
Thatsanee Anukoolprasert
Department of Fisheries, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
Khajornkiat Srinuansom
Department of Fisheries, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
Thanasorn Rukdontri
Department of Fisheries, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
Sataphon Nonkhukhetkhong
Department of Fisheries, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
Rakpong Petkam
Department of Fisheries, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
Pakistan Journal of Nutrition
Year: 2019 | Volume: 18 | Issue: 2 | Page No.: 125-133







ABSTRACT
Background and Objective: Protease-specific activity varies postprandially. This study aimed to identify the optimum time after feeding Nile tilapia (Oreochromis niloticus L.) to collect digestive organ samples (stomach, proximal intestine, distal intestine and liver) to obtain maximum protease-specific activity values. Materials and Methods: One hundred Nile tilapia (average weight 55.19±1.78 g/fish) were acclimatized and fed two times per day for 14 days. At the beginning of the experiment, 6 fish were randomly taken from the tank before feeding (0 h) and then 6 fish were also randomly taken after feeding at 1, 3, 6, 12, 18, 24, 36 and 48 h for digestive organ collection. Homogenized organ pH was measured and extracted for a protease-specific activity assay using azocasein as a substrate. Results: The pH values of all organs differed (p<0.05) after feeding and the pH changes were related to chyme movement in these organs. The protease-specific activities of the stomach and proximal intestine were highest at 24 h after feeding (p<0.05) but the protease-specific activities of the distal intestine and liver were not significantly different (p>0.05) between time points. Conclusion: Postprandial digestive organ sample collection was recommended at 24 h to maximize protease-specific activity for further protease characterization.
PDF Abstract XML References Citation
Received: June 27, 2018;
Accepted: October 12, 2018;
Published: January 15, 2019
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Thatsanee Anukoolprasert, Khajornkiat Srinuansom, Thanasorn Rukdontri, Sataphon Nonkhukhetkhong and Rakpong Petkam, 2019. Postprandial In vitro Protease-Specific Activity of Nile Tilapia (Oreochromis niloticus L.) Digestive Organs. Pakistan Journal of Nutrition, 18: 125-133.
DOI: 10.3923/pjn.2019.125.133
URL: https://scialert.net/abstract/?doi=pjn.2019.125.133
DOI: 10.3923/pjn.2019.125.133
URL: https://scialert.net/abstract/?doi=pjn.2019.125.133
INTRODUCTION
Nile tilapia (O. niloticus L.) has a strong economic impact in many countries1-3. The global production trends of Nile tilapia in aquaculture have been increasing and exceeded 5.45 million tons in 20143. In aquaculture production, feed accounts for 50-70% of production costs4,5, while dietary protein sources are the most expensive feed ingredients6-10.
Protein digestion and utilization are important for fish growth. Many studies on proteolytic enzymes in fish contribute to a better understanding of them and help improve protein utilization for fish production11-31. Proteolytic enzymes, known as proteases or proteinases, catalyze the hydrolysis of proteins to peptides and amino acids commonly found in organs such as the stomach, intestine and liver32.
Many factors affect proteolytic activities include species, dietary composition, developmental stages, bacterial enzyme activities, temperature and intestinal pH13,33-39. In an earlier study, digestive organs of Nile tilapia (O. niloticus L.) with body weights of 5.70, 35.80 and 92.10 g were collected at 16 h after feeding and revealed different protease activities among the three size groups15. Uscanga et al.40 reported varied protease activity at different postprandial times (0, 30, 60 and 90 min) of O. niloticus L. (50.30 g). These results suggest that sampling time is an important factor affecting the activity of enzymes. The present study aimed to investigate the effect of time after feeding on the protease-specific activity of Nile tilapia (O. niloticus L.) from digestive organs including the stomach, proximal intestine, distal intestine and liver.
MATERIALS AND METHODS
Fish and preparation of samples: One hundred Nile tilapia (O. niloticus L.) with similar size (body weight 55.19±1.78 g/fish) obtained from a local fish farm in Khon Kaen province, Thailand, were used in this study. They were acclimatized for 2 weeks prior to the experiment in a round plastic tank with a diameter of 2.00 m containing 2.50 m3 of water. Fish were fed at 06.00 and 18.00 to near satiation using pelletized commercial feed containing 32% crude protein. The experiment used a completely randomized design (CRD). On day 15, fish were randomly taken in 6 fish groups at time points before feeding (0 h) and at 1, 3, 6, 12, 18, 24, 36 and 48 h after feeding. Body weight and standard length of the fish were recorded. Then, fish were euthanized by submersion in ice cold water and the stomach, proximal intestine, distal intestine and liver were dissected out on an ice-chilled tray. The samples were washed with distilled water and weighed and pooled samples of 2 fish and 3 replicates were kept in liquid nitrogen for further analysis. The weights of the stomach, intestine, liver and fish body were calculated as follows:
 |
Digestive organ pH measurement: Digestive organs were homogenized by a tissue grinder (Wheaton, Potter ELV) on ice and then mixed with distilled water 1: 5 (g: mL). The pH of the homogenates was measured using a calibrated pH meter (Denver instrument, Ultrabasic). The mixtures were kept on ice at 4°C prior to a determination of protease-specific activity.
