
Nelson Elita
Study Program of Cultivation of Food Crops, Payakumbuh State Agricultural Polytechnic, Payakumbuh, West Sumatra, Indonesia
LiveDNA: 62.16570
Eka Susila
Study Program of Cultivation of Food Crops, Payakumbuh State Agricultural Polytechnic, Payakumbuh, West Sumatra, Indonesia
Yefri wati
Study Program of Cultivation of Food Crops, Payakumbuh State Agricultural Polytechnic, Payakumbuh, West Sumatra, Indonesia
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 12 | Page No.: 696-701







ABSTRACT
Background and Objective: The aerobic conditions during the vegetative phase of rice cultivation under the System of Rice Intensification (SRI)-method allow microorganisms, including Arbuscular Mycorrhizal Fungi (AMF), to be active. The AMF, which are derived from the rhizosphere of the plant itself (indigenous), are used because these fungi are adaptive and efficient. The compatibility between AMF and plant species is crucial for increasing nutrient uptake and plant growth. This research aims to; (1) Determine the effect of indigenous AMF types on the SRI rice production method and (2) Identify potential indigenous AMF types as sources of inoculum that can be used for rice cultivation under the SRI method. Materials and Methods: The materials used were 3 types of AMF isolates (Glomus sp2, Glomus sp3, Sclerocystis sp.). Greenhouse pot experiments with AMF treatments using a completely randomized included (1) no AMF (control), (2) Glomus sp2, (3) Glomus sp3 and (4) Sclerocystis sp. and were repeated 5 times. Growth production, root colonization and spore density were observed. Root colonization was observed in the Microbiology Laboratory of Payakumbuh State Agricultural Polytechnic. The similarity of variance between treatments was tested using Duncan’s Multiple Range Test (DMRT) at a 5% significance level. Results: Glomus sp3 treatment produced the greatest plant height, grain weight/panicle, 1000-seed weight and yield/hectare. Sclerocystis sp. treatment produced the greatest number of tillers, number of productive tillers and number of grains/panicle. The percentage of colonization and root infection intensity did not differ between Glomus sp3 and Sclerocystis sp. treatments. Conclusion: Glomus sp3 and Sclerocystis sp. are potential sources of inoculum that can be used in the SRI method of rice cultivation.
PDF Abstract XML References Citation
Received: October 10, 2017;
Accepted: August 29, 2018;
Published: November 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Nelson Elita, Eka Susila and Yefri wati, 2018. The Potential Types of Indigenous Arbuscular Mycorrhizal Fungi
as Sources of Inoculum and Their Effect on Rice Production Using
the System of Rice Intensification Method. Pakistan Journal of Nutrition, 17: 696-701.
DOI: 10.3923/pjn.2018.696.701
URL: https://scialert.net/abstract/?doi=pjn.2018.696.701
DOI: 10.3923/pjn.2018.696.701
URL: https://scialert.net/abstract/?doi=pjn.2018.696.701
INTRODUCTION
Infection of paddy fields in Indonesia is a serious problem. It is not easily addressed, because this infection is an accumulation culmination of various activities, such as continuous inorganic fertilization resulting in the accumulation of fertilizers, especially phosphorus, pesticides application and burning crop residues, which kill beneficial microorganisms. These activities lead to degradation of soil quality, causing the critical land area to continue to grow. Rice cultivation by the System of Rice Intensification (SRI) method uses an aerobic system during the vegetative phase, enabling microorganism activity and increasing microorganism availability. Uphoff1 states that aerobic conditions support soil microbes and their abundant diversity in soil through root exudates. The presence of rhizobia in the rhizosphere of rice plants increases protein levels and yield per hectare through the production of auxin and other aphrodisiacs. Root exudates are key factors in various parts of the SRI method and can result in larger rooting systems and more saplings.
