
Widodo
Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Ari Surya Sukarno
Graduate Studies on Biotechnology, Universitas Gadjah Mada, Yogyakarta, Indonesia
Kafaah Estancia
Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Donny Widianto
Faculty of Agriculture, Universitas Gadjah Mada, Yogyakarta, Indonesia
Indratiningsih
Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 11 | Page No.: 524-529







ABSTRACT
Background and Objective: Conjugated linoleic acid (CLA) is a bioactive compound that can be synthesized by probiotics. The goals of this study were to detect CLA production in milk fermented with Lactobacillus casei strain AG and to measure the transcription levels of CLA synthesis-associated genes. Materials and Methods: CLA in fermented milk was detected by gas chromatography-mass spectrometry. The predicted homologous of the cla-hy, cla-dh and cla-dc genes were identified by polymerase chain reaction amplification. For CLA analysis, bacterial cultures were grown in media with or without the addition of 0.4 mg mL–1 linoleic acid. Results: Amplification products of the partial cla-hy, cla-dh and cla-dc homologous of the L. casei strain AG were obtained. The addition of linoleic acid did not change the transcription level of these genes compared to the control (p>0.05). Conclusion: Lactobacillus casei strain AG produced CLA in fermented milk but the genes involved in CLA synthesis were not induced by linoleic acid.
PDF Abstract XML References Citation
Received: May 31, 2018;
Accepted: July 18, 2018;
Published: October 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Widodo, Ari Surya Sukarno, Kafaah Estancia, Donny Widianto and Indratiningsih, 2018. Conjugated Linoleic Acid Synthesis in Milk Fermented with Lactobacillus casei Strain AG. Pakistan Journal of Nutrition, 17: 524-529.
DOI: 10.3923/pjn.2018.524.529
URL: https://scialert.net/abstract/?doi=pjn.2018.524.529
DOI: 10.3923/pjn.2018.524.529
URL: https://scialert.net/abstract/?doi=pjn.2018.524.529
INTRODUCTION
Probiotics are living microorganisms that benefit human health when ingested in adequate doses. Probiotics produce several metabolic compounds with bioactive properties, including conjugated linoleic acid (CLA). Lactobacillus casei has been shown to produce CLA isomers cis-9, trans-11-octadecadienoic acid, trans-9, cis-12-octadecadienoic acid and trans-9, trans-11-octadecadienoic acid1. The best-studied CLA isomers in relation to their in vitro and in some cases, in vivo health effects are cis-9, trans-11- and trans-10, cis-12-octadecadienoic acid2,3. CLA refers to a mixture of positional geometric isomers of linoleic acid (LA) that have two conjugated double bonds. Recently, isomers cis-9, trans-11- and trans-10 cis-12-octadecadienoic acid have been shown to provide potential anti-carcinogenic, anti-inflammatory, anti-obesogenic and anti-cholesterolemic effects2,4,5. The minimum CLA doses at which physiological effects were observed were 3.0 g CLA/day in animals and 0.1-10 g/day in humans6,7. The cis-9, trans-11 isomer is the most dominant CLA isomer in milk fat and accounts for 75-90% of total CLA, while trans-10, cis-12-octadecadienoic acid accounts for 5% of the total CLA in milk8-10.
Fermentation using specific lactic acid bacteria (LAB) starter cultures in milk and meat can increase the content of CLA. The content of CLA in fermented milk products ranges from approximately 4.2-30 mg g–1 of milk fat11-13. The fermentation of low-fat milk (0.05% fat) in yoghurt increased the CLA content from 4.40-5.25 mg total CLA g–1 of total fat. During storage, the content of CLA in the yoghurt did not change, which indicates that CLA is stable during storage14.
Kishino et al.15 isolated and selected CLA-producing LAB and identified that the Lactobacillus plantarum strain AKU 1009a produced the highest CLA levels. The production of CLA by LAB is known to occur through the isomerisation of LA catalysed by linoleate isomerase. Linoleate isomerase from L. plantarum AKU 1009a is part of a multi-component enzyme system. The genes that express the enzymes of this system have been identified and include cla-hy (Accession number AB671229.1), cla-dh (Accession number AB671230.1) and cla-dc (Accession number AB671231.1)16. Lactobacillus plantarum AKU 1009a was able to convert LA into CLA with yields as high as 40 mg mL–1 , of which 38% was cis-9, trans-11-octadecadienoic acid and 62% was trans-9, trans-11-octadecadienoic acid17.