Determination of protease-specific activity: The homogenized sample was mixed with 1 M Tris-HCl buffer pH 7.00 (1:0.05 v/v) and then centrifuged (Centurion Scientific Ltd., K240R) at 15,000×g and 4°C for 60 min. The supernatant was collected and kept in liquid nitrogen for further determination of the enzyme-specific activity of protease (modified from Areekijseree et al.26). The protein concentrations of crude enzymes from the stomach, proximal intestine, distal intestine and liver were determined by following Lowry’s method (1951) using bovine serum albumin as the standard.
Protease-specific activity was analyzed by measuring the increase of oligopeptides, a short chain polypeptide26. The activity was determined by using 250 μL of 0.5% azocasein as a substrate and adding 10 μL of crude enzyme. Substrate was dissolved in KCl-HCl buffer at an acidic pH of 2.00 and NaHCO3-Na2CO3 buffer at an alkaline pH of 9.00. The mixture was incubated at 30°C for 30 min and the reaction was stopped with 1.20 mL of 10% Trichloroacetic acid (TCA). Blank was prepared by mixing with crude enzyme and stopping activity with TCA followed by adding substrate. The mixture was centrifuged at 10,000×g for 15 min at 4°C and the collected supernatant (1.20 mL) was mixed with 1.40 mL of 1.0 M NaOH. Total protease specific activity was determined as U mg–1 protein/min. One unit (U) of protease activity was specified as an increase in absorbance at 440 nm per minute at the specific reaction condition, as measured by a spectrophotometer (Biochrom, Libra S80).
Statistical analysis: Statistical analysis of data was performed using one-way analysis of variance (one-way ANOVA). Duncan's multiple range test at p<0.05 was used when a significant difference existed. A correlation between postprandial time and biometric parameters of fish was calculated.
RESULTS
Postprandial physical characteristics of fish: Body weight, total length and hepatosomatic index (HSI) of Nile tilapia (O. niloticus L.) at the time of sample collection are shown in Table 1. There were no significant differences among treatments (p>0.05). The average body weight at all periods was 55.19±1.78 g/fish and the total length was 14.59±0.14 cm/fish. The mean HSI was 2.66±0.12. Stomach weight, intestine weight, relative stomach weight, relative intestine weight and visceral somatic index (VSI) were significantly different between time period groups (p<0.05). In general, both stomach and intestine weights were significantly decreased (p<0.01) after feeding. The highest stomach weight was in the 1st h after feeding (3.16±1.16 g/fish) and the lowest weight was at 48 h (0.69±0.17 g/fish). The average intestine weight was higher than 2.00 g until 6 h and lower onward. The average relative stomach weight was 0.03±0.00 g g–1 body weight before feeding and at 6 h after feeding and decreased as time increased. The relative intestine weights were lowest at 36 h and highest before feeding at 0.02 and 0.05 g g–1 body weight, respectively. The vesicle somatic index (VSI) was calculated from the sum of stomach, intestine, liver and other entrails per total body weight of fish. VSI was highest at 6 h after feeding (11.24±0.45) and then decreased as time increased (p<0.05).
Postprandial pH changes: The pH values of the stomach, proximal Intestine, distal intestine and liver of Nile tilapia (O. niloticus L.) are shown in Fig. 1. The stomach pH was between 4.18 and 6.61 (Fig. 1a). The lowest stomach pH was 4.18 ± 0.12 before feeding and significantly differed from the others (p<0.05). The proximal intestine pH ranged from 6.89-7.53 (Fig. 1b) and significantly differed (p<0.05) among collection time points. The distal intestine pH ranged from 6.95-7.97 (Fig. 1c).
