
Endah Puspitojati
Magelang Agricultural Extension College, Jl.Kusumanegara, Umbulharjo, Yogyakarta 55167, Indonesia
Muhammad Nur Cahyanto
Department of Food and Agricultural Product Technology, Faculty of Agricultural Technology, Universitas Gadjah Mada, Jl. Flora, Bulaksumur, Yogyakarta 55281, Indonesia
Yustinus Marsono
Department of Food and Agricultural Product Technology, Faculty of Agricultural Technology, Universitas Gadjah Mada, Jl. Flora, Bulaksumur, Yogyakarta 55281, Indonesia
Retno Indrati
Department of Food and Agricultural Product Technology, Faculty of Agricultural Technology, Universitas Gadjah Mada, Jl. Flora, Bulaksumur, Yogyakarta 55281, Indonesia
LiveDNA: 62.18887
Pakistan Journal of Nutrition
Year: 2019 | Volume: 18 | Issue: 5 | Page No.: 464-470







ABSTRACT
Background and Objective: Angiotensin-converting enzyme (ACE) inhibitory peptides from various food sources have been studied extensively. We are interested in studying the inhibition of ACE from fermented tempe made from the jack bean, which is a commonly used source of plant-based protein. The purpose of this study was to determine the optimal incubation period for the production of peptides with high ACE inhibitory activity. Materials and Methods: After soaking and boiling, the jack beans were inoculated with tempe inoculum and incubated for 0-120 h. During the incubation, various parameters, such as proteolytic activity, peptide content, hydrolysis degree, protein pattern and ACE inhibition, were monitored. All the parameters were measured using standard methods. Results: Tempe fermentation caused hydrolysis of the jack bean protein, producing soluble high molecular weight proteins (from previously insoluble) and protein fragments (monitored from SDS PAGE data). The highest protein hydrolysis occurred after 96 h fermentation period, which were observed from the highest proteolytic activity (25.39 units g–1), hydrolysis degree (19.85%) and peptide content (6.05 mg mL–1). The highest ACE inhibitory activity did not coincide with the highest levels of protein hydrolysis but rather were obtained after 72 h of tempe fermentation (IC50 = 1.03 mg mL–1). Conclusion: Jack bean tempe is a potential source of ACE inhibitory peptides, the highest ACE inhibitory activity occurred after 72 h of incubation.
PDF Abstract XML References Citation
Received: November 21, 2018;
Accepted: February 03, 2019;
Published: April 15, 2019
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Endah Puspitojati, Muhammad Nur Cahyanto, Yustinus Marsono and Retno Indrati, 2019. Production of Angiotensin-I-Converting Enzyme (ACE) Inhibitory Peptides during the Fermentation of Jack Bean (Canavalia ensiformis) Tempe. Pakistan Journal of Nutrition, 18: 464-470.
DOI: 10.3923/pjn.2019.464.470
URL: https://scialert.net/abstract/?doi=pjn.2019.464.470
DOI: 10.3923/pjn.2019.464.470
URL: https://scialert.net/abstract/?doi=pjn.2019.464.470
INTRODUCTION
Angiotensin-converting enzyme (ACE) plays an important role in blood pressure regulation. This enzyme converts inactive angiotensin I to angiotensin II which causes an increase in blood pressure1. The ACE inhibitory peptide possesses biological activity to inhibit the function of ACE2. Studies on food derived ACE-inhibitory peptides have been widely reported. Vegetable proteins are potential sources of ACE inhibitory peptides because they can be produced in a cost-efficient and environmentally friendly manner compared to animal proteins. Additionally, vegetable derived peptides can be an alternative protein source for vegetarians3. The activity of ACE inhibitory peptides are influenced by the protein sources, chain length, amino acids composition and peptide sequences2,4. The peptides can be prepared by solvent extraction2, enzymatic hydrolysis5,6 or fermentation7-11.