Mycorrhizae form a symbiotic mutualism with high level plant roots. Soil hygiene can be quickly improved by exploiting mycorrhizae as bioremediators, because mycorrhizaes are more developed in unfavourable (marginal) environmental conditions2. The presence of mycorrhizae infection in the roots of rice plants can increase the variation and amount of nutrients, especially biological phosphorus, that can be absorbed by the roots, while also increasing the availability of soil phosphorus.
Indigenous mycorrhizae are more adaptive and develop quickly. Thus, their ability to absorb nutrients is greater, resulting in increased plant growth rate. According to Ruzicka et al.3 the success of mycorrhizal associations with plant roots is strongly influenced by the suitability of the AMF type to the host plant species. Nogueira and Cardoso4 and Tawaraya et al.5 suggest that differences in the behavior of each genus of Arbuscular Mycorrhizal Fungi (AMF) in different plants, affects plant growth and production.
The SRI method of rice cultivation with an aerobic system affects the quality of rice produced. The reduction of water requirements during the vegetative phase, means that the aerobic system during the vegetative phase increases the number of tillers to increase crop production6. Rice production by the SRI method reaches 9-10 t ha–1, while the intensification area managed by the SRI method reaches 20,040 ha, meaning that this methods can increase national production.
The AMF symbiosis with plants is very important and can be used for cultivation interests, especially when seedlings grow better in the field7. This symbiosis can increase soil nutrient uptake and root resistance to drought, maintain roots under disease attacks, supply additional growth hormone and growth regulators which enhance plant resistance to root pathogen attack8. Utilization of AMF has several advantages: the organisms used are available locally in nature and can be reproduced quickly with simple technology and their application is easy and relatively safe9,10.
Based on the above description, indigenous AMF is adaptive and efficient, environmentally friendly, economical and easily available. Indigenous AMF are believed to increase the production and quality of wetland soil intensification by the SRI method. This research aims to, (1) Determine the effect of indigenous AMF types on SRI rice production method and (2) Identify potential indigenous AMF types as sources of inoculum that can be used in the SRI method.
MATERIALS AND METHODS
Experimental design: The pot study was conducted in a greenhouse, using a completely randomized design. Three treatments were 3 types of AMF isolates (Glomus sp2, Glomus sp3, Sclerocystis sp.) and 1 treatment without AMF (control). All treatments in this experiment were replicated 5 times. Root colonization and spore density were observed at the Microbiology Laboratory of Payakumbuh State Agricultural Polytechnic. To examine the effect of treatments on the observed response, analysis of variance was performed using the Statistical Analysis System (SAS) program (Copyright© 2017, SAS Institute Inc., USA). Other brands and product names are trademarks of their respective companies. Duncan’s Multiple Range Test (DMRT) was then used to determine the difference between treatments at a 5% significance level.
Spore collection and execution: The AMF that were used were from spores derived from rice fields cultivated under the SRI method in Harau. Lima Puluh Kota District. Previous experimental results, revealed three types of indigenous AMF isolates (>50 spores/100 g), i.e., Glomus sp2, Glomus sp3 and Sclerocystis sp. These indigenous AMF were tested for their potential as sources of inoculum in the SRI method of rice cultivation (Fig. 1).
Material and equipment preparation: Prior to preliminary trials, preparations included: procurement of the materials and equipment needed for research, cleaning and sanitation of a greenhouse as the place of research and media preparation.
Land preparation: Media in the form of soil was derived from intensified wetland rice cultivated by the SRI method.
 | |
| Fig. 1(a-c): | (a) Glomus sp2, spore (globus), passed through a 63 μm screen, brownish yellow, no ornament, slick spore surface, (b) Glomus sp3, spore (oval), light brown, passed through a 250 μm screen, jagged spore wall, thick and (c) Sclerocystis sp., spore irregular (sub globus), passed through a 125 μm screen, dark orange brown, dirty and filled spores, non-slippery spore surface |
It was dried, sieved through a mesh size of 10 mesh and sterilized with the aim of killing all organisms in the soil samples; so that only the inoculated AMF would grow. Sterilization was performed by steaming (heating). Sterile soil samples were loaded into pots of 20 kg/bucket.