| Table 1: | Alteration in body weight, stomach weight, intestine weight, vesicle somatic index (VSI) and hepatosomatic index (HSI) of Nile tilapia during the period after feeding |
 | |
| Data represented as Mean±SEM, a,b,cDifferent superscript letters indicate a significant difference between values on a given row (p<0.05) | |
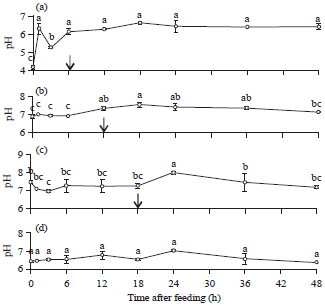 | |
| Fig. 1(a-d): | Postprandial changes in the pH of the (a) Stomach, (b) Proximal intestine, (c) Distal intestine and (d) Liver of Nile tilapia |
| Each point represents the mean of 3 replicates±SEM. Different letters indicate a significant difference between values on a point of time after feeding (p<0.05). ↓: Indicates the chyme exhausted from the organ | |
The highest pH in the distal intestine was at 24 h after feeding (7.97±0.05) and was significantly different from other collection time points (p<0.05). The pH of the liver ranged from 6.35-7.01 (Fig. 1d), which was not significantly different (p>0.05) among collection time points in the experiment. The lowest pH (6.35±0.15) of the liver was found at 48 h after feeding. At 24 h after feeding, the liver had the highest measured pH, which was 7.01±0.22.
Postprandial changes of In vitro protease-specific activity: Postprandial In vitro protease-specific activity of the stomach, proximal intestine, distal intestine and liver extracts are shown in Fig. 2. The alkaline protease activities of the stomach are shown as solid lines in Fig. 2a. The highest alkaline protease activity of the stomach was found at 24 h postprandial (0.18±0.01 U mg–1 protein/min) and was significantly different (p<0.05) from other collection times. The acidic protease activity of the stomach was the highest at before feeding (0 h), shown in Fig. 2a (dash line). For protease-specific activities in the proximal intestine, a peak of alkaline protease activity was found before feeding (0.23±0.02 U mg–1 protein/min) and it increased again after 24 h (0.18±0.05 U mg–1 protein/min) (Fig. 2b). The protease-specific activities of the distal intestine were approximately 0.02-0.39 U mg–1 protein/min (Fig. 2c) and were not significantly different (p>0.05).
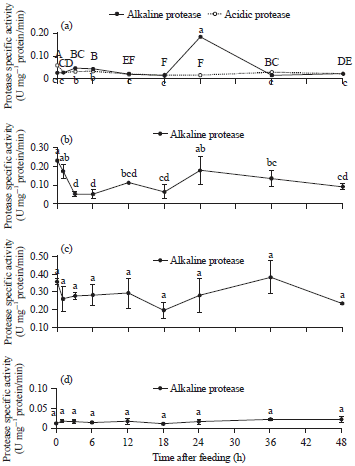 | |
| Fig. 2 (a-d): | Postprandial changes in protease-specific activity of the (a) Stomach, (b) Proximal intestine, (c) Distal intestine and (d) liver of Nile tilapia |
| The data points represent the mean of 3 replicates±SEM. Different letters indicate a significant difference between values at a point of time after feeding (p<0.05) | |
The lowest protease specific activity was 18 h postprandial (0.20±0.05 U mg–1 protein/min), while the highest protease specific activity was at 36 h (0.39±0.09 U mg–1 protein/min). The alkaline protease activities from liver extracts were 0.01-0.02 U mg–1 protein/min in the present study, which were the lowest compared to other organs (Fig. 2d) and there were no significant differences (p>0.05) in protease-specific activities measured in liver extracts from all time points.
DISCUSSION
In the present study, with a short experimental period (48 h), there were no significant differences in O. niloticus L. weight, which was similar to an earlier report in which fish fed and starved for 3 days were not significantly different41. However, the results in our study contrasted those of an earlier study with a 7-day starvation period in which Danio rerio weight, after 7-day starvation, was slightly higher than the fed fish42. Similarly, the weight of Oncorhynchus mykiss decreased when starved for 4 weeks43.
The HSI of O. niloticus L. did not change when starved for 48 h (Table 1). This was in agreement with another study which showed the liver weight was stable until 9 days of starvation in Pagrus major 44. In general, HSI provides an indication of the status of energy reserves and health of the animal because it shrinks under adverse environmental conditions45. The correlation coefficient between collecting time and physical characteristics (weight of stomach, intestine and VSI) showed a tendency to decrease, r = -0.293, -0.701 and -0.502, respectively. These results indicated that the weight of digestive organs decreased postprandially. Changes in body weight and HSI from all collection times were not observed following 48 h starvation, which was due to the short experimental period, unlike stomach and intestine weight, where changes after feeding were most likely observed due to chyme movement.