Tempe is a fermented food obtained from protein-rich beans through solid-fungal fermentation. The inoculated cooked beans were wrapped in plastic or banana leaves and incubated at 30-31°C until the beans were compactly bound by the mycelia12. The important fungi in tempe productions are Rhizopus microspores, R. oligosporus, R. rhizopodiformis and R. chinensis13. During fermentation, proteolytic enzymes convert high molecular weight proteins to low molecular proteins. This action increases the ACE inhibitory activity. The fermentation is stopped after the substrate is evenly grown over the mycelium, requiring a time of 24-48 h for common beans such as soybean8,14.
In general, a wide variety of legumes can be made into tempe, including jack beans. Jack beans (Canavalia ensiformis) are a hard-seeded legume that have a high protein content15-17. The hydrophobic amino acid content, including proline, leucine and isoleucine, is high16. These amino acids support ACE inhibitory properties. In the process of making jack bean tempe, these seeds require a longer soaking time than most legumes in general. This immersion aims to eliminate the HCN content and soften the seeds18. Jack bean tempe is similar to soybean tempe in color, flavor, texture and acceptability15. In this study, the time variation in tempe fermentation made from jack beans was quite long (0-120 h). The hard seeds required a long period for fungi growth. This study was conducted to determine pattern of ACE inhibitory peptide production and the optimal time for production. The results were also compared with jack bean fermented tempe after a 48 h of fermentation.
MATERIALS AND METHODS
Materials: Jack bean seeds (Canavalia ensiformis) and Raprima commercial tempe inoculum (containing Rhizopus oligosporus) were from local market. Angiotensin Converting Enzyme (from rabbit lung) and HHL (Hippuryl-L-Histidyl-L-Leucine) were from Sigma-Aldrich Company. O-phtaldialdehyde (OPA) was from Merck and the protein marker was from Biorad. All other chemicals were of analytical grade.
Preparation of jack bean tempe: The jack bean seeds were washed, soaked for 24 h and then boiled for 30 min at ratio of jack bean to water of 1: 4. The boiled water was discarded and the jack beans were soaked again for 24 h. Thes seeds were then peeled and cut into 4-6 pieces followed by soaking again in tap water for 48 h. The soaking water was changed every 12 h. The sliced seeds were boiled for 30 min after which the water was discarded. The seeds were then drained and cooled (±30°C). The tempe inoculum of 0.02% was inoculated to the cold seeds, evenly mixed and, wrapped using banana leaves. The incubation was carried out at room temperatures (30°C) for 0, 12, 24, 36, 48, 72, 96 and 120 h. The resulting tempe were lyophilized and kept for further analysis.
Preparation of crude protease extracts: Protease extraction was performed according the method of Elegado and Fujio19 with slight modifications. Jack bean tempe (3 g) was added to 30 mL of phosphate buffer 0.05 M, pH 6 and then homogenized for 3 min using a warring blender. The mixture was incubated at 30°C for 30 min in a water bath shaker and then centrifuged at 9,391 g for 10 min at 4°C. The supernatant was collected and used as the enzyme solution.
Peptide extraction: Peptide extraction was performed by the method of Rusdah20 with slight modifications. The freeze dried tempe (1 g) was added to 30 mL of distilled water and homogenized using a warring blender for 3 min. The mixture was incubated at 30°C for 60 min and then centrifuged at 20,000 g for 15 min. The supernatant was the peptide sample.
Analysis methods
ACE-inhibitory activity: The ACE-inhibitory activity of jack bean tempe was determined by the method of Cushman and Cheung21 with slight modifications. The peptide sample of 50 μL was mixed with 50 μL of substrate (Hip-His-Leu 8 mM) and incubated at 37°C for 10 min. The ACE solution (25 mU mL–1) was added and incubated at the same temperature for 30 min. The reaction was terminated by the addition of 200 μL of 1 M HCl. The hippuric acid formed during the reaction process was extracted with 1.5 mL of ethyl acetate. The mixture was shaken for 120 sec and centrifuged at 4000 g at 4°C for 15 min. A 1 mL volume of the clear upper layer was collected and dried. The residue was redissolved in 3 mL of distilled water. The absorbance was determined at 228 nm using a Spectrophotometer UV-VIS (Dynamica Scientific Halo SB-10). The ACE-inhibitory activity was calculated as follows:
where, A is the absorbance in the presence of ACE, B is the absorbance in the presence of ACE and the inhibitor and C is the absorbance of the reaction blank10,22. The IC50 is the amount of peptide required for inhibiting 50% of ACE activity.