Inoculation and planting: Nursery paddies were previously formed on the seed bed. Twelve-day-old seedlings were moved to pots. The AMF were inoculated at planting. The AMF isolates were suitably treated and, contained approximately 70 spores each11. To each pot 15 g of previously standardized inoculum was added in the planting hole.
Fertilization: Half of the recommended doses of urea (150 kg ha–1 or 0.75 g/pot), SP36 (100 kg ha–1 or 0.50 g/pot) and KCl (50 kg ha–1 or 0.25 g/pot) were applied at planting and urea was applied 2 times (at planting and after 30 days) according to the procedure described by Larijani and Hoseini12.
Plant height (cm), number of panicles per hill, number of grains per panicle, 1000 seeds weight (g) and dry grain weight per hectare (ton) were observed in the greenhouse. Laboratory observations included root colonization and spore density. Spore density was calculated at the end of the observation, 10 g of AMF inoculum was added to a 500 mL glass cup along with 200 mL of sterile distilled water and the mixture was then stirred and left for 30 seconds. Spore suspensions were filtered throught mesh sizes of 500, 250, 100 and 50 μm. The remaining suspension was transferred to a centrifuge tube and 60% glucose was added; the tube was then centrifuged for 10 min at 1000 rpm. The spores in the centrifuge tubes were passed through a 50 μm sieve and carefully washed with water to remove glucose. The sponges retained on the sieve were transferred to a Petri dish and observed under a microscope according to the method described by Higo et al.8.
Root samples were collected from the rhizosphere of rice plants resulting after screening, to calculate root colonization, as follows:
| • | Percentage of AMF colonization on plant roots, calculated as described by Kormanik and McGraw13 |
| • | Infection intensity and classification of root infection rate were based on the classification of the Institute of Mycorrhizal Research and Development8: | |
| • | Class 1, when infection 0-5 % (very low, +) | |
| • | Class 2, when infection 6-25% (low, ++) | |
| • | Class 3, when infection 26-50% (medium, +++) | |
| • | Class 4, when infection 51-75% (high, ++++) | |
| • | Class 5, when the infection >75% (very high, +++++) | |
A higher class of infection intensity, indicates a higher intensity of AMF infections in plant roots and is one indicator in determining the effectiveness of AMF colonization in plants. In other words, the higher the colonization intensity between AMF and plants is, the more beneficial the symbiosis between the two.
RESULTS AND DISCUSSION
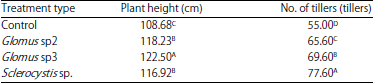
Vegetative growth: Table 1 shows that the Glomus sp3 treatment produced a significantly greater plant height than did the other AMF isolates. This indicates that Glomus sp3 can influence plant height in rice cultivated by the SRI method. The plant height in the Glomus sp2 treatment was not significantly different form that in the Sclerocystis sp. treatment but was significantly greater than that of the control.
The results show that inoculation with the 3 AMF isolates produced significantly different results compared to the control. The AMF can suppress nutrient loss from the soil by enlarging the zone of nutritional interception; this can also be achieved through washing with percolation water14. As reported by Yanai et al.15, AMF are associated with an increase in the effective area due to external hyphae extending up to 7-10 m g–1 soil. There was a difference in compatibility between the 3 AMF types, as seen in plant height and number of tillers. Glomus sp3 produced a greater plant height, than did Glomus sp2 and Sclerocystis sp. which produced the same height. Glomus sp3 and Sclerocystis sp. produced the most tillers.