The stomach pH was significantly different (p<0.05). The stomach pH was lowest before feeding. Although postprandial stomach pH (1 through 6 h) increased from 6 h until 48 h, pH was relatively stable from 6.13-6.61 (p>0.05) on an empty stomach. The stomach pH increased when there was no chyme. This was in agreement with other studies on O. niloticus L.46 and Alcolapia graham 47 which showed that pH was higher in fish with empty stomachs and was lower in fish with full stomachs. A lower pH condition in the stomach was caused by the secretion of hydrochloric acid (HCl)48,49. The results of this study and earlier reports supported the stomach pH changes in which the pH of a full stomach was acidic47,50,51. The pH of the intestine displayed a trend of increasing alkalinity as time increased. The proximal intestine had a low pH (6.89-6.92) at 0 through 6 h after feeding, which was probably caused by the movement of chyme from the stomach into the proximal intestine. The pH was increased to 7.34±0.11 at 12 h and was highest at 18 h (7.53±0.13). The proximal intestine was empty at 12 h postprandial. This was similar to earlier reports in that the proximal intestinal pH was neutral and remained unchanged after feeding. In other species (T. rendalliin , O. mossambicus, C. gariepinus and A. graham) their proximal intestine pH was 7-838,48. The distal intestine pH had a statistically significant difference (p<0.05) between the average intestinal pH of the collection time. At 18 h after feeding, the chyme was not visible in the distal intestine. The distal intestine pH increased after the distal intestine was empty at 24 h after feeding, then pH was maintained from 7.17-7.43. Similarly, A. graham had an average hindgut pH of 8.1247. In the distal intestine, pH values of S. aurata were increased from 6.90-7.80 at 6 h47. In the distal intestine, the pH of S. aurata increased from 6.90-7.80 at 6 h after feeding34. In the distal intestine of T. rendalli, O. mossambicus and C. gariepinus, the pH values were in the alkaline range at all experiment time points38. In all intestinal sections of O. mykiss, alkaline pH values were recorded over time following feeding (0 through 72 h) and by 48 h, pH values returned to similar conditions before ingestion of a meal49. The highest pH values of the proximal and distal intestines in S. aurata were alkaline (pH>7.00) at 12 h34. The pH variations found in the different parts of the digestive organs could be explained by the flow of chyme along the digestive tract34,49. Secretion of bile (bicarbonate) also contributed to the alkaline pH of the proximal and distal intestines36,52. In addition, our results showed that the movement of chyme affected the change in pH over time in the digestive organs but not in the liver. In terms of chyme movement, chyme was observed in the stomach for 6 h, then moved to the proximal intestine and remained in both the proximal intestine and distal intestine for 6 h. Eventually, no feed was observed in the digestive tract at 18 h after feeding.
In terms of protease activity, the highest acidic protease activity in the stomach was found before feeding, when pH was the lowest. Similar findings were reported by Simpson53 and Zhao et al.54 who observed that the acidic proteases from fish stomachs, pepsin displayed high activity between pH 2.0 and 4.0. Similarly, the highest pH for acidic protease activity was 2.5 and 1.6-3.3 in O. nilotica and hybrid juvenile tilapia (O. niloticus×O. aureus) stomachs, respectively55,56. In other species, the highest protease specific activity in the stomach of Symphysodon aequifasciatus 25, S. aurata and Dentex dentex 57 were found at pH 2 and at pH 1.8 in Colossoma macropomum 58. In general, the enzyme activities changed when the pH of the stomach changed18. The stomach protease-specific activities of O. mossambicus, C. gariepinus and T. rendalli were the highest at 12, 12 and 31 h, respectively, after feeding38. Another experiment on S. aurata reported acid protease activity between pH 3 and 4.5, which was high at 0 h and 3 h, respectively, after a meal18. On the other hand, the alkaline protease activity in the stomach of S. aurata was highest at 1 h after feeding but the acid protease activities did not differ from other times after feeding35. The activity of alkaline protease from the stomach was highest at pH 8-9 in O. niloticus L. (5.7, 35.8 and 92.1 g)15. In the stomach and intestine of S. dumerilii, the values of protease-specific activities at 6 h after feeding were similar to those measured in an empty stomach (after 2 days of starvation, which was 48 h after feeding)13. In all in vitro protease-specific activities of the stomach, it was found to be lower than that of the intestine and this was in agreement with other studies conducted on O. niloticus L.15,59, T. rendalli 38, Pagellus bogaraveo, Seriola dumerilii 13, C. gariepinus 38 and Scleropages formosus 28.