Peptide content and degree of hydrolysis (DH): The peptide sample was calculated for peptide content by the ο-phthaldialdehyde (OPA) spectrophotometric assay23. The absorbance at 340 nm was measured using a Spectrophotometer UV-VIS (Dynamica Scientific Halo SB-10). Tryptone was used to generate the standard curve24. The DH was defined as the percentage of cleaved peptide bonds as follows:
where, (NH2)tx is the number of free amino groups at X min, (NH2)t0 is the number of free amino groups at 0 min of hydrolysis and (NH2)tot is the total number of amino groups25.
Protein pattern: The protein pattern was analyzed using SDS-PAGE according to the method of Laemmli26 with a 5% resolving gel and 13% separating gel. The peptides extracts was diluted in SDS sample buffer [containing 0.5 M Tris-HCl pH 6.8, 87% glycerol (w/v), 10% SDS (w/v), 0.5% bromophenol blue (w/v) and distilled water] at a ratio of 1 : 2. The samples were heated at 100°C for 4 min. A 20 μL volume of samples or 5 μL of standard protein marker was loaded into each well. The gel was run at 220 V for approximately 60 min. The gel was soaked in distilled water for 30 min and stained with 0.2% Coomassie Brilliant Blue R-250 (containing 50% methanol, 10% acetic acid and 40% distilled water). The gel was soaked in the destaining solution for 30 min and the solution was replaced three times.
Proteolytic activity: The proteolytic activity was analyzed by the method of Elegado and Fujio19 with a slight modification. The substrate, 5 mL of casein solution 0.65%, was preincubated in a water bath shaker for 5 min at 40°C. The sample (1 mL) was added to the solution and incubated for 10 min at 40°C. The reaction was terminated by the addition of 5 mL of TCA. The mixture was centrifuged at 6000 g for 10 min. The supernatant was collected and then 5 mL of sodium carbonate was added followed by the addition of 1 mL of Folin’s reagent. These solutions were mixed evenly and incubated for 10 min at 40°C. The absorbance was measured at 660 nm using a Spectrophotometer UV-VIS (Dynamica Scientific Halo SB-10). One proteolytic activity unit (U) was defined as the amount of the enzyme that releases 1 μmol of tyrosine per min under the assay conditions.
Proximate and amino acids composition analysis: The proximate (moisture, ash, protein and fat contents) and amino acid composition of the jack bean were determined according to the official AOAC procedures27.
Statistical analysis: The data were analyzed using one-way analysis of variance (ANOVA) and the mean differences between treatments were analyzed by Duncans Multiple Range Tests (DMRT). Statistical analysis was performed using SPSS IBM 23 and the significance difference was determined with a 95% confidence interval.
RESULTS AND DISCUSSION
ACE inhibitory peptides can be produced after the hydrolysis process by the action of proteolytic enzymes produced by growing tempe fungi. The amount of inhibitory activity was influenced by the protein source, fermentation process and incubation time. The peptides derived from vegetables protein are preferred by the scientific community and vegetarians because they are more sustainable than animal protein28. In addition, vegetable protein meets the requirements of a healthy and balanced diet29. Jack bean is a vegetable protein source with an adequate essential amino acids composition16. The proximate and amino acid composition of jack bean seeds are shown in Table 1 and 2.
The protein content of jack bean is higher than the velvet bean30, lima bean31 and mung bean32. In a previous study, Murdiati et al.17 reported that the protein content of jack bean was 32.32%, slightly higher than detected in this study. This difference was likely caused by the variation of geographic locations, which may affect the content and quality of the jack bean seed protein16.
