.42 reported a significant influence of the glucose concentration on reproductive performance, as glucose is an essential energy source for ovarian cyclicity. From the results obtainedin this study, glucose ranged from 83.6-88.8 mg dL–1. This result is quite normal. Normally, these blood glucose levels exist in addition to the energy supplied by the feed and because of the control mechanism of insulin and glucagon that regulates the balance of blood glucose in livestock. Blood glucose levels in this study were higher than those in a study conducted by Munzaronah et al.43, who observed an average blood glucose level of 75.27 mg dL–1 in Java cattle fed different protein levels. This discrepancy likely occurred because the level of the ration digestibility in this study was high enough to support an elevated supply of nutrients and glucose production in livestock and is probably better than the levels found by Munzaronah et al.43. Syahrir et al.44 reported that in male PO cattle that received 50% rice straw feed and 50% concentrate, the blood glucose level was 70-76 mg dL–1. Differences in blood glucose levels were observed in the various studies mentioned above because feed factors, especially energy composition, greatly determine the level of blood glucose. In the reports above, the resulting blood glucose levels are higher because, in addition to using forage, researchers also utilized concentrates in the ration. The availability of calcium in the body is very important with respect to its role in bone and tooth formation, as well as its role in physiological and biochemical processes in the body45. The calcium levels that were obtained in this study ranged from 7.5-10.2 mg dL–1. This result was in accordance with the standard requirement of minerals for cows, which is 8-12 mg dL–1 46. Blood urea can be used as an index of protein use. The results of this study showed that the blood urea levels of the Simmental cattle ranged from 28.31-34.48 mg dL–1. This result is quite normal and is supported by Hungate47, who stated that the normal range of blood urea levels is 26.6-56.7 mg dL–1. Measurement of the blood urea concentration is an indicator of the utilization of NH3 and energy in the rumen. The efficiency of NH3 utilization for protein synthesis in the rumen depends on energy availability. If there is a lack of energy, the protein will be presented inexcessive amounts and cannot be utilized by rumen microbes. It can be concluded that a high concentration of urea in plasma is caused by high dietary crude protein48.
| Table 3: | Effect of treatment on blood metabolic parameters |
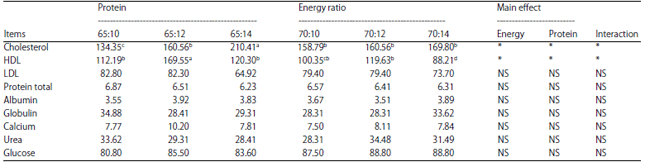 | |
| Means within row bearing different superscripts differ significantly (p<0.05) | |
CONCLUSION
High energy but low protein content in the diet (TDN 70%, crude protein 12%) produced the best response in terms of intake, digestibility, weight gain, ration efficiency and blood metabolic profiles of Simmental beef cattle in a tropical area.
SIGNIFICANCE STATEMENT
The study discover the proper energy and protein ratio to increase the productivity of Simmental cattle in the tropical area. Generally, the data is available on protein and energy requirements for temperate regions. This study will helps researchers to formulate rations to increase the productivity of Simmental cattle that are kept in tropical areas based on agricultural waste and to get the feed affordable by the farmers. Thus introduced a new approach to produce ration from rice straw ammoniated that contain energy and protein to meet requirements of livestock for tropical area.
ACKNOWLEDGMENT
S
The authors would like to acknowledge the financial support provided by PUSN Grant 2017 By Ristek Dikti with contract No. 02/ADD/UN.16./7IPP.PUSN/LPPM/2017. Special thanks are given to the Surya Farm for providing the cattle and performing the feeding procedures.
REFERENCES
- Abubakr, A.R., A.R. Alimon, H. Yaakub, N. Abdullah and M. Ivan, 2013. Digestibility, rumen protozoa and ruminal fermentation in goats receiving dietary palm oil by-products. J. Saudi Soc. Agric. Sci., 12: 147-154.
CrossRefDirect Link - Rotger, A., A. Ferret, S. Calsamiglia and X. Manteca, 2006. Effects of nonstructural carbohydrates and protein sources on intake, apparent total tract digestibility and ruminal metabolism in vivo and in vitro with high-concentrate beef cattle diets. J. Anim. Sci., 84: 1188-1196.