The proximal intestine protease-specific activities were decreased in low pH before feeding to 6 h after feeding. Then, specific activities of the protease in all intestines of O. niloticus L. were highest when chyme moved from the stomach into the proximal intestines at 6 h after stomach emptiness. This was in agreement with the estimated total gut evacuation of O. niloticus L. reported in an earlier study at 7.15 h40. The highest activity of the alkaline protease in the proximal intestine of O. mykiss was 6 h after stomach emptiness49. The activity of alkaline protease in the duodenum of S. aurata was higher from 0-2 h after feeding but not significantly different from other times18. The in vitro protease-specific activities of the intestine were higher when the pH increased. This was in agreement with earlier reports in which protease-specific intestinal activities were high when the pH was neutral or alkaline15,22,25,53,60. Based on the results of this study, the protease-specific activities of the distal intestine were higher than those of the proximal intestine. In contrast, the protease-specific activity of the proximal intestine of O. mykiss 49 and O. niloticus L.40 was higher than that of the distal intestine over 48 h after feeding.
The alkaline protease activities from liver extract in the present study were the lowest compared to the other organs, which was similar to earlier reports15,22,28. In addition, liver enzymes have inactive forms known as zymogens for preventing self-digestion32.
The time after feeding was a key factor in studying the enzyme activities because feeds are involved in the secretion of enzymes35,38,59. Protease-specific activity in the digestive tract of Rutilus rutilus caspicus was decreased after starvation for 1-3 weeks61. Stomach, proximal intestine and distal intestine of T. rendalli, O. mossambicus and C. gariepinus revealed the highest enzyme activity at 12 h after feeding38. Differences in gut evacuation may be due to species, dietary composition and chyme movement in the digestive tract, which are also factors affecting the activity of enzymes13,33,35,38.
Variations in pH and protease specific activity in the digestive system appeared to be affected by the movement of chyme at different times after feeding18,33,38. Other factors, including species, developmental stages, temperature and dietary composition, could also affect the pH and protease-specific activity changes13,33,35,38,39. In this study, a similar size of Nile tilapia fed the same formula diet was used; thus, species and dietary composition effects were eliminated.
The digestive system was empty at 18 h and the specific activities of the protease in all organs showed an increment from 18 h to their highest at 24 h, suggesting it was the best period after feeding for digestive tract sample collection to obtain the possible highest enzyme activity for further needed enzyme assays.
CONCLUSION
Starvation for 48 h did not cause a significant effect on body weight, length, or HSI. The movement of chyme at different times after feeding affected changes in the pH values of the digestive system. The highest protease-specific activities of digestive organs were found at 24 h after feeding in this study and thus it is recommended as the optimum time for digestive organ sample collection in further investigations on protease characterization in Nile tilapia (Oreochromis niloticus L.).
SIGNIFICANCE STATEMENT
This study discovers the time after feeding which effect on pH and protease-specific activity changes that can be beneficial for precise In vitro protein digestion of Nile tilapia. This study will help researchers uncover the critical area of the In vitro protease-specific activity in digestive organs in fish that are essential for an effective feed formulation.
ACKNOWLEDGMENTS
The authors wish to express our sincere thanks to the National Research Council of Thailand and Graduate School of Khon Kaen University, Thailand, for funding and to the Department of Fisheries, Faculty of Agriculture, Khon Kaen University, for use of the experimental facilities.
REFERENCES
- Gupta, M.V. and B.O. Acosta, 2004. A review of global tilapia farming practices. Aquacult. Asia, 9: 7-12.
Direct Link - Ansah, Y., E. Frimpong and E. Hallerman, 2014. Genetically-improved Tilapia strains in Africa: Potential benefits and negative impacts. Sustainability, 6: 3697-3721.
CrossRefDirect Link - Abdel-Tawwab, M., M.H. Ahmad, Y.A.E. Khattab and A.M.E. Shalaby, 2010. Effect of dietary protein level, initial body weight and their interaction on the growth, feed utilization and physiological alterations of Nile tilapia, Oreochromis niloticus (L.). Aquaculture, 298: 267-274.
CrossRefDirect Link - Mohanta, K.N., S. Subramanian and V.S. Korikanthimath, 2013. Evaluation of different animal protein sources in formulating the diets for blue gourami, Trichogaster trichopterus fingerlings. J. Aquacult. Res. Dev., Vol. 4.
CrossRefDirect Link - Pereira, R., L.M.P. Valente, I. Sousa-Pinto and P. Rema, 2012. Apparent nutrient digestibility of seaweeds by rainbow trout (Oncorhynchus mykiss) and Nile tilapia (Oreochromis niloticus). Algal Res., 1: 77-82.
CrossRefDirect Link - Alarcon, F.J., M. Diaz and F.J. Moyano, 1997. Studies on digestive enzymes in fish: Characterization and practical applications. Cah Options Mediterraneennes, 22: 113-121.
Direct Link - Caruso, G., M.G. Denaro and L. Genovese, 2009. Digestive enzymes in some teleost species of interest for mediterranean aquaculture. Open Fish Sci. J., 2: 74-86.