| Table 1: | Proximate legume composition |
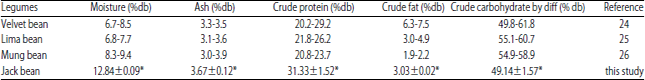 | |
| *Values are given as the Mean±SD of n = 2 | |
| Table 2: | Jack bean amino acid composition |
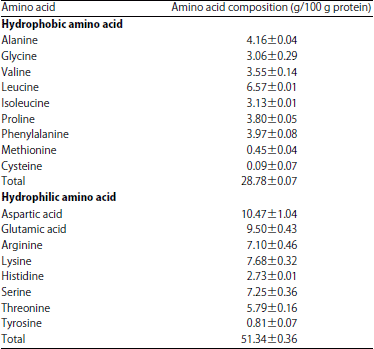 | |
| Data are the Mean±SD of n = 2 | |
Jack bean seeds have higher essential amino acid contents (especially leucine and isoleucine) than common legumes such as Vigna mungo and V. radiata16. The amino acid composition of jack bean seeds will affect the bioactive peptides released during tempe fermentation. Table 2 shows that glutamic acid and aspartic acid were the major amino acids in the jack bean that are similar to the soybean. Soybean has been previously investigated regarding ACE-inhibitory peptides8. High acidic amino acids could chelate the zinc atoms of ACE, which are needed for enzyme activation4. Leucine and proline were the most abundant hydrophobic amino acid in jack bean. Both types of the amino acids have an important role in ACE-inhibitory activity33. The protein sources, hydrolysis conditions, the degree of hydrolysis, molecular mass, amino acid composition and the position of amino acids in the peptide sequences influenced the ACE-inhibitory activity of the peptides and protein hydrolysates2,34.
Proteolytic activity: The main factor affecting protein hydrolysis during tempe fermentation was how much of the proteolytic enzyme was released into the seeds by the fungi. The protein hydrolysis products could be peptides, amino acids and the decomposition of substances28. Figure 1 shows that the first proteolytic activity was observed after 24 h of fermentation, which was equal to 3.30 unit g–1.
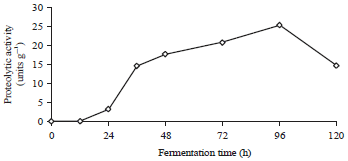 | |
| Fig. 1: | Proteolytic activity during tempe fermentation |
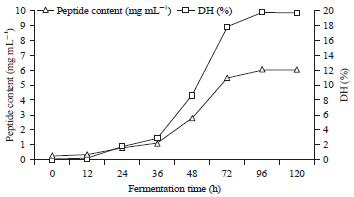 | |
| Fig. 2: | The degree of hydrolysis and peptide content during tempe fermentation |
Along with the production of proteases, the growth of fungal mycelia started to cover the surface of the seeds. In general, the tempe fermentation process from soybeans attained optimum conditions before 48 h of fermentation, as indicated by the compact mycelia covering the surface of the seed. However, the tempe fermentation from jack beans was somewhat different, as the proteolytic activity was still increasing after 96 h of incubation (25.39 units g–1). These results differed from those of other legumes, such as tempe from barley and grass pea, which achieved optimum conditions after 36 and 32 h, respectively35,36. These results indicate that a hard seed texture will slow the growth of mycelia, which results in slow protein hydrolysis. In Fig. 2 shows that high protease activity in line with high hydrolysis levels.
The degree of hydrolysis and peptide content: The results show that the degree of hydrolysis and peptide content increased with the length of fermentation time (Fig. 2).
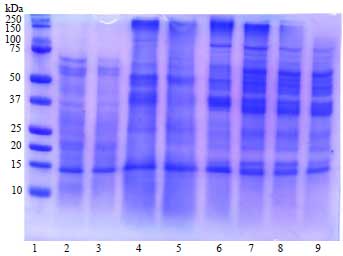 | |
| Fig. 3: | The protein pattern after different fermentation times |
| Lane 1: Marker, lane 2-9 jack bean fermented by a commercial tempe starter for 0, 12, 24, 36, 48, 72, 96 and 120 h, respectively | |
The peptide content before fermentation was 0.26 mg mL–1, increasing to 2.77 mg mL–1 after 48 h of fermentation and reaching a maximum to 6.05 mg mL–1 after 96 h. Likewise, a similar pattern was observed for the degree of hydrolysis of the protein, which increased with increasing incubation time and reached a maximum after 96 h fermentation (19.85%). Meanwhile, the degree of hydrolysis of the soybean fermented with Rhizopus oligosporus and Bacillus substillis was 20% after 48 h of fermentation37. From these data, we conclude that high protease activity will result in high the degree of hydrolysis values and therefore eventually, the peptide content will also be high. Figure 1 and 2 show this relationship.