In our previous study, we isolated L. casei strain AG from the human gastrointestinal tract, for its potential use as a probiotic18. Based on an in silico analysis, the L. casei strain AG has one or more homologous of the L. plantarum AKU 1009a genes that are responsible for CLA synthesis. Therefore, this study aims to investigate the ability of the L. casei strain AG to synthesise CLA during milk fermentation and to measure the transcription level of CLA-synthesis genes upon the addition of LA.
MATERIALS AND METHODS
Starter culture and growth conditions: The L. casei strain AG, isolated from a human intestine, was used in this study18. Bacterial cells were sub-cultured by plating on de Man-Rogosa-Sharpe agar (MRS, Merck, Jakarta, Indonesia) supplemented with 0.5 g L–1 L-cysteine (Sigma-Aldrich Ltd., Singapore) and incubated at 37°C for 24 h under micro-aerobic conditions.
Milk fermentation and sample preparation: Fresh milk was pasteurised at 80°C for 30 min. The pasteurised milk was inoculated with a bulk starter L. casei strain AG (5% v/v) and incubated at 37°C overnight (18 h). Samples were then harvested by centrifugation at 5000 rpm/5°C for 30 min. The supernatants were collected for CLA analysis by gas chromatography-mass spectrometry (GC-MS), according to Alonso et al.1
Fat extraction: Fat extraction was carried out according to Alonso et al.1. Fermentation supernatants (6 mL) were mixed with 12 mL isopropanol and homogenised. The solution was then mixed with 9 mL hexane and centrifuged at 23,000 rpm/5°C for 5 min. The upper layer was collected, filtered using sodium sulphate and washed with 7 mL hexane. Evaporation of the hexane at room temperature (30-40°C) resulted in condensed fat. The esterification of the fat residue was carried out with 300 μL of 14% boron trifluoride in methanol at 50-60°C for 2 h. After esterification, 800 μL hexane was added and the sample was analyzed by GC-MS.
GC-MS analysis: GC-MS analysis was performed using a DB-1 Agilent J and W column (30 m×0.25 mm i.d.) with helium as the carrier gas and electron ionisation at 70 eV. The following conditions were used: a column heater temperature of 80°C, injection temperature of 310°C, injection model split, 16.5 kPa, 40 mL min–1 total flow and 0.50 mL min–1 column flow, with a split ratio of 73.2 mL injection volume. The CLA peak was identified by retention time comparisons.
Polymerase chain reaction (PCR) amplification: Genomic DNA was isolated using the Geneaid gSYNC™ DNA Extraction Kit (Geneaid Biotech Ltd., New Taipei City, Taiwan).
| Table 1: | Primers used to amplify the partial homologue genes in L. casei strain AG |
 | |
PCR amplification (Applied Biosystems thermal cycler 2720, Foster City, CA, USA) was performed using 2x KAPA Taq Readymix (Kapa Biosystems, Cape Town, South Africa), according to the manufacturer’s instructions. The primers used to detect CLA-synthesis genes were designed to recognise conserved regions of cla-hy, cla-dh and cla-dc genes, based on published L. casei genomes. The primer sequences were designed using primer 3 and are presented in Table 1. The following amplification programme was used: 1 min at 95°C, followed by 30 cycles of 30 sec at 94°C, 30 sec at 55°C, 2 min at 72°C and finally, 1 cycle of 5 min at 72°C. Amplified bands were resolved by electrophoresis on 2% (w/v) agarose gel (Sigma-Aldrich Ltd) and visualised by ethidium bromide staining.
The purified DNA was sequenced using the Applied Biosystem 3730-XL Analyser (1st Base, Singapore). The resulting sequences were compared using the National Centre for Biotechnology Information (NCBI) database. Phylogenetic trees were constructed in Mega 6.0.
Quantitative real-time PCR (qRT-PCR) analysis: Bacterial cultures were grown in a medium with or without the addition of 0.4 mg mL–1 LA (Sigma-Aldrich Ltd.). LA was added as an aqueous solution at 1% (v/v) in Tween 80 (Merck). The broth medium was inoculated with 2% (v/v) of a bacterial starter culture and incubated at 37°C for 4 h. After incubation, cell cultures were harvested for total RNA isolation.