CrossRefDirect Link - Clements, K.D., D. Raubenheimer and J.H. Choat, 2009. Nutritional ecology of marine herbivorous fishes: Ten years on. Funct. Ecol., 23: 79-92.
CrossRefDirect Link - Klahan, R., N. Areechon, R. Yoonpundh and A. Engkagul, 2009. Characterization and activity of digestive enzymes in different sizes of Nile tilapia (Oreochromis niloticus L.). Kasetsart J. Nat. Sci., 43: 143-153.
Direct Link - Sultana, Z., S. Ahmed, S. Iqball and A.H. Chisty, 2010. Determination of in vitro protein digestibility of different feed ingredients for Nilotica (Oreochromis nilotica). Bangladesh Res. Public. J., 4: 87-94.
Direct Link - Lin, S. and L. Luo, 2011. Effects of different levels of soybean meal inclusion in replacement for fish meal on growth, digestive enzymes and transaminase activities in practical diets for juvenile tilapia, Oreochromis niloticus × O. aureus. Anim. Feed Sci. Technol., 168: 80-87.
CrossRefDirect Link - Marquez, L., R. Robles, G.A. Morales and F.J. Moyano, 2012. Gut pH as a limiting factor for digestive proteolysis in cultured juveniles of the gilthead sea bream (Sparus aurata). Fish Physiol. Biochem., 38: 859-869.
CrossRefDirect Link - Martinez‐Montano, E. and J.P. Lazo, 2012. In vitro protein digestibility of dietary ingredients throughout ontogeny of California halibut, Paralichthys californicus, larvae. J. World Aquacult. Soc., 43: 51-62.
CrossRefDirect Link - Thongprajukaew, K., U. Kovitvadhi, S. Kovitvadhi, A. Engkagul and K. Rungruangsak-Torrissen, 2013. Evaluation of growth performance and nutritional quality of diets using digestive enzyme markers and in vitro digestibility in Siamese fighting fish (Betta splendens Regan, 1910). Afr. J. Biotechnol., 12: 1689-1702.
Direct Link - Yasumaru, F. and D. Lemos, 2014. Species specific in vitro protein digestion (pH-stat) for fish: Method development and application for juvenile rainbow trout (Oncorhynchus mykiss), cobia (Rachycentron canadum) and Nile tilapia (Oreochromis niloticus). Aquaculture, 426: 74-84.
CrossRefDirect Link - Jan, M. and I. Ahmed, 2016. Assessment of fecundity, gonadosomatic index and hepatosomatic index of snow trout, Schizothorax plagiostomus in river Lidder, from Kashmir Himalaya, India. Int. J. Fish. Aquat. Stud., 4: 370-375.
Direct Link - El-Sayed, A.F., I. Nmartinez and F.J. Moyano, 2000. Assessment of the effect of plant inhibitors on digestive proteases of Nile tilapia using in vitro assays. Aquacult. Int., 8: 403-415.
CrossRefDirect Link - Pujante, I.M., M. Diaz-Lopez, J.M. Mancera and F.J. Moyano, 2017. Characterization of digestive enzymes protease and α-amylase activities in the thick-lipped grey mullet (Chelon labrosus, Risso 1827). Aquacult. Res., 48: 367-376.
CrossRefDirect Link - Chong, A.S.C., R. Hashim, L. Chow-Yang and A.B. Ali, 2002. Partial characterization and activities of proteases from the digestive tract of discus fish (Symphysodon aequifasciata). Aquaculture, 203: 321-333.
CrossRefDirect Link - Areekijseree, M., A. Engkagul, U. Kovitvadhi, A. Thongpan, M. Mingmuang, P. Pakkong and K. Rungruangsak-Torrissen, 2004. Temperature and pH characteristics of amylase and proteinase of adult freshwater pearl mussel, Hyriopsis (Hyriopsis) bialatus Simpson 1900. Aquaculture, 234: 575-587.
CrossRefDirect Link - Lemos, D., A.N. Del Toro, J.H. Cordova-Murueta and F. Garcia-Carreno, 2004. Testing feeds and feed ingredients for juvenile pink shrimp Farfantepenaeus paulensis: In vitro determination of protein digestibility and proteinase inhibition. Aquaculture, 239: 307-321.
CrossRefDirect Link - Natalia, Y., R. Hashim, A. Ali and A. Chong, 2004. Characterization of digestive enzymes in a carnivorous ornamental fish, the Asian bony tongue Scleropages formosus (Osteoglossidae). Aquaculture, 233: 305-320.