The protein pattern: The protein pattern during tempe fermentation is described in Fig. 3. The protein bands with a molecular weight (MW) of 14.70 kDa were present at all stages of fermentation. At 0 and 12 h of fermentation, no pattern change is observed. Starting from 24 h of fermentation, new bands of proteins with lower MW appeared as a result of protein degradation by the proteases. These protein bands had an MW of 59.71, 27.32 and 16.70 kDa. These low molecular weight proteins had the potential to be an inhibitor of ACE38. In contrast, at the beginning of fermentation, there was no band with a high MW but after 24 h of fermentation a protein band with a high MW (109.69 kDa) was observed. This finding is likely due to the changes in protein solubility from insoluble before fermentation to soluble as a result of proteolytic hydrolysis.
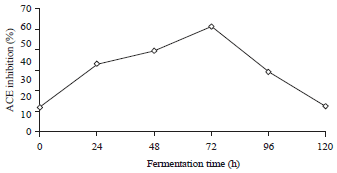 | |
| Fig. 4: | ACE inhibition during tempe fermentation |
ACE-inhibitory activity: Figure 4 shows that at 0 h of fermentation (unfermented jack bean), ACE inhibitory activity is present. This finding is probably due to the presence of small peptides with an MW of 14.78 kDa in the boiled beans (Fig. 3). The properties of bioactive peptides have been found in foods including natural substances and can also be produced through hydrolysis39. As shown in Fig. 4, the fermentation process increased the ACE inhibitory activity, reaching a maximum after 72 h of incubation (60.04% inhibition), after which the activity decreased. The ACE inhibition activity was influenced by the proteolytic activity released by Rhizopus oligosporus during tempe fermentation. As shown in Fig. 3, after 72 h of fermentation several lower MW protein bands appears near 16.70 kDa. These proteins have the possibility to contribute to ACE inhibitory activity.
The IC50 value of ACE-inhibitory activity after 72 h of fermentation was 1.03 mg mL–1. This IC50 value was lower than the IC50 values reported for fermented tomato (1.5 mg mL–1)9, miso (1.27 mg mL–1), tofu (1.77 mg mL–1) and soy sauce (3.44 mg mL–1) but was higher than natto (0.16 mg mL–1)40. This result reflects the capabilities of Rhizopus oligosporus proteases to hydrolyze jack bean proteins into bioactive peptides after various times of fermentation.
The results of this study suggest that the best time for jack bean tempe consumption is after 72 h of fermentation when the tempe has the highest ACE inhibitory activity, however, the fate of ACE-inhibitory peptides in the gastrointestinal tract has not been studied. Further work is required to demonstrate the ability of ACE inhibitory-peptides from jack bean tempe in antihypertensive animal model.
CONCLUSION
Jack bean is a legume with hard seeds that is a potential source of ACE inhibitor peptides. Fermentation into tempe takes longer than tempe from common beans, with an optimum time of 72 h. Under this condition, jack bean tempe produced high ACE inhibitory activity, as high as 60.04% (IC50 = 1.03 mg mL–1).
SIGNIFICANCE STATEMENT
This study reports the best incubation time of jack bean tempe fermentation for the production of high ACE-inhibitory peptides that can be beneficial for consumers of tempe, especially hypertensive people. This study will help the other researchers explore additional methods of hydrolysis of jack bean protein related to the ability to produce ACE inhibitory peptides. The potential finding of fermented jack bean tempe as a source of ACE inhibitory peptides has not been previously published and research on the absorption of these peptides is ongoing.
ACKNOWLEDGMENTS
The authors wish to acknowledge the financial support of the Doctoral Program from the Indonesia Endowment Fund for Education (Lembaga Pengelola Dana Pendidikan), The Ministry of Finance, Republic of Indonesia. The authors also thank The Ministry of Research, Technology and Higher Education, Republic of Indonesia for supporting this project through the Postgraduate Team Project (Proyek Tim Pasca) 2018.