For qRT-PCR analysis, total RNA was extracted using the Geneaid PrestoTM Mini RNA Bacteria Kit (Geneaid Biotech Ltd., New Taipei City, Taiwan) with a DNAse treatment, according to the manufacturer’s instructions. RNA concentration was measured using spectrophotometry. RT-PCR (Real-Time PCR Bio-Rad CFX-96 thermal cycler, Singapore) was carried out using a KAPA SYBR® FAST Universal One-Step qRT-PCR Kit (Kapa Biosystems) in duplicate. Relative quantification of the results was calculated using the equation 2-ΔΔCT 19. Differences between the treatments and control were analyzed by independent t-tests using SPSS 16.0 (SPSS Inc., Chicago).
RESULTS AND DISCUSSION
CLA detection in fermented milk: CLA in milk fermented by the L. casei strain AG and corresponding non-fermented milk standard was detected in the form of fatty acid methyl esters (FAME). FAME forms of CLA and LA can be distinguished by GC-MS, due to their different retention times and same molecular weights. Fresh milk (non-fermented milk standard) used in this study contains LA but does not include CLA (data not shown). GC-MS results showed that milk fermented by the L. casei strain AG contained CLA (Fig. 1).
This study showed that the L. casei strain AG produced CLA isomers when it was used as a starter culture for milk fermentation (Fig. 1). Based on retention time, the chromatogram peak number 23 in milk fermented with the L. casei strain AG (Fig. 1) was close to the peak representing the CLA standard trans-10, cis-12-octadecadienoic acid. Meanwhile, the chromatogram peak number 19 was LA with a molecular weight (m/z) of 294. The presence of CLA in milk fermented by the L. casei strain AG was in agreement with the previous study by Alonso et al.1. Using L. casei and L. acidophilus as starters to ferment skimmed milk, Alonso et al.1 observed CLA production in milk supplemented with 0.02% LA. CLA isomers produced by L. casei were cis-9, trans-11-, trans-10, cis-12- and trans-9, trans-11-octadecadienoic acid. However, CLA isomers produced by the L. casei strain AG have not been previously identified.
Detection CLA-synthesising genes: The nucleotide sequences of cla-hy, cla-dh and cla-dc genes from L. plantarum AKU 1009a were aligned with the genome of L. casei strain Zhang and this approach revealed homologous of 1710, 861 and 876 bp, respectively. These homologue sequences were used to design primers cla-hy, cla-dh and cla-dc, with amplification products of 143, 206 and 161 bp, respectively. The length of the amplification products generated by these three primers was confirmed to be about 100-200 bp (Fig. 2).
The size of the amplification products obtained from L. casei strain AG, using the three selected primer pairs, correlated with the predicted target lengths.
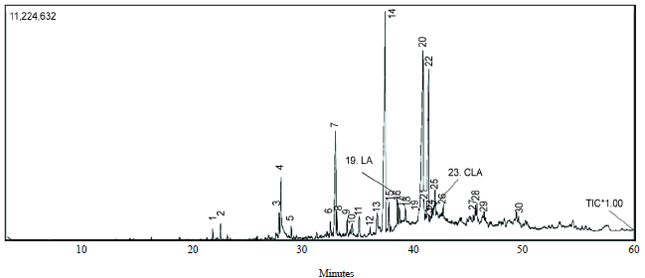 | |
| Fig. 1: | Gas chromatography-mass spectrometry chromatogram, indicating the peaks corresponding to linolenic acid (LA) and conjugated linolenic acid (CLA) |
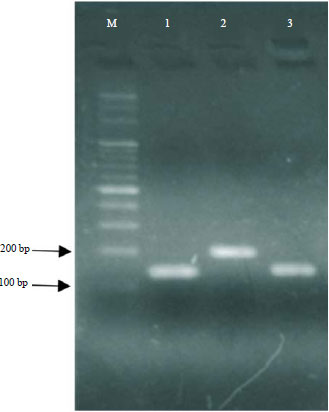 | |
| Fig. 2: | Amplified bands obtained using the primers cla-hy (1), cla-dh (2) and cla-dc (3) |
The amplified bands were then sequenced, to obtain nucleotide sequences. The nucleotide sequence of the cla-hy amplification product showed a 76% homology with the genes putatively encoding linoleate isomerase from Lactobacillus sakei strain LMG 13558 and 73% homology with the cla-hy gene from L. plantarum (Fig. 3). The nucleotide sequence of the cla-dh amplification product had 100% homology to the putative short-chain dehydrogenase/oxidoreductase gene from L. paracasei strain 1195 and 74% homology with the cla-dh gene from L. plantarum (Fig. 4). The nucleotide sequence of the cla-dc amplification product was 148 bp, instead of the expected 161 bp. This sequence was 77% homologous to the cla-dc gene for isomerase from L. plantarum (Fig. 5).