Previous studies have reported that the compatibility of AMF isolates with host plants, determines AMF effectiveness in plants. As reported by Nogueira and Cardoso4 and Bertham16, Gigaspora margarita produced better growth and yield characteristics of soybean than did Glomus manihotis. This result suggests a difference in the behaviour of each AMF genus in infected plants.
| Table 1: | Vegetative characteristics of rice cultivated under the SRI method and treated with indigenous AMF isolates |
 | |
The numbers in the columns followed by the same upper case letter are not significantly different at a 5% significance level as determined by DNMRT | |
In-depth studies have attributed behavioural differences, such as those in nutrient uptake in the canopy, to differences in phosphatase activity among AMF genera growth. However, the results of this study indicate that Glomus sp3 and Sclerocystis sp. were the most suitable AMF isolates for rice growth under the SRI method.
Generative growth: All generative growth parameters showed that the AMF isolate treatments produced significantly different results from the control. As shown in Table 2, the number of productive tillers, number of grains/panicle and grain weight/panicle were significantly greater in the Sclerocystis sp. treatment, than in the other AMF isolate treatments. The 1000 seeds weight and yield/hectare were not significantly greater in the Sclerocystis sp. Treatment than in the Glomus sp3.
The external hyphae of AMF play a role in nutrient uptake. Janos et al.17 stated that the external hyphae of AMF helps bring nutrients into plant tissues, Suraya18 reported an increase in growth and nutrient uptake in teak plants. In addition hyphae play an important role in increasing the uptake of P and other nutrients, such as potassium, calcium, magnesium, iron, copper, manganese, zinc and boron19. For high growth and maximum yield, plants require some nutrients to be available throughout their life cycle, from planting to harvest20. AMF inoculation occurs once in the plant life cycle, producing an infection that will remain throughout. This means that nutrients will be available until the end of the cycle with the help of AMF.
Compared with the control treatment, all AMF isolate types positively influenced all generative growth parameters. According to Musfal21 increased dosing of AMF and NPK fertilizer improves yield by up to 40% compared to the control. Generative growth differed between treatments with AMF isolates and the control. Of the three types of AMF isolates applied, Glomus sp3 and Scleroscystis sp. treatment produced better results than did Glomus sp2 treatment. It is suspected that both spores have an average ability to remove external hyphae. This is indicated by the increase in all generative growth parameters compared to those under the Glomus sp2 treatment. These results suggest that these two types of indigenous AMF have the potential as sources of inoculum that can be used in the SRI method of rice cultivation.
| Table 2: | Generative growth of rice cultivated with the SRI method and treated with indigenous AMF isolates |
 | |
| The numbers in the columns followed by the same uppercase letter are not significantly different at a 5% significance level as determined by DNMRT | |
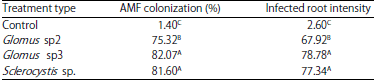
AMF infection test in the laboratory: Inoculation of plants with different types of AMF produced different infections intensities between treatments and between treatment and the control. This means that there are differences in the level of compatibility between the types of AMF inoculated with rice plants under the SRI method. The AMF colonization intensity was greatest in the Glomus sp3 treatment; this intensity did not significantly differ from that in the Sclerocystis sp. but was significantly greater than that in the Glomus sp2 treatment (Table 3). This result suggests that Glomus sp3 and Sclerocystis sp. are more effective in rice roots under SRI rice cultivation than is Glomus sp2. The AMF infection was still observed in the controls. This results demonstrates that some dormant AMF spores were still presents in the soil, even though the soil had been sterilized; the number of surviving spores should be small. According to Saputra et al.22 the AMF presence is strongly influenced by the area of roots distribution. The root distribution of plants under symbiosis with AMF is very wide, reaching 90%.