REFERENCES
- Aluko, R.E., 2015. Antihypertensive peptides from food proteins. Annu. Rev. Food Sci. Technol., 6: 235-262.
CrossRefPubMedDirect Link - Daskaya-Dikmen, C., A. Yucetepe, F. Karbancioglu-Guler, H. Daskaya and B. Ozcelik, 2017. Angiotensin-I-Converting Enzyme (ACE)-inhibitory peptides from plants. Nutrients, Vol. 9, No. 4.
CrossRefDirect Link - Garcia, M.C., P. Puchalska, C. Esteve and M.L. Marina, 2013. Vegetable foods: A cheap source of proteins and peptides with antihypertensive, antioxidant and other less occurrence bioactivities. Talanta, 106: 328-349.
CrossRefPubMedDirect Link - Aluko, R.E., 2015. Structure and function of plant protein-derived antihypertensive peptides. Curr. Opin. Food Sci., 4: 44-50.
CrossRefDirect Link - Segura-Campos, M.R., C.P. Espadas-Alcocer, L. Chel-Guerrero and D. Betancur-Ancona, 2013. ACE-I inhibitory peptide fractions from enzymatic hydrolysates of velvet bean (Mucuna pruriens). Agric. Sci., 4: 767-773.
CrossRefDirect Link - Torruco-Uco, J., L. Chel-Guerrero, A. Martinez-Ayala, G. Davila-Ortiz and D. Betancur-Ancona, 2009. Angiotensin-I converting enzyme inhibitory and antioxidant activities of protein hydrolysates from Phaseolus lunatus and Phaseolus vulgaris seeds. Food Sci. Technol., 42: 1597-1604.
CrossRefDirect Link - Ibe, S., K. Yoshida, K. Kumada, S. Tsurushiin, T. Furusho and K. Otobe, 2009. Antihypertensive effects of natto, a traditional Japanese fermented food, in spontaneously hypertensive rats. Food Sci. Technol. Res., 15: 199-202.
CrossRefDirect Link - Gibbs, B.F., A. Zougman, R. Masse and C. Mulligan, 2004. Production and characterization of bioactive peptides from soy hydrolysate and soy-fermented food. Food Res. Int., 37: 123-131.
CrossRefDirect Link - Moayedi, A., L. Mora, M.C. Aristoy, M. Hashemi, M. Safari and F. Toldra, 2017. ACE-inhibitory and antioxidant activities of peptide fragments obtained from tomato processing by-products fermented using Bacillus subtilis: Effect of amino acid composition and peptides molecular mass distribution. Applied Biochem. Biotechnol., 181: 48-64.
CrossRefDirect Link - Wenno, M.R., E. Suprayitno, Aulanni’am and Hardoko, 2016. The physicochemical characteristics and Angiotensin Converting Enzyme (ACE) inhibitory activity of skipjack tuna (Katsuwonus pelamis) “Bakasang”. J. Teknol., 2: 119-124.
CrossRefDirect Link - Wikandari, P.R. and L. Yuanita, 2016. Pengaruh degradasi enzim proteolitik terhadap aktivitas angiotensin converting enzyme inhibitor bekasam dengan Lactobacillus plantarum B1765. Agritech, 36: 170-175.
CrossRefDirect Link - De Reu, J.C., R.M. ten Wolde, J. de Groot, M.R. Nout, F.M. Rombouts and H. Gruppen, 1995. Protein hydrolysis during soybean tempe fermentation with Rhizopus oligosporus. J. Agric. Food Chem., 43: 2235-2239.
CrossRefDirect Link - Sukasih, E. and E.Y. Purwani, 2017. Introductory study on processing of fermented jack bean (Canavalia ensiformis). Indonesian J. Agric. Postharvest Res., 12: 129-136.
Direct Link - Sridha, K.R. and S. Seena, 2006. Nutritional and antinutritional significance of four unconventional legumes of the genus Canavalia-A comparative study. Food Chem., 99: 267-288.
CrossRefDirect Link - Murdiati, A., S. Anggrahini, Supriyanto and A. Alim, 2015. Increased protein content of wet noodle from tapioca substituted by white jack bean (Canavalia ensiformis L.) flour. Agritech-J. Teknol. Pertanian, 35: 251-260.