Some LAB, especially those belonging to the genus Lactobacillus, produce CLA from LA and ricinoleic acid. CLA production is a response to LA toxicity, as converting LA into CLA can reduce its toxicity20. Previously, it was thought that CLA was generated as the product of LA catalysis by a single enzyme, linoleate isomerase. However, Kishino et al.16 showed that catabolism of LA into CLA involves multiple enzymes and reactions. The pathway for CLA synthesis has been described for L. plantarum AKU 1009a, which produces CLA isomers trans-10, cis-12-octadecadienoic acid and cis-9, trans-11-octadecadienoic acid. Synthesis of the CLA isomer trans-10, cis-12-octadecadienoic acid requires only a single enzyme that is expressed by the cla-hy gene. The other isomer, cis-9, trans-11-octadecadienoic acid is synthesized by two enzymes that are expressed by the cla-dh and cla-dc genes. The majority of species within the Lactobacillus genus possess one or more of these genes. The synthesis of CLA isomers depends on the enzyme cofactors FAD and NADH or NADPH.
Transcription of CLA-synthesising genes: The addition of LA in the growth medium was used to induce CLA synthesis.
 | |
| Fig. 3: | Phylogenetic tree of cla-hy based on the sequence amplified from Lactobacillus casei AG |
 | |
| Fig. 4: | Phylogenetic tree of cla-dh based on the sequence amplified from Lactobacillus casei AG |
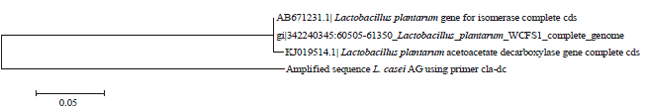 | |
| Fig. 5: | Phylogenetic tree of cla-dc based on the sequence amplified from Lactobacillus casei AG |
In this study, we measured the transcription levels of the cla-hy, cla-dh and cla-dc genes. The gene expression of cla-hy, cla-dh and cla-dc, when grown in m e dia with the addition of 0.4 mg mL–1 LA, was normalised to an expression without the addition of the control.
The expression level of the cla-hy, cla-dh and cla-dc genes upon the addition of 0.4 mg mL–1 LA was 0.74, 0.69 and 1.43, respectively. The addition of 0.4 mg mL–1 LA in the growth medium did not change the expression of these genes (p>0.05). According to Alonso et al.1, the addition of 0.4 mg mL–1 LA in the growth medium of L. casei E4 and E10 increased CLA production. In the current study, the gene expression of cla-hy, cla-dh and cla-dc was not upregulated by the addition of 0.4 mg mL–1 LA.
Milk fermented by the L. casei strain AG produced CLA and it possesses CLA-synthesising genes. However, the transcription activity of the cla-hy, cla-dh and cla-dc genes in the L. casei strain AG was not induced by the addition of 0.4 mg mL–1 LA.
SIGNIFICANCE STATEMENT
This study discovers the synthesis of conjugated linoleic acid (CLA) in milk fermented with L. casei strain AG. The ability of L. casei AG to synthesise CLA in fermented milk can be beneficial for human health. This study will support the utilization of L. casei AG as starter culture for dairy fermentation and thus develop probiotic-based food industries.
ACKNOWLEDGMENT
This study was fully supported by Hibah Kompetensi, provided by the Indonesian Ministry for Research Technology and Higher Education.
REFERENCES
- Alonso, L., E.P. Cuesta and S.E. Gilliland, 2003. Production of free conjugated linoleic acid by Lactobacillus acidophilus and Lactobacillus casei of human intestinal origin. J. Dairy Sci., 86: 1941-1946.
CrossRefDirect Link - Bhattacharya, A., J. Banu, M. Rahman, J. Causey and G. Fernandes, 2006. Biological effects of conjugated linoleic acids in health and disease. J. Nutr. Biochem., 17: 789-810.