Root percentage was also correlated with root infection intensity. The effect of AMF inoculations significantly differed among AMF isolate types. Table 3 shows that the roots infection intensity by AMF in rice plants was very high (78.78) in the Glomus sp3 treatment, unlike the infection intensity in Sclerocystis sp. (77.34); both types of AMF spores are included in Class 5 (very high) of root infection intensity. In the Glomus sp2 treatment, the root infection intensity was 67.92, this spore is included in Class 4 (high). In the control treatment, the root infection intensity was in Class 1 (very low). This classification is based on The Institute of Mycorrhizal Research and Development, USDA Higo et al.8. The high rice root AMF infection intensity demonstrates that the vegetative growth and generative parameters were also high.
Rillig et al.23 suggested that inoculation with AMF may increase the root infection intensity by increasing the number of spores added to or formed around the plant, although high spore counts would not necessarily indicate higher roots infection intensity. In other words, AMF inoculation dose does not determine the roots infection in intensity. This value depends on the suitability of the AMF type to the host plant. Although the number of spores applied is small, if AMF adaptation to the host plant high, the spores will grow to high numbers. In this study, although the AMF inoculation doses were the same, there were differences in the infection intensity among the 3 types of AMF isolates. The results of this study, show that Glomus sp 3 and Sclerocystis sp. treatment were more effective in the roots of rice under the SRI cultivation method than was Glomus sp2 treatment.
| Table 3: | Percentage of AMF colonization and root infection intensity in rice cultivated with the SRI method and treated with different types of indigenous AMF isolate under SRI rice cultivation method |
 | |
The numbers in the columns followed by the same upper case letter are not significantly different at a 5% at a significance level as determined by DNMRT | |
| Table 4: | Observation of spore density of indigenous AMF isolates in rice cultivated under the SRI method |
 | |
The numbers in the columns followed by the same upper case letter are not significantly different at a 5% significance level as determined by DNMRT | |
Test of AMF spore density in the laboratory: The results of the variance analysis regarding spore density in rice cultivated under the SRI method are presented in Table 4.
Sclerocystis sp. produced the greatest spore density (89.80 spores). This value was not significantly different from that of Glomus sp3 (87.80 spores). This result indicates that Sclerocystis sp. and Glomus sp3 were better adapted than Glomus sp 2 to the roots of rice cultivated under the SRI method and thus produced the most spores. However, further research for SRI rice cultivation at locations with different altitudes is needed. Susila et al.24 reported that spore production was influenced by environmental factors. There was a negative correlation between the number of spores and altitude. The number of spores in the rhizosphere of shallot increased as the altitude decreased from the sea level in West Sumatra.
CONCLUSION
There was a difference in the level of compatibility between the inoculated AMF types regarding vegetative and generative growth of rice cultivated by the SRI method. The AMF Glomus sp3 and Sclerocystis sp. produced the most effective response regarding vegetative and generative growth. In addition, both species provided the highest level of colonization and produced the most spores, making them potential sources of inoculum that can be used in the SRI method of rice cultivation.
SIGNIFICANCE STATEMENT
This study discovers the indigenous mycorrhizae in the rhizosphere of rice cultivated by the System of Rice Intensification method. The study will help the researcher to uncover the critical areas of native mycorrhizae that are also found in wetlands and refute the theory that mycorrhizae only exist on dry land that many researchers were not able to explore. Thus the new theory of indigenous mycorrhiza in wetland rice fields may be arrived at.
ACKNOWLEDGMENT
This study was supported and funded by DRPM RISTEK DIKTI and P3M of Payakumbuh State Agricultural Polytechnic, West Sumatra, Indonesia.
REFERENCES
- Uphoff, N., 2003. Higher yields with fewer external inputs? The system of rice intensification and potential contributions to agricultural sustainability. Int. J. Agric. Sustain., 1: 38-50.
CrossRefDirect Link - Vimal, S.R., J.S. Singh, N.K. Arora and S. Singh, 2017. Soil-plant-microbe interactions in stressed agriculture management: A review. Pedosphere, 27: 177-192.
CrossRefDirect Link - Ruzicka, D., S. Chamala, F.H. Barrios-Masias, F. Martin and S. Smith et al., 2013. Inside arbuscular mycorrhizal roots-molecular probes to understand the symbiosis. Plant Genome, Vol. 6.