CrossRef - Elegado, F.B. and Y. Fujio, 1993. Growth of Rhizopus strains on soybean and their protease formation. Bull. Fac. Agric. Kyushu Univ., 37: 315-324.
Direct Link - Cushman, D.W. and H.S. Cheung, 1971. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol., 20: 1637-1648.
CrossRefPubMedDirect Link - Church, F.C., H.E. Swaisgood, D.H. Porter and G.L. Catignani, 1983. Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins. J. Dairy Sci., 66: 1219-1227.
CrossRefDirect Link - Wulandari, R.D., E. Rahayu, Y. Marsono and T. Utami, 2017. [Antioxidant activity and angiotensin-I converting enzyme inhibitor of yogurt with ficus glomerata roxb leaf extract]. Agritech, 37: 246-255.
CrossRefDirect Link - Lin, H.C., A.M. Alashi, R.E. Aluko, B.S. Pan and Y.W. Chang, 2017. Antihypertensive properties of tilapia (Oreochromis spp.) frame and skin enzymatic protein hydrolysates. Food Nutr. Res., Vol. 61, No. 1.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Singh, B.P., S. Vij and S. Hati, 2014. Functional significance of bioactive peptides derived from soybean. Peptides, 54: 171-179.
CrossRefDirect Link - Duranti, M., 2006. Grain legume proteins and nutraceutical properties. Fitoterapia, 77: 67-82.
CrossRefDirect Link - Vadivel, V. and K. Janardhanan, 2000. Nutritional and anti-nutritional composition of velvet bean: An under-utilized food legume in South India. Int. J. Food Sci. Nutr., 51: 279-287.
CrossRefPubMedDirect Link - Oshodi, A.A. and M.O.A. Adeladun, 1993. Proximate composition, some nutritionally valuable minerals and functional properties of three varieties of Lima bean (Phaseolus lunatus Linn.) flour. Int. J. Food Sci. Nutr., 43: 181-186.
CrossRefDirect Link - Habibullah, M. Abbas and H.U. Shah, 2007. Proximate and mineral composition of mung bean. Sarhad J. Agric., 23: 463-466.
Direct Link - Hang, M. and X.H. Zhao, 2012. Fermentation time and ethanol/water-based solvent system impacted in vitro ACE-inhibitory activity of the extract of Mao-tofu fermented by Mucor spp. CyTA-J. Food, 10: 137-143.
CrossRefDirect Link - Ni, H., L. Li, G. Liu and S.Q. Hu, 2012. Inhibition mechanism and model of an angiotensin I-converting enzyme (ACE)-inhibitory hexapeptide from yeast (Saccharomyces cerevisiae). PloS One, Vol. 7, No. 5.
CrossRefDirect Link - Sher, M.G., M. Nadeem and Q. Syed, 2011. Study on protease from barley tempeh and in vitro protein digestibility. Jordan J. Biol. Sci., 4: 257-264.
Direct Link - Starzynska-Janiszewska, A., B. Stodolak and A. Wikiera, 2015. Proteolysis in tempeh-type products obtained with Rhizopus and Aspergillus strains from grass pea (Lathyrus sativus) seeds. Acta Sci. Pol. Technol. Aliment., 14: 125-132.
PubMedDirect Link - Weng, T.M. and M.T. Chen, 2011. Effect of two-step fermentation by Rhizopus oligosporus and Bacillus subtilis on protein of fermented soybean. Food Sci. Technol. Res., 17: 393-400.
CrossRefDirect Link - Malaguti, M., G. Dinelli, E. Leoncini, V. Bregola, S. Bosi, A. Cicero and S. Hrelia, 2014. Bioactive peptides in cereals and legumes: Agronomical, biochemical and clinical aspects. Int. J. Mol. Sci., 15: 21120-21135.
CrossRefDirect Link - Kuba, M., K. Tanaka, S. Tawata, Y. Takeda and M. Yasuda, 2003. Angiotensin I-converting enzyme inhibitory peptides isolated from tofuyo fermented soybean food. Biosci. Biotechnol. Biochem., 67: 1278-1283.
CrossRefPubMedDirect Link