CrossRefDirect Link - Mawati, S., A.P. Adi, Soedarsono and Sunarso, 2016. Effect of feed with different energy-protein ratios on parameters of sheep ruminal fermentation. Pak. J. Nutr., 15: 1055-1060.
CrossRefDirect Link - Greenwood, P.L., A.S. Hunt, R.M. Slepetis, K.D. Finnerty, C. Alston, D.H. Beermann and A.W. Bell, 2002. Effects of birth weight and postnatal nutrition on neonatal sheep: III. Regulation of energy metabolism. J. Anim. Sci., 80: 2850-2861.
PubMed - Boonsaen, P., N.W. Soe, W. Maitreejet, S. Majarune, T. Reungprim and S. Sawanon, 2017. Effects of protein levels and energy sources in total mixed ration on feedlot performance and carcass quality of Kamphaeng Saen steers. Agric. Nat. Resour., 51: 57-61.
CrossRefDirect Link - Wanapat, M., 2009. Potential uses of local feed resources for ruminants. Trop. Anim. Health. Prod., 41: 1035-1049.
CrossRefDirect Link - Chen, X.B., Y.B. Chen, M.F. Frankiln, E.R. Orskov and W.J. Shand, 1992. The effect of feed intake and body weight on purine derivative excretion and microbial protein supply in sheep. J. Anim. Sci., 70: 1534-1542.
PubMedDirect Link - Davidson, S., B.A. Hopkins, D.E. Diaz, S.M. Bolt, C. Brownie, V. Fellner and L.W. Whitlow, 2003. Effects of amounts and degradability of dietary protein on lactation, nitrogen utilization and excretion in early lactation holstein cows. J. Dairy Sci., 86: 1681-1689.
CrossRefDirect Link - Allen, M.S., 2000. Effects of diet on short-term regulation of feed intake by lactating dairy cattle. J. Dairy Sci., 83: 1598-1624.
CrossRefPubMedDirect Link - Tauqir, N.A., M.A. Shahzad, M. Nisa, M. Sarwar, M. Fayyaz and M.A. Tipu, 2011. Response of growing buffalo calves to various energy and protein concentrations. Livestock Sci., 137: 66-72.
CrossRefDirect Link - Tan, Z. and M.R. Murphy, 2004. Ammonia production, ammonia absorption and urea recycling in ruminants. A review. J. Anim. Feed Sci., 13: 389-404.
CrossRefDirect Link - Dabiri, N. and M.L. Thonney, 2004. Source and level of supplemental protein for growing lambs. J. Anim. Sci., 82: 3237-3244.
PubMedDirect Link - Promkot, C. and M. Wanapat, 2005. Effect of level of crude protein and use of cottonseed meal in diets containing cassava chips and rice straw for lactating dairy cows. Asian Aust. J. Anim. Sci., 18: 502-511.
Direct Link - Jiang, Q., G. Li, T. Zhang, H. Zhang and X. Gao et al., 2015. Effects of dietary protein level on nutrients digestibility and reproductive performance of female mink (Neovison vison) during gestation. Anim. Nutr., 1: 65-69.
CrossRefDirect Link - Hoffman, P.C., N.M. Esser, L.M. Bauman, S.L. Denzine, M. Engstrom and H. Chester-Jones, 2001. Short communication: Effect of dietary protein on growth and nitrogen balance of Holstein heifers. J. Dairy Sci., 84: 843-847.
Direct Link - Archibeque, S.L., H.C. Freetly, N.A. Cole and C.L. Ferrell, 2007. The influence of oscillating dietary protein concentrations on finishing cattle. II. Nutrient retention and ammonia emissions. J. Anim. Sci., 85: 1496-1503.
CrossRefDirect Link - Gabler, M.T. and A.J. Heinrichs, 2003. Dietary protein to metabolizable energy ratios on feed efficiency and structural growth of prepubertal holstein heifers. J. Dairy Sci., 86: 268-274.
Direct Link - Bach, A., S. Calsamiglia and M.D. Stern, 2005. Nitrogen metabolism in the rumen. J. Dairy Sci., 88: E9-E21.
CrossRefDirect Link - Porto, M.O., M.F. Paulino, S.C. Valadares Filho, E. Detmann and J. Cavali et al., 2012. Nutritional requirements of energy, protein and macrominerals for maintenance and weight gain of young crossbred Nellore x Holstein bulls on pasture. R. Bras. Zootec., 41: 734-745.