CrossRefDirect Link - Nogueira, M.A. and E.J.B.N. Cardoso, 2006. Plant growth and phosphorus uptake in mycorrhizal rangpur lime seedlings under different levels of phosphorus. Pesquisa Agropecuaria Brasil., 41: 93-99.
CrossRefDirect Link - Tawaraya, K., M. Naito and T. Wagatsuma, 2006. Solubilization of insoluble inorganic phosphate by hyphal exudates of arbuscular mycorrhizal fungi. J. Plant Nutr., 29: 657-665.
CrossRefDirect Link - Gathorne-Hardy, A., D.N. Reddy, M. Venkatanarayana and B. Harriss-White, 2016. System of rice intensification provides environmental and economic gains but at the expense of social sustainability: A multidisciplinary analysis in India. Agric. Syst., 143: 159-168.
CrossRefDirect Link - Bingham, M.A. and S.W. Simard, 2013. Seedling genetics and life history outweigh mycorrhizal network potential to improve conifer regeneration under drought. For. Ecol. Manage., 287: 132-139.
CrossRefDirect Link - Higo, M., K. Isobe, M. Yamaguchi, R.A. Drijber, E.S. Jeske and R. Ishii, 2013. Diversity and vertical distribution of indigenous arbuscular mycorrhizal fungi under two soybean rotational systems. Biol. Fertil. Soils, 49: 1085-1096.
CrossRefDirect Link - Verbruggen, E. and E.T. Kiers, 2010. Evolutionary ecology of mycorrhizal functional diversity in agricultural systems. Evol. Applic., 3: 547-560.
CrossRefDirect Link - Bonfante, P. and A. Genre, 2010. Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis. Nature Commun., Vol. 1.
CrossRefDirect Link - Cruz, C., J.J. Green, C.A. Watson, F. Wilson and M.A. Martins-Loucao, 2004. Functional aspects of root architecture and mycorrhizal inoculation with respect to nutrient uptake capacity. Mycorrhiza, 14: 177-184.
CrossRefDirect Link - Cavagnaro, T.R., S.F. Bender, H.R. Asghari and M.G. van der Heijden, 2015. The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci., 20: 283-290.
CrossRefDirect Link - Janos, D.P., M.S. Schroeder, B. Schaffer and J.H. Crane, 2001. Inoculation with arbuscular mycorrhizal fungi enhances growth of Litchi chinensis Sonn. trees after propagation by air-layering. Plant Soil, 233: 85-94.
CrossRefDirect Link - Lehman, R.M., W.I. Taheri, S.L. Osborne, J.S. Buyer and D.D. Douds, Jr., 2012. Fall cover cropping can increase arbuscular mycorrhizae in soils supporting intensive agricultural production. Applied Soil Ecol., 61: 300-304.
CrossRefDirect Link - Hodge, A. and A.H. Fitter, 2010. Substantial nitrogen acquisition by arbuscular mycorrhizal fungi from organic material has implications for N cycling. Proc. Nat. Academy Sci., 107: 13754-13759.
CrossRefDirect Link - Rillig, M.C., C.A. Aguilar‐Trigueros, J. Bergmann, E. Verbruggen, S.D. Veresoglou and A. Lehmann, 2015. Plant root and mycorrhizal fungal traits for understanding soil aggregation. New Phyt., 205: 1385-1388.
CrossRefDirect Link - Susila, E., A. Anwar, A. Syarif and A. Agustian, 2017. Population and diversity of indigenous arbuscular mycorrhizal fungi from shallots rhizosphere in different altitudes in West Sumatra. Int. J. Adv. Sci. Eng. Infor. Technol., 7: 1886-1893.
CrossRefDirect Link
Nelson Elita Reply
Dear PJN
I am very satisfied with the publication of this article. Thank you for your cooperation.