CrossRefDirect Link - Rungruangsak-Torrissen, K., R. Moss, L.H. Andresen, A. Berg and R. Waagbo, 2006. Different expressions of trypsin and chymotrypsin in relation to growth in Atlantic salmon (Salmo salar L.). Fish Physiol. Biochem., 32: 7-23.
CrossRefDirect Link - Infante, J.L.Z. and C.L. Cahu, 2007. Dietary modulation of some digestive enzymes and Metabolic processes in developing marine fish: Applications to diet formulation. Aquaculture, 268: 98-105.
CrossRefDirect Link - Ali, H., M.M. Haque, M.M.R. Chowdhury and M.I. Shariful, 2009. In vitro protein digestibility of different feed ingredients in Thai koi (Anabas testudineus). J. Bangladesh Agric. Univ., 7: 205-210.
CrossRefDirect Link - Britz, P.J., T. Hecht and J. Knauer, 1996. Gastric evacuation time and digestive enzyme activity in abalone Haliotis midae fed a formulated diet. S. Afr. J. Mar. Sci., 17: 297-303.
CrossRefDirect Link - Deguara, S., K. Jauncey and C. Agius, 2003. Enzyme activities and pH variations in the digestive tract of gilthead sea bream. J. Fish Biol., 62: 1033-1043.
CrossRefDirect Link - Montoya, A., J.F. Lopez-Olmeda, M. Yufera, M.J. Sanchez-Muros and F.J. Sanchez-Vazquez, 2010. Feeding time synchronises daily rhythms of behaviour and digestive physiology in Gilthead seabream (Sparus aurata). Aquaculture, 306: 315-321.
CrossRefDirect Link - Nikolopoulou, D., K.A. Moutou, E. Fountoulaki, B. Venou, S. Adamidou and M.N. Alexis, 2011. Patterns of gastric evacuation, digesta characteristics and pH changes along the gastrointestinal tract of gilthead sea bream (Sparus aurata L.) and European sea bass (Dicentrarchus labrax L.). Compa. Biochem. Physiol. Part A: Mol. Integr. Physiol., 158: 406-414.
CrossRefPubMedDirect Link - Santos, J.F., P.F. Castro, A.L.G. Leal, A.C.V. de Freitas Junior, D. Lemos, L.B. Carvalho Jr. and R.S. Bezerra, 2013. Digestive enzyme activity in juvenile Nile tilapia (Oreochromis niloticus, L.) submitted to different dietary levels of shrimp protein hydrolysate. Aquacult. Int., 21: 563-577.
CrossRefDirect Link - Hlophe, S.N., N.A.G. Moyo and I. Ncube, 2014. Postprandial changes in pH and enzyme activity from the stomach and intestines of Tilapia rendalli (Boulenger, 1897), Oreochromis mossambicus (Peters, 1852) and Clarias gariepinus (Burchell, 1822). J. Applied Ichthyol., 30: 35-41.
CrossRefDirect Link - Kuzmina, V.V., G.V. Zolotareva and V.A. Sheptitskiy, 2014. Effect of pH on the activity of proteinases in intestinal mucosa, chyme and microbiota of fish from the Cuciurgan reservoir. J. Ichthyol., 54: 591-597.
CrossRefDirect Link - Uscanga, A., F.J. Moyano and C.A. Alvarez, 2010. Assessment of enzymatic efficiency on protein digestion in the tilapia Oreochromis niloticus. Fish Physiol. Biochem., 36: 1079-1085.
CrossRefDirect Link - Takagi, Y., 2001. Effects of starvation and subsequent refeeding on formation and resorption of acellular bone in tilapia, Oreochromis niloticus. Zool. Sci., 18: 623-629.
CrossRefDirect Link - Drew, R.E., K.J. Rodnick, M. Settles, J. Wacyk and E. Churchill et al., 2008. Effect of starvation on transcriptomes of brain and liver in adult female zebrafish (Danio rerio). Physiol. Genomics, 35: 283-295.
CrossRefDirect Link - Sanchez-Muros, M.J., L. Garcia-Rejon, L. Garcia-Salguero, M. de la Higuera and J.A. Lupianez, 1998. Long-term nutritional effects on the primary liver and kidney metabolism in rainbow trout. Adaptive response to starvation and a high-protein, carbohydrate-free diet on glutamate dehydrogenase and alanine aminotransferase kinetics. Int. J. Biochem. Cell Biol., 30: 55-63.
CrossRefDirect Link - Sakamoto, S., M. Furuichi and Y. Yone, 1978. Effect of starvation on organ weight and chemical component of red sea bream. J. Fac. Agric. Kyushu Univ., 23: 71-77.