CrossRefPubMedDirect Link - Benjamin, S. and F. Spener, 2009. Conjugated linoleic acids as functional food: An insight into their health benefits. Nutr. Metab., Vol. 6.
CrossRefDirect Link - O'Shea, E.F., P.D. Cotter, C. Stanton, R.P. Ross and C. Hill, 2012. Production of bioactive substances by intestinal bacteria as a basis for explaining probiotic mechanisms: Bacteriocins and conjugated linoleic acid. Int. J. Food Microbiol., 152: 189-205.
CrossRefDirect Link - Ip, C., M. Singh, H.J. Thompson and J.A. Scimeca, 1994. Conjugated linoleic acid suppresses mammary carcinogenesis and proliferative activity of the mammary gland in the rat. Cancer Res., 54: 1212-1215.
PubMedDirect Link - Jiang, J., L. Bjorck and R. Fonden, 1998. Production of conjugated linoleic acid by dairy starter cultures. J. Applied Microbiol., 85: 95-102.
CrossRefDirect Link - Chin, S.F., W. Liu, J.M. Storkson, Y.L. Ha and M.W. Pariza, 1992. Dietary sources of conjugated dienoic isomers of linoleic acid, a newly recognized class of anticarcinogens. J. Food Compos. Anal., 5: 185-197.
CrossRefDirect Link - Kennedy, A., K. Martinez, S. Schmidt, S. Mandrup, K. LaPoint and M. McIntosh, 2010. Antiobesity mechanisms of action of conjugated linoleic acid. J. Nutr. Biochem., 21: 171-179.
CrossRefDirect Link - Khanal, R.C. and K.C. Olson, 2004. Factors affecting Conjugated Linoleic Acid (CLA) content in milk, meat and egg: A review. Pak. J. Nutr., 3: 82-98.
CrossRefDirect Link - Parodi, P.W., 1994. Conjugated linoleic acid: An anticarcinogenic fatty acid present in milk fat. Aust. J. Dairy Technol., 49: 93-97.
Direct Link - Lin, H., T.D. Bovlston, M.J. Chang, L.O. Luedecke and T.D. Shultz, 1995. Survey of the conjugated linoleic acid contents of dairy products. J. Dairy Sci., 78: 2358-2365.
PubMedDirect Link - Dhiman, T.R., G.R. Anand, L.D. Satter and M.W. Pariza, 1999. Conjugated linoleic acid content of milk from cows fed different diets. J. Dairy Sci., 82: 2146-2156.
CrossRefDirect Link - Shantha, N.C., L.N. Ram, J. O'Leary, C.L. Hicks and E.A. Decker, 1995. Conjugated linoleic acid concentrations in dairy products as affected by processing and storage. J. Food Sci., 60: 695-697.
CrossRefDirect Link - Kishino, S., J. Ogawa, Y. Omura, K. Matsumura and S. Shimizu, 2002. Conjugated linoleic acid production from linoleic acid by lactic acid bacteria. J. Am. Oil Chem. Soc., 79: 159-163.
CrossRefDirect Link - Kishino, S., M. Takeuchi, S.B. Park, A. Hirata and N. Kitamura et al., 2013. Polyunsaturated fatty acid saturation by gut lactic acid bacteria affecting host lipid composition. Proc. Natl. Acad. Sci. USA., 110: 17808-17813.
CrossRefDirect Link - Kishino, S., J. Ogawa, A. Ando, T. Iwashita, T. Fujita, H. Kawashima and S. Shimizu, 2003. Structural analysis of conjugated linoleic acid produced by Lactobacillus plantarum and factors affecting isomer production. Biosci. Biotechnol. Biochem., 67: 179-182.
CrossRefDirect Link - Widodo, T.T. Taufiq, E. Aryati, A. Kurniawati and W. Asmara, 2012. Human origin Lactobacillus casei isolated from Indonesian infants demonstrating potential characteristics as probiotics in vitro. Indonesian J. Biotechnol., 17: 79-89.
Direct Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Jenkins, J.K. and P.D. Courtney, 2003. Lactobacillus growth and membrane composition in the presence of linoleic or conjugated linoleic acid. Can. J. Microbiol., 49: 51-57.
CrossRefDirect Link