CrossRefDirect Link - Ginting, S.P., 2005. [Synchrony of protein and energy in the rumen to maximize the production of microbial protein]. WARTAZOA, 15: 1-10.
Direct Link - Early, R.J., O. Mahgoub and C.D. Lu, 2001. Energy and protein utilization for maintenance and growth in Omani ram lambs in hot climates. II. Composition of tissue growth and nitrogen metabolism. J. Agric. Sci., 136: 461-470.
CrossRefDirect Link - Huuskonen, A. and P. Huhtanen, 2015. The development of a model to predict BW gain of growing cattle fed grass silage-based diets. Animal, 9: 1329-1340.
CrossRefDirect Link - Huuskonen, A., P. Huhtanen and E. Joki-Tokola, 2014. Evaluation of protein supplementation for growing cattle fed grass silage-based diets: A meta-analysis. Animal, 8: 1653-1662.
CrossRefDirect Link - Tufan, T., C. Arslan, K. Onk, M. Sari and M. Tilki, 2016. Effects of feeding by hay, grass silage and corn silage on growth performance, rumen fluid and blood serum parameters in beef cattle. Revue De Medecine Veterinaire, 167: 99-105.
Direct Link - Schroeder, G.F. and E.C. Titgemeyer, 2008. Interaction between protein and energy supply on protein utilization in growing cattle: A review. Livestock Sci., 114: 1-10.
CrossRefDirect Link - Suryani, H., M. Zain, R.W.S. Ningrat and N. Jamarun, 2017. Effect of dietary supplementation based on an ammoniated palm frond with direct fed microbials and virgin coconut oil on the growth performance and methane production of Bali cattle. Pak. J. Nutr., 16: 599-604.
CrossRefDirect Link - Shahzad, M.A., N.A. Tauqir, F. Ahmad, M.U. Nisa, M. Sarwar and M.A. Tipu, 2011. Effects of feeding different dietary protein and energy levels on the performance of 12-15-month-old buffalo calves. Trop. Anim. Health Prod., 43: 685-694.
CrossRefDirect Link - Carstens, G.E. and L.O. Tedeschi, 2006. Defining feed efficiency in beef cattle. Proceedings of the 38th Beef Improvement Federation Annual Research Symposium, April 2006, Choctaw, Mississippi, pp-12.
CrossRef - Arthur, J.P. and R.M. Herd, 2008. Residual feed intake in beef cattle. Revista Brasileira de Zootecnia, 37: 269-279.
CrossRefDirect Link - Afzalani, M. Zein, N. Jamarun and E. Musnandar, 2015. Effect of increasing doses of essential oil extracted from Berastagi orange (Citrus sinensis L.) peels on performance, rumen fermentation and blood metabolites in fattening Bali cattle. Pak. J. Nutr., 14: 480-486.
CrossRefDirect Link - Oikonomou, G., G. Arsenos, G.E. Valergakis, A. Tsiaras, D. Zygoyiannis and G. Banos, 2008. Genetic relationship of body energy and blood metabolites with reproduction in holstein cows. J. Dairy Sci., 91: 4323-4332.
CrossRef - Munzaronah, Soedarsono, C.M.S. Lestari, E. Purbowati and A. Purnomoadi, 2010. [Blood parameters of java cattle under different protein levels of feeding]. Proceedings of the Prodising Seminar Nasional Teknologi Peternakan dan Veteriner Pusat Penelitian dan Pengembangan Peternakan, August 3-4, 2010, Bogor, pp: 243-248.
- Syahrir, S., K.G. Wiryawan, A. Parakkasi and M. Winugroho, 2010. [Blood profile of beef cattle offered mulberry leave meal to substitute feed concentrate]. JITP, 1: 12-18.
Direct Link - Bindari, Y.R., S. Shrestha, N. Shrestha and T.N. Gaire, 2013. Effects of nutrition on reproduction-A review. Adv. Applied Sci. Res., 4: 421-429.
Direct Link - Pujiastari, N.N.T., P. Suastika and N.K. Suwiti, 2015. Level of mineral calcium and iron on the bali cattle being farmedin the rice field. Buletin Vet. Undayana, 7: 66-72.
Direct Link - Moallem, U., R. Blanck, H. Lehrer, L. Livshitz, M. Zachut and A. Arieli, 2011. Effects of high dietary crude protein on the characteristics of preovulatory follicles in dairy heifers. J. Dairy Sci., 94: 785-792.
CrossRefDirect Link