Direct Link - Ashwini, L., S. Benakappa, H.N. Anjanayappa and L. Akshay, 2016. Observation on the gonado-somatic index-GSI and hepato-somatic index-HSI of Decapterus russelli Mangaluru coast. Int. J. Eng. Sci. Comput., 6: 7396-7399.
Direct Link - Getachew, T., 1989. Stomach pH, feeding rhythm and ingestion rate in Oreochromis niloticus L. (Pisces: Cichlidae) in Lake Awasa, Ethiopia. Hydrobiologia, 174: 43-48.
CrossRefDirect Link - Bergman, A.N., P. Laurent, G. Otiang'a-Owiti, H.L. Bergman, P.J. Walsh, P. Wilson and C.M. Wood, 2003. Physiological adaptations of the gut in the Lake Magadi tilapia, Alcolapia graham, an alkaline-and saline-adapted teleost fish. Compa. Biochem. Physiol. Part A: Mol. Integr. Physiol., 136: 701-715.
CrossRefDirect Link - Papastamatiou, Y.P., S.J. Purkis and K.N. Holland, 2007. The response of gastric pH and motility to fasting and feeding in free swimming blacktip reef sharks, Carcharhinus melanopterus. J. Exp. Mar. Biol. Ecol., 345: 129-140.
CrossRefDirect Link - Bucking, C. and C.M. Wood, 2009. The effect of postprandial changes in pH along the gastrointestinal tract on the distribution of ions between the solid and fluid phases of chyme in rainbow trout. Aquacult. Nutr., 15: 282-296.
CrossRefDirect Link - Teferi, Y., D. Admassu and S. Mengistou, 2000. The food and feeding habit of Oreochromis niloticus L. (Pisces: Cichlidae) in lake Chamo, Ethiopia. Ethiop. J. Sci., 23: 1-12.
CrossRefDirect Link - Alemayehu, N. and G. Abebe, 2004. Food habits and diel feeding rhythm of introduced fish, T. zillii gervais, 1948 (Pisces: Cichlidae) in Lake Zwai, Ethiopia. Ethiop. J. Sci., 27: 9-16.
Direct Link - Zhao, J., Y. Liu, J. Jiang, P. Wu and G. Chen et al., 2012. Effects of dietary isoleucine on growth, the digestion and absorption capacity and gene expression in hepatopancreas and intestine of juvenile Jian carp (Cyprinus carpio var. Jian). Aquaculture, 368: 117-128.
CrossRefDirect Link - El-Beltagy, A.E., T.A. El-Adawy, E.H. Rahma and A.A. El-Bedawey, 2004. Purification and characterization of an acidic protease from the viscera of bolti fish (Tilapia nilotica). Food Chem., 86: 33-39.
CrossRefDirect Link - Jun-Sheng, L., L. Jian-Lin and W. Ting-Ting, 2006. Ontogeny of protease, amylase and lipase in the alimentary tract of hybrid juvenile tilapia (Oreochromis niloticus × Oreochromis aureus). Fish Physiol. Biochem., 23: 295-303.
CrossRefDirect Link - Alarcon, F.J., M. Diaz, F.J. Moyano and E. Abellan, 1998. Characterization and functional properties of digestive proteases in two sparids; gilthead seabream (Sparus aurata) and common dentex (Dentex dentex). Fish Physiol. Biochem., 19: 257-267.
CrossRefDirect Link - Bezerra, R.D.S., J.F.D. Santos, M.A.D.S. Lino, V.L.A. Vieira and L.B. Carvalho Jr., 2000. Characterization of stomach and pyloric caeca proteinases of tambaqui (Colossoma macropomum). J. Food Biochem., 24: 189-199.
CrossRefDirect Link - Vera, L.M., N. De Pedro, E. Gomez-Milan, M.J. Delgado, M.J. Sanchez-Muros, J.A. Madrid and F.J. Sanchez-Vazquez, 2007. Feeding entrainment of locomotor activity rhythms, digestive enzymes and neuroendocrine factors in goldfish. Physiol. Behav., 90: 518-524.
CrossRefDirect Link - Fagbenro, O., O. Adedire, O. Fateru, I. Oluwabukola and O. Ogunlana et al., 2005. Digestive enzyme assays in the gut of Oreochromis niloticus Linnaeus 1757, Parachanna (Channa) obscura Gunther 1861 and Gymnarchus niloticus Cuvier 1829. Anim. Res. Int., 2: 292-296.
CrossRefDirect Link - Abolfathi, M., A. Hajimoradloo, R. Ghorbani and A. Zamani, 2012. Effect of starvation and refeeding on digestive enzyme activities in juvenile roach, Rutilus rutilus caspicus. Compa. Biochem. Physiol. Part A: Mol. Integr. Physiol., 161: 166-173.
CrossRefDirect Link